Genomic epidemiology of vancomycin-resistant Enterococcus faecium in Eastern Denmark from 2020 to 2022, and identification of vanB Tn1549 insertion sites
Maja Johanne Søndergaard Knudsen, Jose Alfredo Samaniego Castruita, Ingrid Maria Cecilia Rubin, Sarah Mollerup, Helle Krogh Johansen, Rasmus L. Marvig, Karen Leth Nielsen, Barbara Juliane Holzknecht, Morten Hoppe, Michael Kemp, Henrik Westh, Mette Pinholt

TL;DR
This study analyzed vancomycin-resistant Enterococcus faecium in Denmark from 2020 to 2022, identifying new genomic patterns and Tn1549 insertion sites.
Contribution
The study reports ten new chromosomal Tn1549 insertion sites in vanB VREfm isolates from Denmark.
Findings
Most vanB isolates had Tn1549 inserted in araA2 or sir2 genes.
Eight new chromosomal insertion sites were identified.
Three isolates had Tn1549 on plasmids.
Abstract
We aimed to describe the genomic epidemiology of vancomycin-resistant Enterococcus faecium (VREfm) in Eastern Denmark from 2020 to 2022, identify and characterise the vanB Transposon 1549 (Tn1549) insertion sites among vanB VREfm clones and identify emerging VREfm clones. We analysed all VREfm from our routine diagnostic sequencing during the study period. Using the Seqsphere + v.8.2.0 software (Ridom GmbH, Münster, Germany, (http://www.ridom.de/seqsphere), minimum spanning trees were created to visualise clusters. Tn1549 insertion sites were determined by in silico PCR. Nanopore sequencing was performed to assemble the regions surrounding Tn1549, which helped determine the insertion site locations. We included 2,437 isolates in the study. A total of 463 isolates carried vanA, 1,963 isolates carried vanB, and 11 isolates carried both genes. Of all isolates carrying vanB, 254 isolates…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
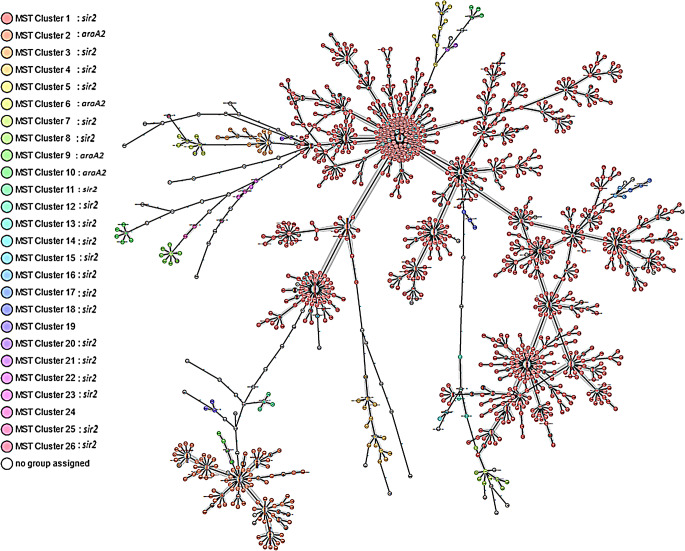 Figure 1
Figure 1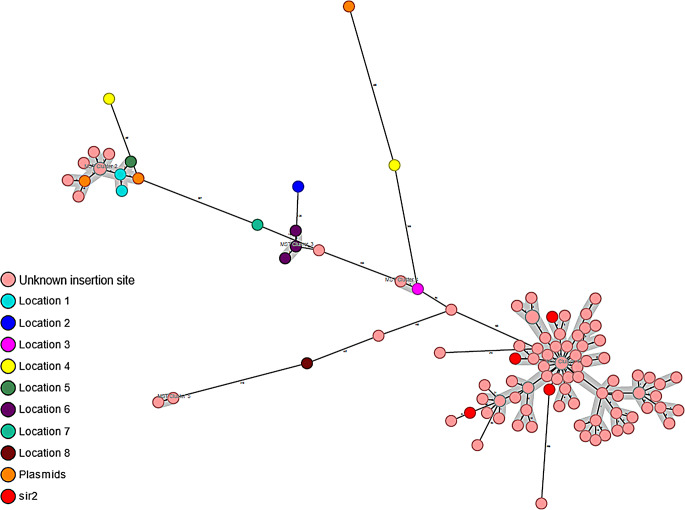 Figure 2
Figure 2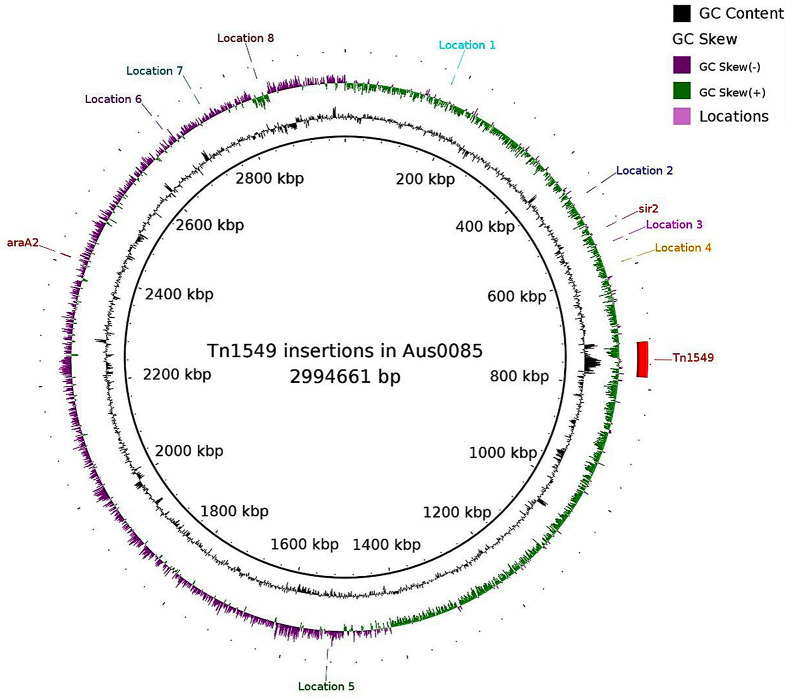 Figure 3
Figure 3- —Copenhagen University
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsAntimicrobial Resistance in Staphylococcus · Streptococcal Infections and Treatments · Bacterial biofilms and quorum sensing
Introduction
Enterococcus faecium can cause nosocomial infections associated with intravenous catheter, invasive infections in the abdomen, and bacteraemia [1]. A Danish cohort study investigated enterococcal bacteraemia and found the incidence of E. faecium bacteraemia to be 6.6/100,000 person-years, and the 30-day mortality in this population to be 34.6% [2]. In another Danish study, the authors founds no difference in the 30-day mortality between patients with vancomycin-resistant E. faecium (VREfm) bacteraemia, and patients with vancomycin-susceptible E. faecium bacteraemia (37.6% vs.37.0%) [1]. Te in-hospital mortality rate of VREfm infections is 18%, risin to above 69% at 90 ays [3–5].
