Associative learning by C. elegans is variable when butanone is paired with starvation
Samiha Tasnim, Amber Liu, Antony Jose

TL;DR
The study shows that C. elegans worms show inconsistent learning when butanone is paired with starvation, suggesting that such learning might be rare in natural environments.
Contribution
The study reveals unexplained variability in associative learning in C. elegans when exposed to butanone and starvation.
Findings
Pre-exposure to butanone with starvation led to inconsistent chemotaxis responses in C. elegans.
Variability in associative learning was observed despite controlled laboratory conditions.
The findings suggest that associative learning in natural environments may be rare.
Abstract
The nematode C. elegans has been reported to show a reduction in its preference for the odorant butanone after prior exposure to butanone coupled with starvation. Here we report unexplained variability in such associative learning. Pre-exposure of unfed worms to butanone resulted in different responses during different trials of subsequent chemotaxis assays – from strong avoidance to enhanced attraction. Given this variation in associative learning despite the artificially controlled lab setting, we speculate that in dynamic natural environments such learning might be rare and highlight the challenge in discovering evolutionarily selected mechanisms that could underlie learning in the wild.
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
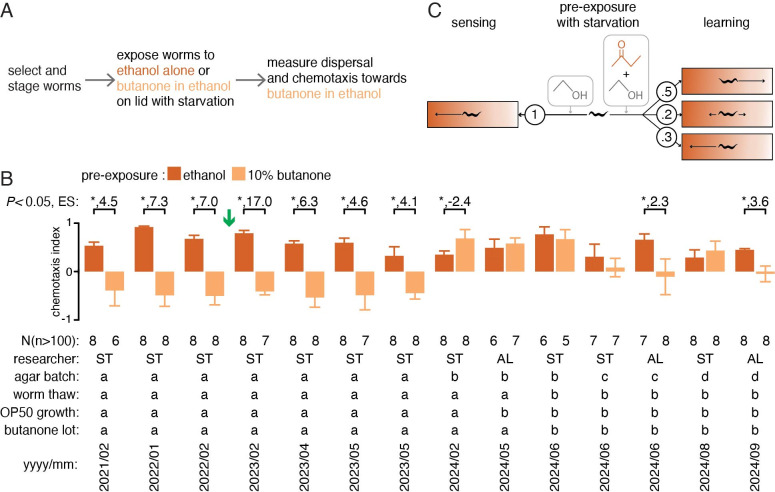 Figure 1
Figure 1|
|
|
|
Agar |
VWR cat#97064-334 unknown lot (batch a in Fig. 1
VWR cat#97064-334 lot#23K0156549 (batch b in Fig. 1
Carolina® cat#842130 (batch c in Fig. 1
VWR cat#97064-334 lot#24G2356953 (batch d in Fig. 1
|
|
Bacto-peptone |
Gibco™, Thermo Fisher Scientific cat#211677 unknown lot for Fig. 1
Gibco™, Thermo Fisher Scientific cat#211677 lot#4024890 for Fig. 1
|
|
NaCl |
Supelco®, VWR cat# EMD-SX0420-1 |
|
Cholesterol |
Sigma-Aldrich cat# C8667-5G |
|
CaCl |
MilliporeSigma™, FisherScientific cat#M1023820500 |
|
MgSO 4 |
MilliporeSigma™, FisherScientific cat#MMX00751 |
|
K2HPO 4 |
VWR Chemicals BDH® cat#BDH9266-500G |
|
KH 2 PO 4 |
VWR Chemicals BDH® cat#BDH9268-2.5KG |
|
Na 2 HPO 4 *7H 2 O |
Unknown for Fig. 1
Spectrum Chemical, The Lab Depot cat#S1400-500GM-EA for Fig. 1
|
|
KH 2 PO 4 |
VWR Chemicals BDH® cat#BDH9268-2.5KG |
|
MgSO4 |
MilliporeSigma™, FisherScientific cat#MMX00751 |
|
NaOH |
Supelco®, MilliporeSigma cat#SX0590-3 |
|
Ethanol |
Sigma Aldrich, cat#E7023-500ML |
|
2-
|
Sigma Aldrich, cat#360473-500ML. Lot#SHBL7664 for Fig. 1
|
|
35 mm plate |
Falcon™ Bacteriological Petri Dishes with Lid, FisherScientific, cat#08-757-100A |
|
100 mm plate |
Falcon® Petri Dishes, VWR, cat#25373-100 |
|
Assay plate |
Nunc® Rectangular Dishes, VWR cat#73521-424 |
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsEmotional Intelligence and Performance
Description
Despite their short lives and complex environments, * C. elegans * have been reported to be capable of associative learning (reviewed in Zhang et al., 2024). A ~1 hr pre-exposure to ‘attractive' odorants in the absence of food eliminates the attraction when tested using a subsequent assay and such learning, that presumably associates starvation with the odorant, was not observed if the pre-exposure was done in the presence of food (Nuttley et al., 2002). Incorporating the pre-exposure to butanone with starvation into a recently developed assay for chemotaxis (Fig. 1 A ; Tasnim et al., 