Yellow Fever Virus (YFV) Detection in Different Species of Culicids Collected During an Outbreak in Southeastern Brazil, 2016–2019
Giovana Santos Caleiro, Lucila Oliveira Vilela, Karolina Morales Barrio Nuevo, Rosa Maria Tubaki, Regiane Maria Tironi de Menezes, Luis Filipe Mucci, Juliana Telles-de-Deus, Eduardo Sterlino Bergo, Emerson Luiz Lima Araújo, Mariana Sequetin Cunha

TL;DR
This study found yellow fever virus in multiple mosquito species during an outbreak in Brazil, raising concerns about potential urban spread.
Contribution
The detection of YFV in various mosquito species, including non-primary vectors, highlights new transmission risks.
Findings
YFV was detected in 1.4% of 3731 mosquito pools across nine species.
Viral loads varied among species, with Aedes aegypti testing negative.
Infection in non-primary vectors raises concerns about reurbanization of YFV.
Abstract
Yellow fever virus (YFV) is an endemic arbovirus in parts of Africa and the Americas. In Brazil, following the eradication of the urban transmission cycle, YFV is maintained in a sylvatic cycle involving several species of neotropical primates and mosquitoes of the genera Haemagogus and Sabethes, which serve as primary and secondary vectors, respectively. During the 2016–2019 outbreak in São Paulo State, a total of 3731 mosquito pools were collected from sites with ongoing epizootic events in 192 municipalities. The RT-qPCR analysis detected YFV in 46 pools (1.4%) across nine mosquito species, including both primary and secondary vectors, as well as species from the genera Aedes and Psorophora. Differences in viral loads were observed among species. While Aedes aegypti was not found to be positive, the detection of natural YFV infection in other Aedes species raises concerns about…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
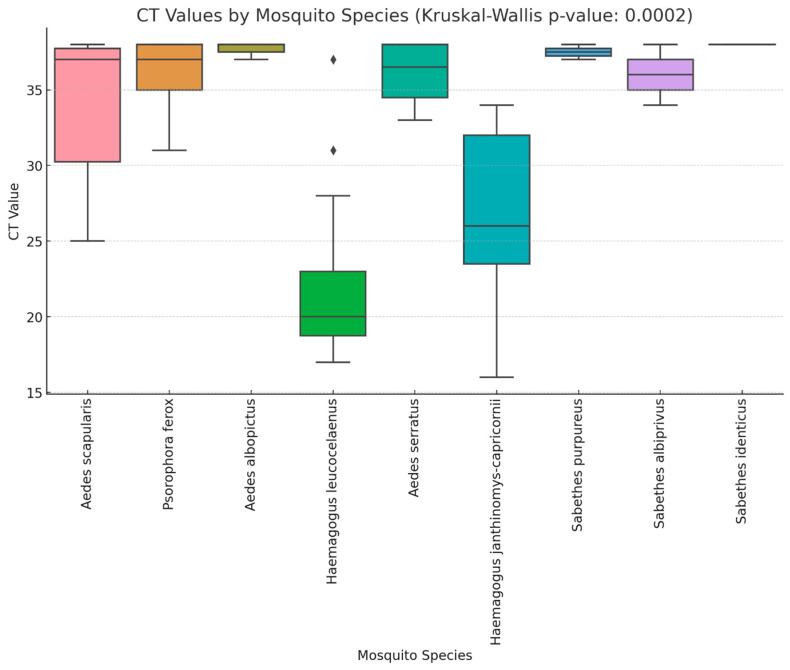 Figure 1
Figure 1- —Secretaria de Estado de Saúde de São Paulo (SES)
- —Fundo Especial de Saúde para Imunização em Massa e Controle de Doenças (Fesima)
- —Fedial (Programa de Formação para Investigação Científica) scholarship from Instituto Adolfo Lutz
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsMosquito-borne diseases and control · Vector-borne infectious diseases · Insect and Pesticide Research
1. Introduction
Yellow fever (YF) (formerly yellow fever virus (YFV) is a severe disease caused by the arbovirus Orthoflavivirus flavi, a member of the Flaviviridae family, and remains a significant public health concern in parts of Africa and the Americas [1]. YF may cause significant morbidity and mortality rates in the human populations, as well as impacting the neotropical primate population [2]. Despite the availability of the live attenuated 17-DD vaccine, a high case fatality rate (CFR) of 40% to 60% persists, particularly in South America [1,3]. In Brazil, after the eradication of the urban YFV cycle in 1942 [4], transmitted by Aedes aegypti mosquitoes, YFV is maintained by a sylvatic transmission cycle involving several species of neotropical primates (NTPs) and forest canopy-dwelling mosquitoes, mainly Haemagogus spp. and Sabethes spp., and human cases are caused by a spillover process in green areas [4]. YF surveillance is based on confirmation of epizootic events through virus detection by RT-qPCR an/or immunohistochemistry in accordance with the Ministry of Health Guidelines [5].