Vancomycin-resistance in E. faecium from human isolates is mainly caused by the acquisition of vanA or vanB. Studies have shown that the van operons can be transmitted between E. faecium strains by horizontal gene transfer [6, 7]. In this study, we focus on the vanA gene, which is located on Transposon 1546 (Tn1546) and most often carried on plasmids, and the vanB gene, which is located on Transposon 1549 (Tn1549), inserted in the E. faecium chromosome, or rarely on plasmids [8, 9].
From 2013, Denmark observed an increase in vancomycin-resistant E. faecium (VREfm) due to the emergence of several vanA VREfm clones. From 2019, vanB VREfm started to increase and outcompeted vanA VREfm [10, 11].
The emergence of vanB VREfm has challenged rapid diagnostic of VREfm from rectal screening samples by PCR since vanB is present in multiple gut commensals other than E. faecium. As previously described, a multiplex PCR, targeting both vanB, and the specific Tn1549 insertion sites in the chromosome in the two most dominating vanB clones (a L-arabinose isomerase gene (araA2), and a silent information regulator gene (sir2) in our area, was developed at the DCM Hvidovre Hospital to overcome this diagnostic challenge [8, 12].
Accurate and rapid diagnostic of VREfm performed directly on rectal swabs (within 12 h) is crucial to maintain infectious hygiene precautions in a hospital setting, and to prevent hospital outbreaks [13]. In our regions, we carry out contact isolation precautions for all patients that are carriers of or infected with VREfm. In one of the regions’ four DCMs (Hvidovre Hospital) we use the results from the multiplex PCR to determine which VREfm clone patients are colonised or infected with. This allows for timely analysis of VREfm dissemination in hospital wards. In addition, when we need to perform cohort isolation in case of lack of single rooms in the wards, we use the results from the multiplex PCR. In those cases, we perform cohort isolation of patients that have the same van gene and Tn1549 insertion site on the multiplex PCR. We use VREfm whole genome sequencing (WGS) for continuous surveillance of VRE on a regional level. This is important to identify the VREfm clone-distribution in our hospitals and emergence of new clones that require diagnostic changes.