2025) resulted in worms moving away from butanone – an altered response that was reproduced in 6 subsequent trials (Fig. 1 B ; effect sizes ranging from ~4.1 to ~17.0). This apparent associative learning was eliminated when the pre-exposure was performed in the presence of food (Supplemental Dataset 1; chemotaxis index (CI) after ethanol pre-exposure = 0.75 +/- 0.12 and CI after butanone pre-exposure = 0.8 +/- 0.18). However, in 7 subsequent trials without food the measured associative learning, if any, varied widely (effect sizes from -2.4, indicating increased attraction to butanone , to 3.6, indicating decreased attraction to butanone ). The previously detected learned butanone avoidance could not be reproduced despite varying agar (2 trials each using 2 new sources), researcher (3 trials by one and 4 by another), worms (5 trials using a 2 ^nd^ isolate), *E. coli * (6 trials using a 2 ^nd^ isolate), and butanone (5 trials using a 2 ^nd^ lot). In contrast, the initial response to odorant sensation remained reproducible ( Fig. 1B ) and initial responses to a different attractive odorant benzaldehyde and an aversive odorant nonanone were also reproducible (Tasnim et al., 2025). Artificially categorizing the responses measured in each trial (Fig. 1 B ) into trials detecting learned avoidance and trials with no detectable learning enables an evaluation of the pattern using the runs test (Swed and Eisenhart, 1943; Bujang and Sapri, 2018). This test does not support a significant deviation from random (r = 5 > L _c _ = 3; see Statistics in Materials and Methods).
Our measurements of behavior without immobilization reveal that unlike the reproducible response upon sensing an odorant (Tasnim et al., 2025), the observed response after associative learning is variable and not distinguishable from random (Fig. 1 C ), suggesting that reproducible learning, if any, requires artificial environments that are not easily controlled. Under this hypothesis, past reports of learning (reviewed in Zhang et al., 2024) likely include undeclared or cryptic experimental conditions (e.g., unknown aspects of agar (Bargmann, 2024)) that are needed for the reproducible detection of associative learning. The discovery of such conditions will support the wider study of the learning detected in the laboratory. Another possibility is that robust learning by * C. elegans * has been lost through growth in the laboratory (Frézal & Félix, 2015). Finally, in the dynamic natural environment of * C. elegans * where evolutionary forces sculpt molecular mechanisms, learning to associate an odorant with starvation could be rare.
Methods
Worm growth and selection
The * C. elegans * wild type Bristol N2 was obtained from the Caenorhabditis Genetics Centre (University of Minnesota, Minneapolis, MN, USA). Worms were grown at 20ºC, and chemotaxis assays were performed at room temperature (~25ºC).
Growth, pre-exposure, and assay plates were prepared using appropriate volumes of Nematode Growth Medium (NGM): 4 ml per 35 mm plate, 20 ml per 100 mm plate and 8 ml per assay plate. NGM was prepared by combining 17 g bacteriological agar, 2.5 g bacto-peptone, and 3 g NaCl in 1 L water, autoclaving, cooling to 60ºC, then adding 1 ml each of cholesterol (5mg/ml in ethanol), 1 M CaCl 2 and 1 M MgSO 4 , and 25 ml of potassium phosphate buffer (pH 6.0) (see Table 1). The use of various sources of agar was inspired by anecdotal reports that behavior can vary between batches of agar (e.g., (Bargmann, 2024)).
