Three new species of Psectrocladius Kieffer (Diptera, Chironomidae) from China

Abstract
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
 Figure 1
Figure 1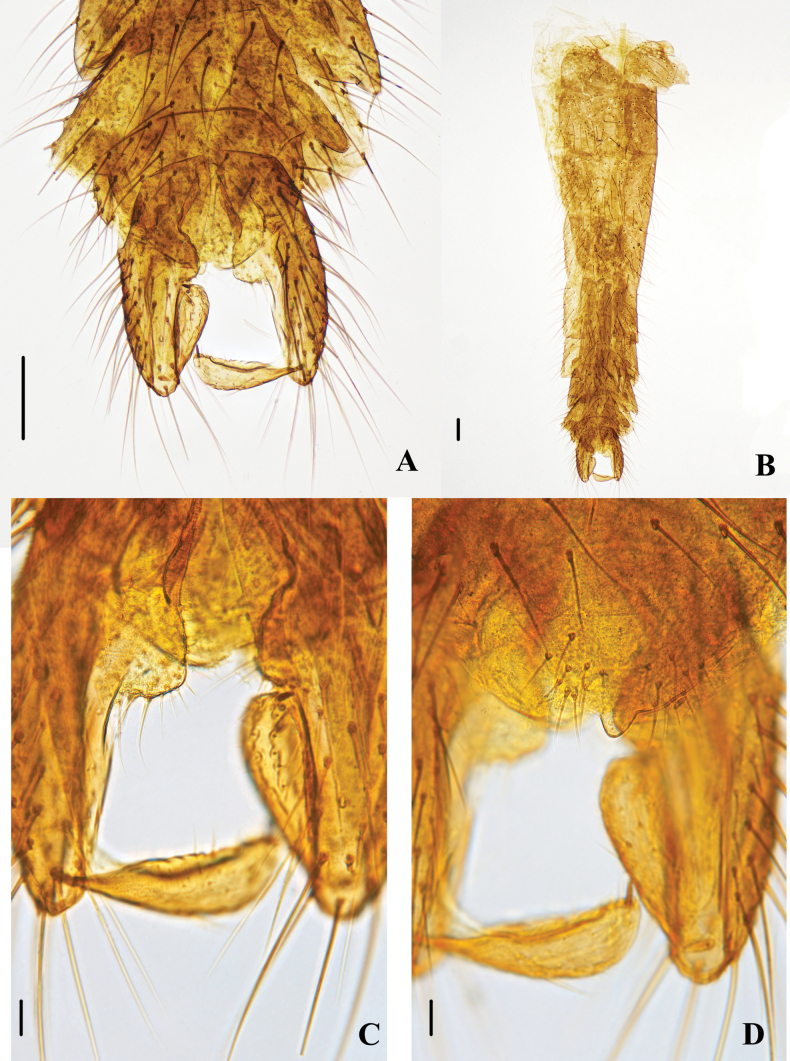 Figure 2
Figure 2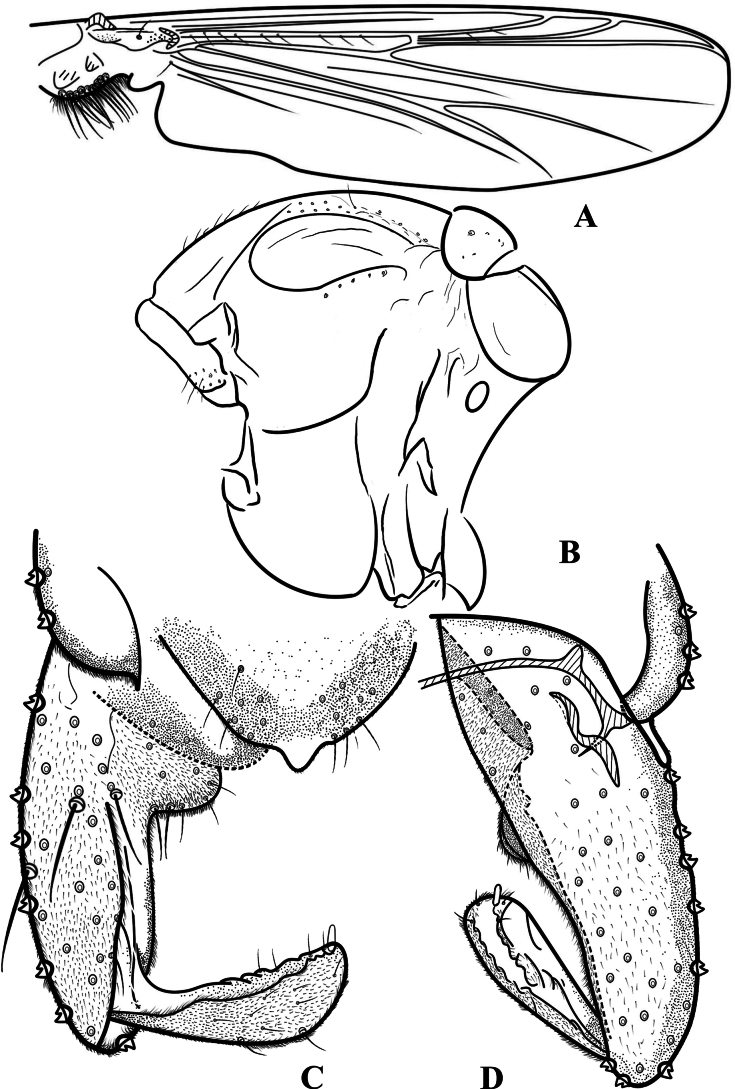 Figure 3
Figure 3 Figure 4
Figure 4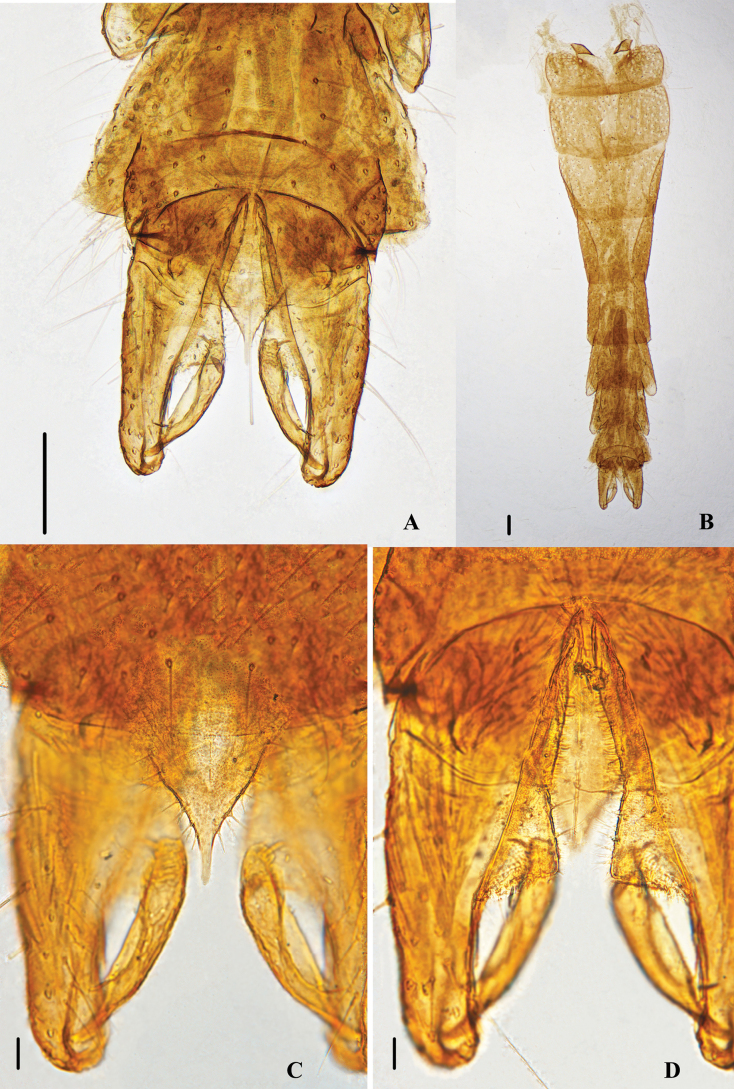 Figure 5
Figure 5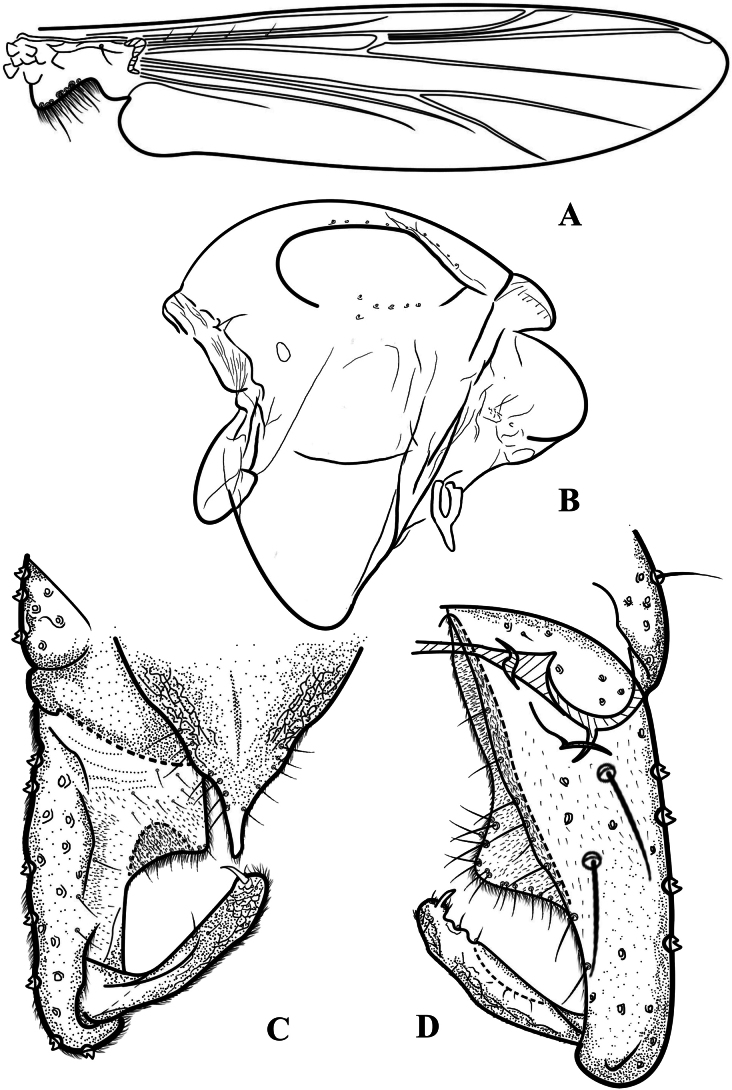 Figure 6
Figure 6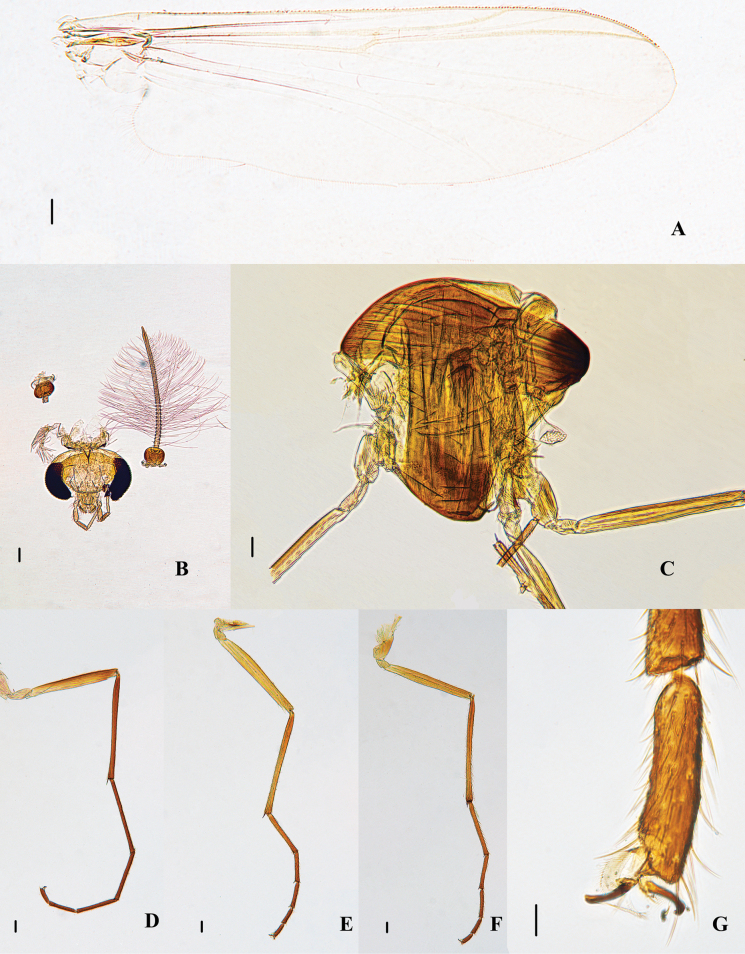 Figure 7
Figure 7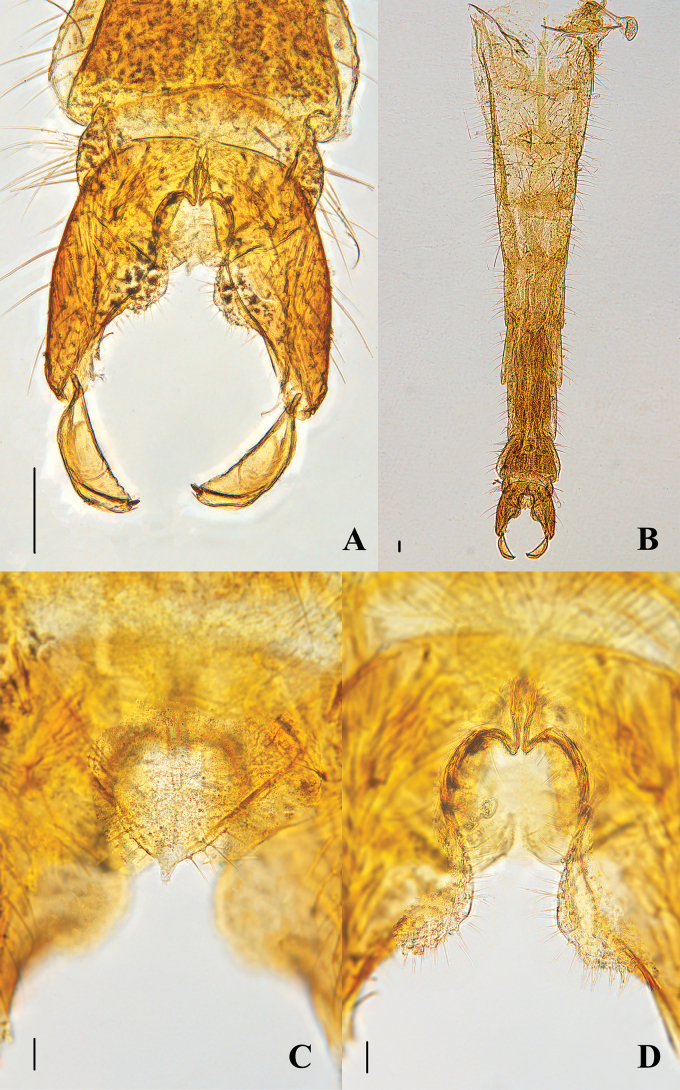 Figure 8
Figure 8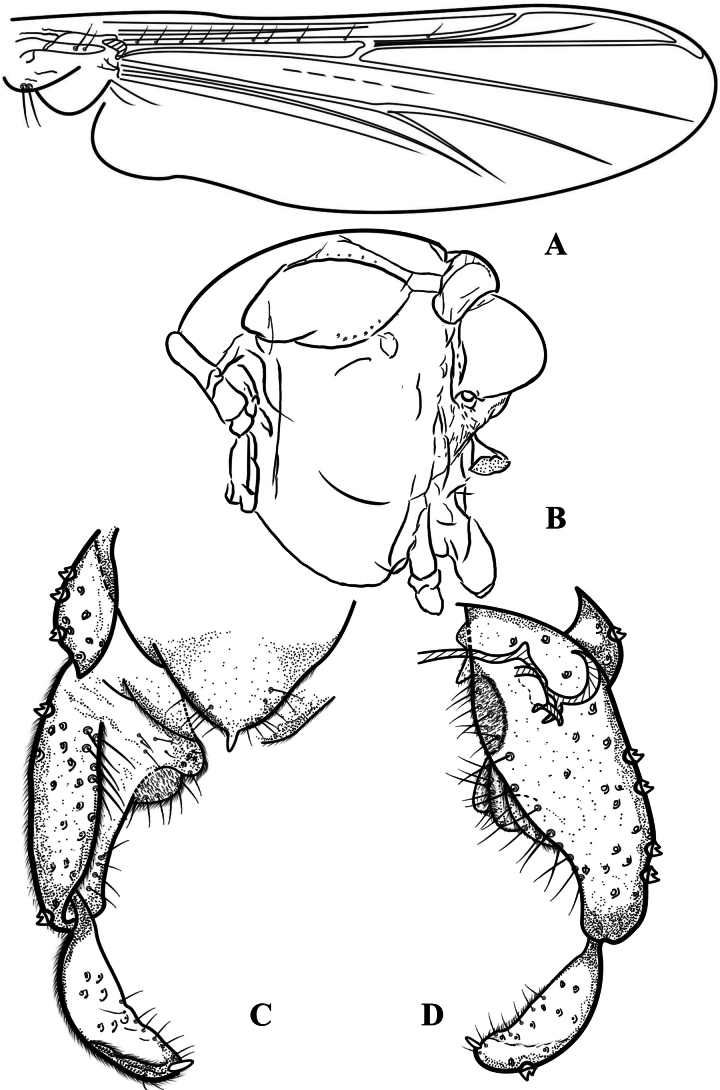 Figure 9
Figure 9|
|
|
|
|
| |
| P1 | 990–1188, 1074 | 822–927, 864 | 567–742, 625 | 339–432, 378 | 269–324, 288 |
| P2 | 823–984, 882 | 762–889, 822 | 371–453, 398 | 242–270, 242 | 180–229, 197 |
| P3 | 819–945, 878 | 681–817, 754 | 568–697, 604 | 373–445, 399 | 262–328, 283 |
|
|
|
|
|
| |
| P1 | 110–130, 118 | 136–171, 152 | 0.55–0.62, 0.58 | 2.70–2.76, 2.74 | 2.85–3.27, 3.12 |
| P2 | 107–119, 113 | 112–130, 123 | 0.43–0.47, 0.45 | 2.94–3.16, 3.07 | 4.13–4.48, 4.29 |