The aims of this study were to describe the genomic epidemiology of VREfm in Eastern Denmark, from 2020 to 2022 and to identify emergence of new clones, especially vanB clones with non-araA2/non-sir2 Tn1549 insertion sites. Furthermore, we aimed to identify and characterise the Tn1549 insertion sites among vanB VREfm isolates in our collection to evaluate how many different Tn1549 insertion sites that are present in our vanB VREfm collection and to determine the proportion of vanB VREfm that will be identified by the laboratory developed multiplex-PCR.
Methods
Setting and collection of data
In Eastern Denmark we perform routine diagnostic sequencing of one VREfm isolate per patient per year. In the cases, where patients have samples positive for both vanA and vanB genes within the same year, we sequence both isolates in line with our active surveillance strategy. The routine sequencing applies to both screening and clinical isolates. Infection prevention and control measures include isolation of VREfm positive patients and screening of patients from the same room, whenever VREfm is found in a sample from a patient that was not isolated. If a patient from the same room has a positive sample for VREfm, we screen all patients on the ward. In periods with VREfm outbreaks, we perform weekly screenings. We screen patients at admission if they have had a VREfm positive sample within the past six months. In this retrospective study we collected all sequences from VREfm isolates from January 2020 until June 30, 2022, from Eastern Denmark (Region of Zealand and the Capital Region of Denmark with 2.7 million inhabitants, approximately 50% of the Danish population). We have four departments of clinical microbiology (DCMs), all of which have collected isolates that are represented in the study, and the DCMs serve a total of 15 hospitals as well as all general practitioners in the area.
Whole genome sequencing and bioinformatical methods
We used the Nextera XT DNA Sample Preparation Kit (Illumina, San Diego, California, U.S.A) to prepare libraries, and the isolates were sequenced either on the Illumina MiSeq or the NextSeq platform (Illumina Inc., San Diego, USA) in paired end reads. The sequences were assembled with SKESA v.2.2 or SPADES depending on the DCM [14, 15]. We used quality control parameters, and we included only isolates with N50 above 10,000, and genome sizes between 2.7 and 3.3 million base pairs. We used the Seqsphere + v.8.2.0 software (Ridom GmbH, Munster, Germany, http://www.ridom.de/seqsphere/) to analyse all isolates based on the formerly published scheme for E. faecium consisting of 1,423 core genome Multi Locus Sequence Type (cgMLST) target genes [16, 17]. We created Minimum Spanning Trees (MST) to visualise cgMLST CT clusters with the setting of maximum 20 alleles between isolates. Genome annotation was performed in the assemblies using Prokka v.1.14.5, and the annotating proteins in Aus0085 [18, 19]. Regions surrounding Tn1549 were blasted using ncbi-blast v.2.8.1 [20]. Insertions sites were visualised using BRIG v.0.95, R v.4.3.0 and gggenes library [21–23].
Analysis of the Tn1549 insertion sites
We used the multiplex PCR primer sets to perform in silico PCR on all vanB isolates to determine which isolates belonged to the dominating Tn1549 insertion sites in araA2 or sir2 genes in the chromosome [8, 12]. Subsequently, we analysed the non-araA2/non-sir2 isolates. The regions surrounding Tn1549 were used to predict the inserted sites in these isolates. Based on the MST, we proceeded with nanopore sequencing on 12 representative isolates without the Tn1549 insertion site in araA2 or sir2 to construct a better assembly of the genome and the insertion sites (Table 1).
Nanopore sequencing
Genomic DNA libraries were built using the Rapid Barcoding Kit and sequenced using a MinION (Oxford Nanopore Technology, Oxford, UK) with a R9.4.1 flow cell. Hybrid assemblies were done using the Illumina and Nanopore reads with Unicycler [24]. Mauve was used to compare the hybrid assemblies and inspect genome rearrangements [25]. Visualisation of the genomes was done by BLAST Ring Image Generator (BRIG) [26].