Growth plates were seeded with overnight cultures of E. coli OP50 grown at 37ºC in Luria-Bertani (LB) broth (100 µl per 35 mm NGM plate and 500 µl per 100 mm NGM plate) and left at room temperature for ~48 hrs, by when a lawn forms. These seeded plates were stored for up to 4 weeks at 4ºC until use. Worm washes and transfers were done using M9 buffer prepared by adding 5 g NaCl, 11.32 g dibasic Na 2 HPO 4 *7H 2 O and 3 g KH 2 PO 4 to 1 L water, autoclaving, and cooling to ~60ºC before adding 1 ml of 1 M MgSO 4 . A worm bleaching solution (a stock mix of 45 ml 5 M NaOH, 15ml Clorox® bleach (8.25% NaOCl) and 90 ml water aliquots diluted to 50% before use) was used to dissolve worms while leaving the embryos protected by the eggshells. Subsequent hatching resulted in populations of L1-staged animals that were then moved to seeded plates and grown to adulthood before their behavior was assayed. Bleaching was performed only once per experimental replicate and bleached worms were not propagated; unbleached worms were separately maintained as the stock.
Staging
Plates were set up on day 1 by washing an unstarved but crowded 35 mm plate of worms with 1 ml M9 and transferring 10 µl of the suspension onto 12 OP50-seeded 35 mm plates. These plates were grown for 36-48 hrs at 20ºC until large numbers of gravid adults were observed, then worms were bleached by dropwise addition of 250-300 µl of worm bleaching solution, ensuring that all worms were covered using the least volume of bleaching solution to facilitate quick diffusion of bleach into the agar thereby minimizing stress on worms that hatch from the surviving eggs. Bleaching was chosen in contrast to precise staging by picking L4-staged larvae because of the need for hundreds of worms for each experiment and to ensure uniform treatment of all worms (e.g., differences in mechanical handling of worms could impact subsequent behavior). Bleached plates were kept at 20ºC for 24 hrs until the surviving eggs hatched, then L1 worms were pooled from 2 plates by washing with 500 µl M9 and transferred onto a seeded 100 mm plate. These worms were then allowed to grow for an additional 72 hrs (total 96 hrs post-bleach) to get a synchronized cohort of young adult worms which were then pre-exposed and tested for their response to volatile odorant(s). Six 100mm plates of young adult worms were sufficient to pre-expose with one odorant (or vehicle) and to measure both dispersal and chemotaxis in 8 arenas each.
Pre-exposure
M9 buffer (5-6 ml) was used to pool worms from six 100 mm plates into 1.5 ml microcentrifuge tubes and washed a total of 3 times by adding 1 ml of M9 and centrifuging at 11,000 rpm for 2 min. This use of centrifugation is different from the gravity-aided settling that was used in some assays for associative learning (Kaufmann et al., 2011) (we estimate gravity-aided settling until the solution above the worm ‘pellet' is clear to take >7 min, Movie S1). However, centrifugation was consistently used in all trials, including the 9 repeats that showed learning with an effect size >2 (Fig. 1 B ). At the end of the third wash, the supernatant was discarded, retaining 200 µl of liquid with worms and one such tube of worms was used per 100 mm plate during pre-exposure. A P1000 pipette was used to transfer worms onto 100 mm plates with (for one experiment) or without E. coli OP50 food, taking care not to transfer any bacterial pellets and pipetting from the top of the worm “pellet” so as not to transfer any worm carcasses from the bottom of the tube. Ethanol was used as the vehicle and 2- butanone as the test odorant. Both vehicle and odorant stocks were stored in airtight glass bottles protected from the light, while aliquots of each in 1.5 ml Eppendorf tubes were kept for no more than 4 weeks. Fresh odorant dilutions (10% 2- butanone ) were made on the day of the behavioral assay in 1.5 ml microcentrifuge tubes. 5 µl of either the vehicle or the diluted odorant was streaked onto the lid of the plate and the covered plate was then sealed with parafilm. Plates were kept undisturbed at room temperature for 1 hr, then pre-exposed worms were collected and washed three times using M9 buffer. At the end of the third wash the supernatant was discarded, retaining ~200 µl of liquid with worms.