| P3 | 111–123, 115 | 158–192, 176 | 0.65–0.74, 0.69 | 2.26–2.35, 2.30 | 2.53–2.85, 2.71 |
|
|
|
|
|
| |
| P1 | 823–910, 880 | 949–1102, 1031 | 549–575, 560 | 333–351, 341 | 240–269, 256 |
| P2 | 703–858, 776 | 757–865, 827 | 381–408, 392 | 227–240, 210 | 171–176, 172 |
| P3 | 683–792, 752 | 816–913, 875 | 628–678, 652 | 408–440, 428 | 289–315, 300 |
|
|
|
|
|
| |
| P1 | 134–150, 141 | 77–124, 105 | 0.52–0.58, 0.55 | 1.67–2.08, 1.87 | 3.23–3.50, 3.41 |
| P2 | 108–125, 115 | 96–99, 97 | 0.45–0.50, 0.48 | 3.04–3.47, 3.23 | 3.83–4.44, 4.08 |
| P3 | 181–205, 191 | 103–126, 115 | 0.71–0.77, 0.75 | 2.15–29, 2.20 | 2.39–2.63, 2.50 |
| fe | ti | ta1 | ta2 | ta3 | ta4 | ta5 | LR | BV | SV | |
|---|---|---|---|---|---|---|---|---|---|---|
| P1 | 873 | 1060 | 551 | 330 | 268 | 164 | 122 | 0.52 | 2.81 | 3.51 |
| P2 | 800 | 880 | 387 | 243 | 211 | 144 | 128 | 0.44 | 2.85 | 4.34 |
| P3 | 832 | 973 | 702 | 440 | 373 | 246 | 146 | 0.72 | 2.45 | 2.57 |
| 1 | Acrostichals present and distinct; tarsomere 5 dorsoventrally flattened |
|
| – | Acrostichals absent; tarsomere 5 laterally fattened |