Results
Genomic epidemiology of VanA and VanB
In total 2,437 VREfm isolates were included in the study. vanB VREfm was dominant with 1,963 (81%) isolates; however, vanA was still present with 463 isolates (19%). Eleven isolates carried both vanA and vanB. Most of the isolates in the vanB and vanAB subgroup belonged to ST80/CT2406 (n = 1,614, 82%/n = 1,180, 60%). The MST of vanB and vanAB isolates is presented in Fig. 1, where cluster 1 is mainly represented with ST80 isolates.
Fig. 1. Shows the minimum spanning tree of the vanB and vanAB isolates. The isolates are coloured according to the clustering of the isolates by the core genome multilocus sequence types. The vast majority of the cluster 1 isolates has the Tn1549 inserted in sir2, and the vast majority of the isolates in cluster 2 has the Tn1549 inserted in araA2
The most prevalent Sequence Types (ST) and cgMLST Complex Types (CT) in vanA and vanAB carrying isolates were the ST1421/CT1134 (n = 307, 65%/n = 305, 64%). The dominating ST1421 clone was the vancomycin-variable clone that was introduced to our area in the 2016 [11, 27]. The vanA carrying isolates are presented in the supplementary figure, where most of the ST1421/CT1134 are found in cluster 1. The 11 isolates carrying both vanA and vanB was a diversified group that belonged to four different ST, and five different CT.
Tn1549 insertion site
Of the 1,974 vanB isolates, 254 (13%) isolates had the Tn1549 inserted in araA2, and 1,604 (81%) isolates had the Tn1549 inserted in the sir2. Thus, the multiplex PCR targeting vanB and the araA2/sir2 insertion sites identifies 94% of the vanB VREfm isolates in VREfm from Eastern Denmark. We found the sir2 insertion sites in 20 different MST clusters including the biggest cluster 1. The araA2 insertion site was found in four different MST clusters including the cluster 2 (Fig. 1).
In total, we failed to find a complete in silico PCR product in 116 samples, either because they had a non-araA2/non-sir2 insertion site, or incomplete PCR products were present on the contigs (Fig. 2). In these samples, ST80/CT2406 (n = 84, 72.4%/n = 64, 55.1%) were most frequent, as was the case for the vanB isolates with the sir2 insertion site. None of these isolates had undergone the routine analysis with the multiplex PCR detecting both araA2 and sir2 insertion sites.
Fig. 2. Shows the minimum spanning tree of the non-araA2/non-sir2 isolates. The isolates are coloured according to the Tn1549 insertion sites determined by analysis of contigs assembled from Illumina data (7 isolates), or hybrid assembly with Illumina and Nanopore data (12 isolates). The isolates with Tn1549 insertion site in sir2 (red) are all located in the biggest MST cluster
VREfm clones with non-araA2/non-sir2 Tn1549 insertion sites
We were able to identify the Tn1549 insertion sites in 19 of the 116 non-araA2/non-sir2 isolates using Nanopore and Illumina sequencing (12 with Nanopore, and 7 with Illumina). There were eight different predicted locations where Tn1549 was inserted in the genome. In three isolates the transposon was predicted to be located on a plasmid. All non-araA2/non-sir2 isolates are presented in the MST in Fig. 2, where the isolates are coloured according to the insertion site. The location of the insertion sites identified in 19 isolates is shown on Fig. 3 and in Table 1. In the Table 1, we have marked the isolates that have undergone Nanopore sequencing. Four isolates turned out to have the insertion site in sir2, although the in silico PCR failed to find a complete PCR product (coloured red in Fig. 2). Three of these isolates had been Nanopore sequenced.