Chemotaxis and dispersal assays
Arenas were set up as described (Fig. 1 A in Tasnim et al., 2025) and for each test (dispersal or chemotaxis), 2 sets of 4 rectangular arenas were used per pre-exposure treatment (N=8). A template was used to trace quadrants onto extra lids of plates and these lids were placed under each set of 4 arenas to aid transfer of worms to the origins to initiate each assay. A P20 pipette with the tip cut off to increase the bore was used to transfer 10 µl of worms from the top of the worm “pellet” onto the center (origin) of each rectangular plate, minimizing transfer of worm carcasses or remnant bacteria from the bottom of the tube. Typically, one tube of pre-exposed worms yielded enough for testing >100 worms per plate on 4 sets of 4 rectangular arenas (N=16). After transferring worms, 2.5 µl of the odorant in vehicle or the vehicle alone was pipetted at either end of the chemotaxis plates in opposite ends such that the worms in one set of arenas must move in the opposite direction to worms in the other set for the same response. With this orientation, if worms were responding to a gradient of an unknown cue outside the arena, then they will move in the same direction in both arenas, thereby reducing effect size in response to the odorant within the arena. The transfer of worms and odorants/vehicle typically took a total of 5 min per 4 set of 4 rectangular arenas (N=16). The plates were left with the lids on at room temperature for 1 hr. At the end of 1 hr, videos were taken of the plates keeping the extra lid underneath to allow visualization of the quadrants, and the videos were used to count the numbers of young adult worms in each quadrant. The video was taken using an iPhone positioned over the objective using an adapter and typically took 5 min per 4 set of 4 rectangular arenas (N=16).
Statistics
P -values, effect sizes, and error bars were calculated as described in Tasnim et al., 2025. The distribution of outcomes after each trial in Fig. 1 B was tested for significant deviations from random using the runs test (Swed and Eisenhart, 1943; Bujang and Sapri, 2018). With numbers of outcome one (n 1 ) = 9, numbers of outcome two (n 2 ) = 5 and numbers of runs (r) = 5, the lower critical value (L c ) obtained was 3 from the table in Swed and Eisenhart, 1943. Therefore, we cannot reject the hypothesis that our series of runs can arise randomly (i.e., r = 5 > L _c _ = 3, therefore the null hypothesis for one-sided test cannot be rejected).
Data, Materials, and Software Availability
All data generated and the code used are available at Extended Data.
Reagents
Table 1. Materials and reagents
**: **
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Bargmann C 2024925 Cori Bargmann.Neuron 112180896-62732999300210.1016/j.neuron.2024.09.00839326389 · doi ↗ · pubmed ↗
- 2Bujang MA Sapri FE 2018830 An Application of the Runs Test to Test for Randomness of Observations Obtained from a Clinical Survey in an Ordered Population.Malays J Med Sci 2541394-195X 14615110.21315/mjms 2018.25.4.1530914857 PMC 6422539 · doi ↗ · pubmed ↗
- 3Frézal L Félix MA 2015330 C. elegans outside the Petri dish.Elife 410.7554/e Life.0584925822066 PMC 4373675 · doi ↗ · pubmed ↗
- 4Kauffman A Parsons L Stein G Wills A Kaletsky R Murphy C 2011311 C. elegans positive butanone learning, short-term, and long-term associative memory assays.J Vis Exp 4910.3791/249021445035 PMC 3197297 · doi ↗ · pubmed ↗
- 5Nuttley WM Atkinson-Leadbeater KP Van Der Kooy D 2002829 Serotonin mediates food-odor associative learning in the nematode Caenorhabditiselegans.Proc Natl Acad Sci U S A 99190027-8424124491245410.1073/pnas.19210169912202746 PMC 129465 · doi ↗ · pubmed ↗
- 6Swed Frieda S. Eisenhart C. 194331 Tables for Testing Randomness of Grouping in a Sequence of Alternatives The Annals of Mathematical Statistics 1410003-4851668710.1214/aoms/1177731494 · doi ↗
- 7Tasnim S Liu A Jose AM 2025311 A simple yet reliable assay for chemotaxis in C. elegans.Micro Publ Biol 202510.17912/micropub.biology.00151440161433 PMC 11953739 · doi ↗ · pubmed ↗
- 8Zhang Y Iino Y Schafer WR 202494 Behavioral plasticity.Genetics 22810016-673110.1093/genetics/iyae 10539158469 · doi ↗ · pubmed ↗