|
| 2 | Shorter spur of mid tibia either about 2/3 as long as longest spur or lacking, anal point short to moderately long |
|
| – | Shorter spur of mid tibia at most 1/3 as long as longest spur, anal point vestigial to short and strong |
|
| 3 | Degraded anal point, and AR > 2.0 |
|
| – | Anal point is short and rounded at the tip, AR 1.66–1.78 |
|
| 4 | AR about 1.30, Mesonotum with traces of two shortened and darker lateral bands |
|
| – | AR > 1.67, Mesonotum not as above |
|
| 5 | HR < 1.70, gonostylus curves inward from the end 1/3 |
|
| – | HR > 1.70, gonostylus does not curve inward from the end 1/3 |
|
| 6 | Abdomen yellowish brown, uniform color, tergiet XI left and right sides with reticulate pattern |
|
| – | Abdomen II–V yellow, abdomen VI–VIII brown, tergiet XI left and right sides without reticulate pattern |
|
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsFreshwater macroinvertebrate diversity and ecology · Environmental DNA in Biodiversity Studies · Aquatic Invertebrate Ecology and Behavior
Introduction
Kieffer (1906) erected the genus Psectrocladius with Orthocladiuspsilopterus Kieffer, 1906 as the type species. Psectrocladius occurs in a wide variety of aquatic biotopes, predominantly in standing or slow-flowing waters (Cranston et al. 1989). Adults of the genus Psectrocladius can be distinguished from other Orthocladiinae by their combinations of bare eyes, well-developed pulvilli, and straight Cu_1_ (Cranston et al. 1989). The pupae of this genus are unique due to their long posterior spines on tergites V and VIII, the absence of frontal setae and pedes spurii B, and their differentiation from other Orthocladiinae with a thoracic horn, well-developed fringe, and at least one lamelliform (or strong) L seta (Coffman et al. 1986). In addition, Psectrocladius are relatively uniform and distinctive in their larval stage, characterized notably by the palmate structure of S1, which is unique, as well as by the combination of a large ventromental plate, a simple premandible, and spurs on the procercus (Andersen et al. 2013).