Fig. 3. Shows the location of the araA2 Tn1549 insertion site, the sir2 insertion site, and the eight other chromosomal insertion sites that we have identified, based on the Aus0085 assembly (NC_021994)
Table 1. Overview of the Tn1549 insertion sites based on the Aus0085 assembly, as well as strand (+/-), genes or loci, where Tn1549 has been inserted, the ST, and the CT for each isolate. ^a^Tn1549 is located next to the sir2 gene. ^b^Tn1549 is located next to a genomic rearrangement. ^c^Nanopore and illumina reads were used for the assemblySampleTn1549 insertion siteStrandLocus or geneMLST STcgMLST CTV3969^c^177,141(location 1)-EFAU085_RS00880/EFAU085_RS0088514211134VRE5237177,141(location 1)-EFAU085_RS00880/EFAU085_RS0088514211134V4484^c^467,030(location 2)-EFAU085_RS02205117991VE2908528,269^a^(sir2)+ sir2 Unknown5203VRE5912^c^528,269^a^(sir2)+ sir2 802406VE2293^c^528,269^a^(sir2)+ sir2 804836VE2295^c^2,446,246^a, b^(sir2)+ sir2 802946VE2445554,193(location 3)-EFAU085_RS02565/EFAU085_RS025758971860V3997^c^589,859(location 4)-EFAU085_RS02740/EFAU085_RS02745173115VRE6733589,859(location 4)-EFAU085_RS02740/EFAU085_RS027456122362VE2418^c^1,572,492(location 5)-EFAU085_RS07530/EFAU085_RS0782014211134V38322,672,254(location 6)+EFAU085_RS13235/EFAU085_RS181601171180V3870^c^2,672,254(location 6)+EFAU085_RS13235/EFAU085_RS181601171180VRE52162,672,254(location 6)+EFAU085_RS13235/EFAU085_RS181601171180VRE7066^c^2,745,504(location 7)-EFAU085_RS135951175149RHVRE034^c^2,868,125(location 8)-fss3/EFAU085_RS170901731UnknownVE2234125,278 (plasmid 1)-EFAU085_RS1540014211134VRE5694^c^25,484(plasmid 3)-EFAU085_RS1597014211134VRE6980^c^4705(plasmid 1)+EFAU085_RS14720/EFAU085_RS1473016932532
Using the genome annotations, we could see two variants of the Tn1549 sequence from samples that failed to find a complete araA2 or sir2 in silico PCR product. One of the variants has an ISL3 family transposase inserted in the opposite strand between vanS and vanY. Most samples with the predicted variant with the ISL3 family transposase belong to ST80 (95.8%).
Discussion
In this study we have investigated the genomic epidemiology of VREfm in two regions in the Eastern part of Denmark from 2020 to 2022. We have characterised the Tn1549 insertion sites among the vanB clones and evaluated the coverage of our current multiplex PCR. The majority of the VREfm isolates carried vanB and had the Tn1549 inserted in sir2. Our multiplex PCR correctly identified 94.1% of the vanB isolates. We have identified eight chromosomal insertion sites other than araA2 and sir2. None of these isolates are currently a part of an emerging clone, and there is no evidence suggesting these isolates have an increased fitness in comparison to the isolates with araA2 and sir2 insertion sites in our collection of isolates from 2020 to 2022.
Interestingly, in the subgroup of non-araA2/non-sir2 isolates we discovered some isolates that turned out to have the insertion site in sir2 after reassembling the genomes using Nanopore technology (coloured red on the Fig. 2). These isolates were found in the largest MST cluster of the non-araA2/non-sir2 subgroup. We hypothesize that the isolates in this MST cluster have Tn1549 inserted in the sir2 gene, but they show a negative result in the in silico PCR due to the PCR product being located in two different contigs in their assemblies. Like the sir2 clone, most of the isolates in this MST cluster belonged to ST80/CT2406 [12]. We would hypothesize that these isolates would turn positive in the multiplex PCR, which thereby could identify more than 94% o the vanB isolates in Eastern Denmark.
International colleagues from Germany, the Netherlands, and Australia have also identified the Tn1549 insertion sites in their vanB isolates [28–30]. Two of our isolates have the Tn1549 inserted between gene lacI (GenBank locus tag: EFAU085_RS02740) and a gene encoding a hypothetical protein (GenBank locus tag: EFAU085_RS02740), which is also one of the insertion sites that Zhou et al. and Bender et al. found in their studies [29, 30]. We also found that isolate VRE5694 had the Tn1549 inserted in a plasmid gene that encodes for a recombinase family protein (GenBank locus tag: EFAU085_RS15970) or DNA invertase Pin gene in Zhou et al. [29]
Our study is limited by the geographical area, as we only have isolates from Eastern Denmark. In the future we wish to apply this method to a larger collection of isolates from different countries to learn more about preferred insertion sites for Tn1549 in the E. faecium chromosome. Since we found ten different insertion sites in our collection, we hypothesise that we might find an increased number of insertion sites in isolates from a much larger geographical area. Another limitation is that we did not have the possibility to perform Nanopore sequencing on all 116 non-araA2/non-sir2 isolates, therefore we had to select a subgroup from different MST clusters. The strength of our study is that we include isolates from all DCMs in Eastern Denmark serving approximately half of the Danish population. Our results show that most of the VREfm isolates belong to the two biggest clones, allowing us to perform VREfm diagnostics with our multiplex PCR. However, the present study showed that multiple clusters have the same Tn1549 insertion sites (araA2 or sir2), thus, the multiplex PCR cannot be used as a precise typing tool. The fact that we find the sir2 insertion site in 20 different MST clusters, and the araA2 insertion site in four different clusters, indicate that either the Tn1549 has a few preferred insertion sites in the E. faecium chromosome, or the Tn1549/sir2 has spread to vancomycin-susceptible E. faecium through recombination events.