Seasonal climatic variations significantly influence YFV transmission by affecting mosquito population dynamics and viral amplification. During the rainy season, abundant precipitation creates numerous larval habitats, while elevated temperatures and high humidity accelerate mosquito development and viral replication, leading to surges in sylvatic vectors such as Haemagogus and Sabethes spp. These conditions enhance virus amplification among non-human primates and elevate the risk of spillover to humans [6]. Conversely, in the dry season, reduced rainfall limits breeding sites and diminishes vector densities, although desiccation-resistant eggs permit a low-level virus circulation that can rapidly rebound once rains return [7].
From mid-2016 until late 2018, Brazil faced one of the largest YF outbreaks in recent decades, mainly in the southeastern region [8,9,10,11]. São Paulo state, located in southeast Brazil, is the most densely populated state in the country, containing one of the world’s largest urban conurbations [12]. A total of 875 cases of YFV in NTPs between July 2016 and November 2019 and 624 cases of YFV in humans between January 2017 and 18 November 2019 were reported. This outbreak was caused by the 1E lineage belonging to South American I (SA-I) genotype that originated in the Amazon basin [13], which was later disseminated from northern São Paulo into geographically neighboring areas of western MG and into the south of the state [14]. Some epizootic events in Callithrix monkeys were confirmed in large urbanized cities in proximity to urban green areas where Haemagogus and Sabethes mosquitoes were not found [15], indicating that synanthropic mosquitoes were likely involved in viral transmission in these areas. Considering that entomological investigation is a complementary tool to better understand eco-epidemiological aspects of YF after notification of suspected epizootic events, here, we describe different Aedini and Sabethini mosquitoes found to be positive for YFV by RT-qPCR and their ecological factors, showing the continuous threat of the reurbanization of YFV in Brazil.
2. Materials and Methods
2.1. Study Area
This study was conducted in the state of São Paulo, Brazil, which comprises 645 municipalities organized into 15 administrative regions. The state spans approximately 248,196,960 square kilometers and has a population of 44,749,699 inhabitants, primarily concentrated in the coastal region. São Paulo encompasses two distinct biomes, the Cerrado and the Atlantic Forest, both of which have suffered significant deforestation in recent years.
2.2. Epizootic Events and Mosquito Collection
Between November 2016 and June 2019, a total of 3731 mosquito pools from the Aedini tribe and Sabethes genus were formed in 192 municipalities with ongoing epizootic events and adjacent cities. Briefly, dead NTPs were detected and notified by local authorities using the Sistema de Informações de Agravos de Notificação (SINAN), and frozen carcasses were sent to Adolfo Lutz Institute for YFV detection, according to the Brazilian Ministry of Health Guidelines, as previously described [8]. Mosquitoes were then captured at ground level between 9 a.m. and 3 p.m. using entomologic nets and bottle-type manual vacuums in forested and green areas, and Nasci Aspirator in urban dwellings. After sampling, mosquitoes were frozen, transferred to cryogenic tubes, and stored in liquid nitrogen containers for transport. Identification was performed based on morphological characteristics by the Pasteur Institute (formerly the Superintendence for Control of Endemic Diseases—SUCEN). The mosquitoes were subsequently sorted into pools containing 1 to 50 individuals per pool, according to species, collection date, and location. Molecular detection for YFV was carried out on non-engorged mosquitoes (n = 3376) at the local reference laboratory for arthropod-borne viruses at Instituto Adolfo Lutz (IAL) in São Paulo. Pools were triturated in FastPrep-24 5G Instrument (MP Biomedicals, Irvine, CA, USA) and in Magna Lyser (Roche, Basel, Switzerland) in 1 mL of phosphate-buffered saline solution with 0.75% bovine albumin, penicillin (100 units/mL), and streptomycin (100 µg/mL). The resultant suspension was centrifuged at 1800× g for 15 min, and the supernatant was withdrawn and frozen at −70 °C until further use.
2.3. YFV RNA Detection and Statistical Analysis
Viral RNA was extracted using QIAamp Viral RNA Mini Kit following the manufacturer’s instructions (QIAGEN, Hilden, Germany). Detection of YFV RNA was performed using an RT-qPCR protocol [11]. Results with Cycle threshold (Ct) values ≥ 35 were retested. If the new result had a Ct value ≤ 38, the pool was considered positive for YFV. The Kruskal–Wallis test was conducted exclusively among YFV-positive mosquito pools to evaluate differences in viral load by mosquito species, as indicated by their Ct values. To assess differences in yellow fever virus (YFV) viral loads among different mosquito species, a Generalized Linear Model (GLM) was performed with Ct value as the dependent variable. Ct values were used as a proxy for viral load, with lower Ct values indicating higher viral loads.