Through comprehensive morphological studies of larval-adult associations, Wülker (1956) established a taxonomic framework for the genus Psectrocladius, proposing its division into three distinct subgenera: Psectrocladius s. str. Kieffer, Monopsectrocladius Wülker, and Allopsectrocladius Wülker. This revision was accompanied by a diagnostic key for the identification of male adults. Subsequently, Roback (1957) expanded our understanding of the genus through detailed ontogenetic studies, describing three previously unrecognized species. These taxonomic descriptions were particularly significant as they included comprehensive morphological characterizations of female imagines, pupal exuviae, and larval instars, thereby providing a complete life history perspective for these newly discovered taxa.
The taxonomic understanding of Psectrocladius has been progressively refined through a series of significant contributions. Sæther (1969) made notable advancements through his examination of Canadian specimens from Walden Lake, documenting three species, including one novel taxon. Laville (1971), following meticulous examination of type specimens of Psectrocladiusbarbatipes Kieffer, 1923, made a crucial taxonomic determination. His analysis revealed that the larval morphology of this species exhibited intermediate characteristics between the P.psilopterus and P.dilatatus species groups, which correspond to the subgenera Psectrocladius and Allopsectrocladius, respectively. This morphological intermediacy warranted the establishment of a new subgenus, Mesopsectrocladius Laville. In the United Kingdom, Pinder (1978) significantly contributed to the regional taxonomy by developing a comprehensive identification key for British Psectrocladius species, encompassing nine species distributed across the three established subgenera: Psectrocladius, Monopsectrocladius, and Allopsectrocladius. Langton (1980) further enhanced diagnostic capabilities by providing distinct subgeneric keys for both pupal and adult stages, along with specific keys for the subgenus Psectrocladius, covering eight species. This work was subsequently supplemented by Langton’s (1985) revisionary study of Psectrocladiuslimbatellus and associated specimens. Parallel research efforts in Russia and Japan have substantially expanded the known diversity of this genus. Russian researchers (Zelentsov 1980; Zelentzov and Makarchenko 1988; Makarchenko and Makarchenko 2003) documented four species, while Japanese investigations (Yamamoto 2004) significantly increased this number with records of fourteen species, demonstrating the genus’s extensive distribution and morphological diversity across the Palaearctic region.
The study of Psectrocladius in China has evolved through several significant contributions since the initial taxonomic work. Kieffer (1923) provided the first record of this genus in China with the description of Psectrocladiusformosae Kieffer, 1923 from Taiwan Province. Subsequent research by Wang and Zheng (1996) expanded our knowledge through the description of Psectrocladiuslongipennis Wang et Zheng, 1996, based on specimens collected from Qinghai Province. Wang (2000) further documented the genus’ diversity in China, reporting several unidentified taxa along with two adult specimens, one larval specimen, and two records requiring taxonomic verification. Recent investigations in Xizang Autonomous Region have yielded important records of immature stages, with Ge et al. (2024) documenting both larval and pupal specimens of Psectrocladiusnevalis Akhrorov, 1977.
Current taxonomic understanding recognizes seven Psectrocladius species in China, with complete adult descriptions available for only four taxa: P.formosae Kieffer, 1923, P.longipennis Wang et Zheng, 1996, P.obvius (Walker, 1856), and P.sokolovae Zelentzov et Makarchenko, 1988. The remaining species - P.limbatellus (Holmgren, 1869), P.nevalis Akhrorov, 1977, and P.barbimanus (Edwards, 1929) - are known from either immature stages or incomplete records (Ashe and O’Connor 2012; Wang et al. 2020; Ge et al. 2024). The present study significantly contributes to the Chinese Psectrocladius fauna through the description and illustration of three new species. Additionally, we provide a comprehensive diagnostic key for the identification of adult males of all known Psectrocladius species recorded in China, facilitating future taxonomic studies in this region.
Material and methods
The morphology and terminology are based on Sæther (1980). The material examined was mounted on slides using the procedure outlined by Sæther (1969). When three or more specimens were measured, the measurements are provided as the range and mean, with the number of observed specimens in parentheses if it differs from the number (N) stated at the beginning of the description. Color descriptions pertain to ethanol-preserved specimens. The holotype of all new species is deposited at the College of Life Sciences, Tianjin Normal University, Tianjin, China (TJNU).
Taxonomy
Psectrocladius (Mesopsectrocladius) wangi
Taxon classificationAnimaliaDipteraChironomidae
Liu sp. nov.
5EBC6A25-0D60-586C-90EF-41A99B49B30A
https://zoobank.org/D8057D4F-8A80-44BB-BF8A-7040EC8EBB2A
Material examined.
Holotype: • male (TJNU No. 04724), China, Fujian Province, Fujian Agriculture and Forestry University, 26°05'17"N, 119°18'43"E, 22.IV.1993, X.H. Wang, light trap. Paratype: • two male, same data as holotype; one male, China, Guizhou Province, Fanjing Mountain protection temple, 27°55'N, 108°41'E, 28.V.2002, R.L. Zhang, light trap; • one male, China, Fujian Province, Shanghang County, Gutian Town. 25°13'28"N, 116°49'23"E, 4.V.1993. X. H. Wang, sweep.
Diagnosis.
The anal tergite is wider at the top and narrower at the bottom with a rounded posterior margin. The anal point is short and rounded at the tip, thumb-like. The femur setae are thick, resembling spines.
Description.
Male (N = 5, unless otherwise stated in brackets).
Total length 3.37–3.88, 3.56 mm. Wing length 1.78–2.10, 1.92 mm. Total length/wing length 1.75–1.96, 1.86. Wing length/length of profemur 2.08–2.28, 2.23. The thorax is yellow-brown with brown markings, scutellum and the posterior half of the scutellum are lighter in color. The abdomen is brown.