In our collection of isolates from 2020 to 2022 we found no emerging vanB VREfm clones that would pose a threat to the accuracy of our multiplex PCR. With the results from this study and the identification of eight insertion sites other than araA2 and sir2, we have contributed to the knowledge of possible insertion sites for the vanB Tn1549. We will continue the surveillance of the genomic epidemiology of VREfm, as well as the accuracy of our multiplex PCR. In case of a decrease in the accuracy and if a new dominating clone emerges, we will be able to update our multiplex PCR, to continue to provide rapid VREfm results to our infection control organisations.
Conclusion
In the study period, 94% of vanB isolates had the Tn1549 inserted in either araA2 or sir2 and were identified by our multiplex PCR. We identified eight insertion sites other than araA2 and sir2 and found no emerging clones within these insertion sites. We will continue the surveillance, and update our multiplex PCR, when and if this changes.
Electronic supplementary material
Below is the link to the electronic supplementary material.
Supplementary Material 1
Supplementary Material 2
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Bager P et al (Dec. 2024) Comparison of morbidity and mortality after bloodstream infection with vancomycin-resistant versus -susceptible Enterococcus faecium: a nationwide cohort study in Denmark, 2010–2019. Emerg Microbes Infect 13(1). 10.1080/22221751.2024.230996910.1080/22221751.2024.2309969 PMC 1093011138258968 · doi ↗ · pubmed ↗
- 2Zhou X, Willems RJL, Friedrich AW, Rossen JWA, Bathoorn E (2020) Enterococcus faecium: from Microbiological insights to practical recommendations for infection control and diagnostics. 10.1186/s 13756-020-00770-110.1186/s 13756-020-00770-1PMC 741831732778149 · doi ↗ · pubmed ↗
- 3Contreras GA et al (2022) Contemporary clinical and molecular epidemiology of Vancomycin-Resistant enterococcal bacteraemia: A prospective multicenter cohort study (VENOUS I). Open Forum Infect Dis 9(3). 10.1093/ofid/ofab 61610.1093/ofid/ofab 616PMC 883053035155713 · doi ↗ · pubmed ↗
- 4Fox E et al (2022) Risk factors and outcomes associated with persistent Vancomycin resistant enterococcal bacteraemia. BMC Infect Dis 22(1). 10.1186/s 12879-022-07864-810.1186/s 12879-022-07864-8PMC 967044436384497 · doi ↗ · pubmed ↗
- 5Xavier BB et al (2021) Novel vancomycin resistance gene cluster in Enterococcus faecium ST 1486, Belgium, June Eurosurveillance, vol. 26, no. 36, 2021. 10.2807/1560-7917.ES.2021.26.36.210076710.2807/1560-7917.ES.2021.26.36.2100767 PMC 843199334505571 · doi ↗ · pubmed ↗
- 6Lebreton F, Valentino MD, Schaufler K, Earl AM, Cattoir V, Gilmore MS (2018) Transferable Vancomycin resistance in clade B commensal-type Enterococcus faecium. J Antimicrob Chemother 73(6). 10.1093/jac/dky 03910.1093/jac/dky 039PMC 596131529462403 · doi ↗ · pubmed ↗
- 7Pinholt M et al (2021) Investigation of the introduction and dissemination of van B Enterococcus faecium in the Capital Region of Denmark and development of a rapid and accurate clone-specific van B E. faecium PCR, Journal of Antimicrobial Chemotherapy, vol. 76, no. 9, pp. 2260–2267, Aug. 10.1093/jac/dkab 19810.1093/jac/dkab 19834151364 · doi ↗ · pubmed ↗
- 8Werner G et al (Feb. 2011) Host range of enterococcal Van A plasmids among Gram-positive intestinal bacteria. J Antimicrob Chemother 66(2):273–282. 10.1093/jac/dkq 45510.1093/jac/dkq 45521131318 · doi ↗ · pubmed ↗