Head (Fig. 1C). AR 1.66–1.78, 1.72. Temporal setae 14–17, 15; including 3–7, 6 inner verticals; 4–9, 6 outer verticals; and 6–10, 8(3) postorbitals. Clypeus with 21–37, 29 setae. Tentorium 139–172, 160 μm long. Palpomere lengths (II–V in μm): 51–75, 62; 81–117, 94; 78–107, 91; 121–151, 137; The ratio of the length of V to III: 1.29–1.62, 1.43.
Psectrocladius (Mesopsectrocladius) wangi Liu, sp. nov., holotype male A wing B thorax C head D antenna E foreleg F midleg G hindleg H tarsus V. Scale bars: 100 μm (A, B, C, D, E, F, G); 20 μm (H).
Thorax (Figs 1B, 3B). Antepronotals with 5–12, 9 setae, acrostichals 5–7, 6, dorsocentrals 17–28, 23. prealars 4–8, 5. Scutellum with 6–12, 10 setae.
Wing (Figs 2A, 3A). Anal lobe developed. VR 1.17–1.21, 1.19. Costa extension 36–49, 43 µm. The end of R_2+3_ is between R_1_ and R_4+5_. Radius with 8–15, 11 setae. R_1_ with 2–6, 4 setae. Squama with 13–17, 16 setae. Brachiolum with one seta.
Legs (Fig. 1E–G). Tarsomeres without bristles, hind leg with long bristles. Front tibia with one spur, 75–85, 81 µm long. Mid tibial with 2 spurs, the long one 53–60, 58 µm and the other is one-third of it, 20–25, 23 µm. Hind tibia with one spur, 70–80, 73 µm long. The hind tibial comb with 10–14, 12 spurs. Tarsus I and II of mid leg with two pseudospurs. Tarsus III of mid legs with 0–1 pseudospurs. Tarsus I and II of hind leg with two pseudospurs. Tarsus III of hind legs with 0–1 pseudospurs. Lengths (in μm) and proportions of legs as in Table 1.
Hypopygium (Figs 2A, C, D, 3C, D). Tergite IX with 14–18, 16 setae located in the lower part of anal tergite equably. Anal tergite wider at the top and narrower at the bottom, and margo inferior smooth. The end of anal point smooth and as wide as the base of anal point, thumb-like. Transverse sternapodeme 125–150, 129 µm long centrally slightly arched, with prominent oral projections. Virga 73–85, 75 µm long, club-shaped. The dorsal volsella is salient, the ventral volsella is arc-shaped and bears a row of setae. Gonocoxite 272–315, 287 µm long, Gonostylus 111–148, 126 µm long, outer margin with conspicuous setal fringe. Megaseta 15–30, 24 µm long. HR 2.13–2.51, 2.46; HV 2.62–3.19. 2.85.
Psectrocladius (Mesopsectrocladius) wangi Liu, sp. nov., holotype male A hypopygium B abdomen C volsella D anal point. Scale bars: 100 μm (A, B); 20 μm (C, D).
Psectrocladius (Mesopsectrocladius) wangi Liu, sp. nov., holotype male A wing B thorax C hypopygium, dorsal view D hypopygium, ventral view.
Distribution.
China (Fujian, Guizhou).
Etymology.
Named after Xinhua Wang, in honor of his contributions to the study of Chironomidae, noun in nominative case.
Remarks.
The subgenus Psectrocladius (Mesopsectrocladius) currently comprises two recognized species globally (Ashe and O’Connor 2012): P. (M.) barbatipes Kieffer, 1923 and P. (M.) seiryuheius Sasa, Suzuki & Sasai, 1999. These taxa exhibit distinct morphological characteristics that facilitate their differentiation. Psectrocladius (M.) seiryuheius is particularly notable for the complete absence of both anal point and virga, representing a unique apomorphic condition within the subgenus.
Comparative morphological analysis reveals that P. (M.) barbatipes shares certain similarities with the newly described species. However, it can be distinguished by several key characteristics: (1) the presence of a less developed and partially reduced anal point, and (2) a significantly higher antennal ratio (AR > 2.0, following Laville’s 1971 description). Furthermore, P. (M.) barbatipes exhibits distinctive chaetotaxy on its mid legs, characterized by exceptionally long setae, as evidenced by its high bristle ratio (BR > 3.0). This morphological feature suggests an adaptation potentially related to specific ecological requirements or behavioral patterns.
Psectrocladius (Psectrocladius) gracilis
Taxon classificationAnimaliaDipteraChironomidae
Liu sp. nov.
D44E2125-085A-5BD2-8114-D456E05467BA
https://zoobank.org/1FFE46C2-ED89-4F81-959D-7F95D38E4FCC
Material examined.
Holotype: • male (TJNU No.1210), China, Inner Mongolia Autonomous Region, Ulansuhai Nur, 40°55'19"N, 108°50'66"E, IV.1982, X.H. Wang. Sweep. Paratypes: • two males, same data as holotype.
Diagnosis.
Thorax and abdomen dark. Anal tergite inverted triangle, both sides with a reticulate pattern; anal point mid-length, uniform thickness, gonostylus narrow and long.
Description.
Male (N = 3, unless stated).
Total length 3.63–3.66, 3.65 mm. Wing length 2.00, 2.15 (2) mm. Total length/wing length 1.69, 1.82 (2). Wing length/length of profemur 2.43, 2.36 (2). Thorax dark brown to black, abdomen yellowish brown (Fig. 5B).
Head (Fig. 4B). AR 1.67–1.97, 1.80. Temporal setae 10–12, 11; including 2–4, 3 inner verticals; 3–6, 4 outer verticals; and 2–5, 4 postorbitals. Clypeus with 9–11, 10 setae. Tentorium 150–164, 155 μm long. Palpomere lengths (II–V in μm): 48–62, 56; 86–107, 97; 110–130, 113; 131–160, 148. The ratio of the length of V to III: 1.35–1.86, 1.55.
Psectrocladius (Psectrocladius) gracilis Liu, sp. nov., holotype male A wing B head C thorax D foreleg E midleg F hindleg G tarsus V. Scale bars: 100 μm (A, B, C, D, E, F); 20 μm (G).
Thorax (Figs 4C, 6B). Antepronotals 4–10, 7, acrostichals absent, dorsocentrals 10–16, 13, prealars 4–8, 6. Scutellum with 4–6, 5 setae.
Wing (Fig. 4A, 6A). Anal lobe developed. VR 1.16, 1.23 (2). Costa extension 55, 58 (2) µm. The end of R_2+3_ is between R_1_ and R_4+5_. Radius with 5 (1) setae. Squama with 29 (1) setae. Brachiolum with one seta.
Legs (Fig. 4E–G). Front tarsomeres without bristles, tibial comb with one spur, 68–83, 74 µm long, mid tibia with two spurs, one is 53–55, 53 µm long, one is thin and small, 8–10, 9 µm long. Hind tibia with one spur, 70–75, 73 µm long. The tibial comb of mid legs with 13–17, 14 spurs. Tarsus I of mid legs with two pseudospurs. Tarsus II of mid legs with 1–2 pseudospurs. Tarsus III of mid legs with two pseudospurs. Tarsus I and II of hind legs with two pseudospurs. Lengths (in μm) and proportions of legs as in Table 2.
Hypopygium (Figs 5A, C, D, 6C, D). Tergite IX with 13–19, 15 setae ascending along both sides of the base of anal point. Tergite paratergital 10–15, 12 setae. Anal tergite inverted triangle, both sides with a reticulate pattern. Anal point mid-length, 20–29, 25 µm long, uniform thickness. Transverse sternapodeme 115–138, 124 µm long, central slightly arched, with well-developed bilateral ossified processes. Virga 75–80, 78 µm, the top of virga a small part is hook-shaped. Dorsal volsella square-shaped, ventral circular. Both sides of volsella with setae. Gonocoxite 250–276, 266 µm. Gonostylus 117–130, 123 µm, megaseta 12–17, 15 µm. HR 2.08–2.26, 2.16; HV 2.79–3.11, 2.97.
Psectrocladius (Psectrocladius) gracilis Liu, sp. nov., holotype male A hypopygium B abdomen C anal point D volsella. Scale bars: 100 μm (A, B); 20 μm (C, D).
Psectrocladius (Psectrocladius) gracilis Liu, sp. nov., holotype male A wing B thorax C hypopygium dorsal view D hypopygium ventral view.
Distribution.
China (Inner Mongolia).
Etymology.
From the Latin, gracilis, narrow and thin, referring to the character of tergite IX and gonostylus, adjective in the nominative singular.
Remark.
The newly described species, Psectrocladius (Psectrocladius) gracilis Liu, sp. nov., exhibits significant morphological affinities with P. (P.) jintuoctadecimus (Sasa, 1996) in several diagnostic characters. These shared characteristics include: (1) the structural configuration of the inferior volsella, (2) the distinctive reticulate patterning on the lateral aspects of tergite XI, and (3) comparable measurements of both antennal ratio (AR) and leg ratio (LR). However, P. (P.) gracilis Liu, sp. nov. can be readily distinguished by its elongate and slender anal point and gonostylus, which contrast markedly with the nearly triangular morphology observed in P. (P.) jintuoctadecimus (Sasa, 1996), particularly in the latter’s characteristic broad-tipped gonostylus that significantly exceeds its basal width.
Furthermore, while P. (P.) gracilis Liu, sp. nov. shares certain anal point characteristics with P. (P.) limbatellus (Holmgren, 1869), the two species are clearly differentiated by their gonostylus morphology. The gonostylus of P. (P.) gracilis Liu, sp. nov. is diagnostically characterized by its notably narrow and attenuated structure, representing a distinct morphological divergence from the gonostylus configuration observed in P. (P.) limbatellus.
Psectrocladius (Psectrocladius) malum
Taxon classificationAnimaliaDipteraChironomidae
Liu sp. nov.
6BEB00D7-86BE-53A0-BA01-0A05B7D4542E
https://zoobank.org/07A8C464-9E51-408C-BE83-7764045EBBA4
Material examined.
Holotype: • male (TJNU No.12521), China, Inner Mongolia Autonomous Region, Xilinhot City, 43°57'29"N, 116°03'41"E, 1020 m, 10.VIII.1997, H.H. Li, sweep.
Diagnosis.
The base of gonocoxite with a small convex, the mid-upper part of gonocoxite with obvious concave, the outlines of both the left and right sides create a shape reminiscent of an apple. The end of inferior volsella pendulous and tongue-shaped.
Description.
Male (N = 1).
Total length 4.07 mm. Wing length 2.04 mm. Total length/wing length 2.00. Wing length/length of profemur 2.32. Thorax yellow to brown, with dark brown patterns. Abdomen II–V yellow, abdomen VI–VIII yellowish brown.
Head (Fig. 7B). AR 1.70. Temporal setae 13; including 4 inner verticals; 3 outer verticals; and 6 postorbitals. Clypeus with 16 setae. Tentorium 116 μm long. Palpomere lengths (II–V in μm): 54, 102, 89, 203. The ratio of the length of V to III: 2.00.
Psectrocladius (Psectrocladius) malum Liu, sp. nov., holotype male A wing B head C thorax D foreleg E midleg F hindleg G tarsus V. Scale bars: 100 μm (A, B, C, D, E, F); 20 μm (G).
Thorax (Figs 7C, 9B). Antepronotum with 7 setae, acrostichals absent, dorsocentrals 6, prealars 6. Scutellum two setae.
Wing (Figs 7A, 9A). Anal lobe developed. VR 1.17. Costa extension 43 µm. The end of R_2+3_ is between R_1_ and R_4+5_. Radius with 9 setae. R_1_ with one seta. Squama with two setae. Brachiolum with two setae.
Legs (Fig. 7E–G). Front legs, mid legs and hind legs each with one tibial spur, length is 70, 55, 65 µm. The tibial comb of hind legs with 17 spurs. Tarsus I of fore legs with one pseudospurs. Tarsus I, II and III of mid legs with two pseudospurs. Tarsus I, II and III of hind legs with two pseudospurs. Lengths (in μm) and proportions of legs as in Table 3.
Hypopygium (Figs 8A, C, D, 9C, D). Tergite IX with 15 setae located two sides and the base of anal point. Tergite paratergital 11 setae. Anal point 14 µm long. The base of anal point 8 µm wide. Transverse sternapodeme 128 µm long, central slightly arched, with well-developed bilateral ossified processes, triangle-shaped. Virga 53 µm long. The end of inferior volsella pendulous and tongue-shaped. Outer margin of inferior volsella semicircular. Gonocoxite 303 µm, the base of gonostylus with a small convex, base outer margin of gonocoxite concave, the left and right contours form apple shaped. Gonostylus 166 µm, Megaseta 18 µm. HR 1.83; HV 2.45.
Psectrocladius (Psectrocladius) malum Liu, sp. nov., holotype male A hypopygium B abdomen C anal point D volsella. Scale bars: 100 μm (A, B); 20 μm (C, D).
Psectrocladius (Psectrocladius) malum Liu, sp. nov., holotype male A wing B thorax C hypopygium, dorsal view D hypopygium, ventral view.
Distribution.
China (Inner Mongolia).
Etymology.
From the Latin, malum, apple, referring to the convex and concave upper inner margin of the two sides of the gonocoxite, which has an apple-like form.
Remarks.
The gonocoxite morphology provides crucial diagnostic characters for distinguishing Psectrocladius species. Psectrocladius (Psectrocladius) malum Liu, sp. nov. is characterized by a unique concave inner margin at the base of its gonocoxite, representing a distinctive apomorphic feature. This condition contrasts markedly with P. (P.) semicirculatus Sæther, 1969, which exhibits a more complex gonocoxite structure featuring both concave and convex inner margins. Furthermore, P. (P.) malum Liu, sp. nov. can be unequivocally differentiated from P. (P.) semicirculatus by its distinctive tergite XI morphology and anal point configuration.
Comparative analysis reveals that the anal point morphology of P. (P.) malum Liu, sp. nov. shows notable similarities with that of P. (P.) sokolovae Zelentzov & Makarchenko, 1988. However, these species can be readily distinguished by several quantitative characters: P. (P.) sokolovae possesses (1) a significantly higher antennal ratio (AR) and (2) a greater overall body length. Additionally, while P. (P.) barbimanus (Edwards, 1929) also displays a slight concavity on the outer margin of the gonocoxite base, this species is immediately recognizable by its distinctive pear-shaped overall morphology, providing a clear diagnostic feature for species identification.
Key to males of Psectrocladius Kieffer from China
**: **
Supplementary Material
XML Treatment for Psectrocladius (Mesopsectrocladius) wangi
XML Treatment for Psectrocladius (Psectrocladius) gracilis
XML Treatment for Psectrocladius (Psectrocladius) malum
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Andersen TSæther OA Cranston PS Epler JH (2013) The larvae of Orthocladiinae (Diptera: Chironomidae) of the Holarctic Region – Keys and diagnoses. In: Andersen T Cranston PS Epler JH (Eds) The larvae of Chironomidae (Diptera) of the Holarctic Region – Keys and diagnoses.Insect Systematics & Evolution 66 (Supplement), 189–385.
- 2Ashe PO’Connor JP (2012) A world catalogue of Chironomidae (Diptera). Part 2. Orthocladiinae.Irish Biogeographical Society & National Museum of Ireland, Dublin, 968 pp.
- 3Coffman WP Cranston PS Oliver DRSæther OA (1986) The pupae of Orthocladiinae (Diptera: Chironomidae) of the Holarctic Region – keys and diagnoses. In: Wiederholm T (Ed.) Chironomidae of the Holarctic Region – Keys and diagnoses.Part 2. Pupae. Entomologica Scandinavica 28(Supplement), 148–299.
- 4Cranston PS Dillon ME Pinder LCV Reiss F (1989) The adult males of Chironominae (Diptera: Chironomidae) of the Holarctic Region – keys and diagnoses. In: Wiederholm T (Ed.) Chironomidae of the Holarctic Region – Keys and diagnoses. Part 3. Adult males.Entomologica Scandinavica, Supplement 34: 165–352.
- 5Edwards FW (1929) British non-biting midges (Diptera, Chironomidae).Transactions of the Royal Entomological Society of London 77: 279–430. 10.1111/j.1365-2311.1929.tb 00692.x · doi ↗
- 6Ge X Wang C Pei W Tang Y Liu W Yan C (2024) New descriptions of the larval and pupal stages of Orthocladiusnitidoscutellatus and Psectrocladiusnevalis from Xizang, China (Diptera, Chironomidae). Biodiversity Data Journal 12: e 121952. 10.3897/BDJ.12.e 121952 PMC 1101616138617833 · doi ↗ · pubmed ↗
- 7Kieffer JJ (1906) Description d’un genre nouveau et de quelques espèces nouvelles de Diptères de l’Amérique du Sud.Annales de la Société Scientifique de Bruxelles 30: 349–358.
- 8Kieffer JJ (1923) Nouvelle contribution à l’étude des Chironomides de la Nouvelle-Zemble.Report on the Scientific Results of the Norwegian Expedition to Novaya Zemlya 9: 3–11.