The primary YFV vector, Hg. leucocelaenus, was set as the reference category to compare viral loads across species. The model was specified as Ct ~ Species, where Ct value was assumed to follow a Gaussian (normal) distribution with an identity link function. The analysis reports estimated mean differences in Ct values (β coefficients) for each species compared to Hg. leucocelaenus. To evaluate whether seasonal variation (rainy vs. dry) influenced Ct values, season was included as an additional predictor in the GLM. An interaction term (Species × Season) was also tested to assess potential species-specific seasonal effects. All p-values < 0.05 were considered significant. All analyses were performed using Rstudio v.2023.12.1, ggplot2 package [16].
3. Results
A total of 3731 mosquito pools were formed during the outbreak (Table 1), of which 46 pools (1.4%) from nine mosquitoes species tested positive for yellow fever virus (YFV), representing 22 municipalities (8.7%) (Table 2). Additionally, epizootic events were confirmed by RT-qPCR in 82 cities (Supplementary Material Figrue S1). The Ct values of the YFV-positive pools ranged from 16 to 38, with a median of 32 (Figure 1).
Among the species collected, Aedes scapularis accounted for 26.46% of all the mosquitoes, with 0.67% of the pools testing positive, followed by Aedes albopictus (21.66%, 0.41% positive) and Psorophora ferox (11.20%, 1.32% positive). Haemagogus leucocelaenus represented 8.09% of the total, with 5.83% of its pools testing positive, while Haemagogus janthinomys/capricornii comprised 3.4%, with 5.51% positive. Other species testing positive for YFV included Aedes serratus (5.72%, 2.07% positive), Sabethes albiprivus (2.67%, 15.78% positive), Sabethes purpureus (0.80%, 2.08% positive), and Sabethes identicus (0.74%, 1.75% positive).
The analysis of the Ct values among the YFV-positive mosquito pools revealed significant differences in the viral loads (Figure 1). The Haemagogus species consistently exhibited the lowest Ct values, indicating higher viral loads, while Sa. albiprivus, Ae. albopictus, Ae. serratus, and Ps. ferox had higher Ct values, suggesting lower viral loads. The distribution of the Ct values varied across species, with some species displaying a wider range, indicating heterogeneity in infection levels within the same species. While the Ae. scapularis pools generally showed high Ct values, two pools recorded Ct values of 25 and 28, suggesting moderate viral loads.
The Generalized Linear Model (GLM) analysis identified significant differences in Ct values among the mosquito species. Hg. leucocelaenus exhibited the lowest Ct values and was used as the reference species. Compared to Hg. leucocelaenus, Hg. janthinomys-capricornii showed a moderate increase in Ct values (β = 4.71, p = 0.039).
The mosquito species from the Aedes, Psorophora, and Sabethes genera exhibited significantly higher Ct values, indicating lower viral loads. Ae. scapularis had a β coefficient of 11.83 (p < 0.001), while Ps. ferox and Ae. albopictus showed β values of 13.80 and 15.67, respectively (p < 0.001). Among the Sabethes species, Sa. purpureus, Sa. albiprivus, and Sa. identicus exhibited the highest Ct values (β = 14.00 to 16.00, p < 0.01).
These results indicate species-specific differences in YFV viral loads, with the Haemagogus species displaying lower Ct values compared to other genera. A full summary of the GLM estimates is presented in Table 3. The effect of season (rainy vs. dry) on the Ct values was not statistically significant (p = 0.173). The interaction between mosquito species and season also did not significantly influence the Ct values (p > 0.3 for all species).
Out of the 46 positive mosquito pools, 24 (52.2%) were collected during the rainy season (18 October–4 April), and 22 (47.8%) during the dry season (Supplementary Material Figrue S2). The Aedes species were predominantly collected during the rainy season, whereas Haemagogus spp. and Psorophora ferox were mostly collected during the dry season. Notably, Sa. albiprivus and Sa. identicus tested positive exclusively in the dry season. All the YFV-positive mosquito pools were collected within the Atlantic Forest biome (Supplementary Material Figrue S3).
4. Discussion
Brazil is an endemic country for YFV, with the Amazon region acting as a source of viral diversity and dispersal across the country. Although YFV circulation has been documented in southeastern Brazil since the early 21st century, the 2016–2018 outbreak caused by the SA-I genotype, particularly in São Paulo state, was unexpected due to the high number of positive cases reported in both humans and animals [14,17]. Notably, during this outbreak, nine different species of Culicidae, including mosquitoes from the Aedes, Psorophora, and Sabethes genera, were found to be positive for YFV by RT-qPCR. All these mosquitoes were collected in the Atlantic Forest biome, where Haemagogus leucocelaenus act as the primary vector [18,19,20]. While Sabethes spp. are traditionally considered secondary vectors [3], limited information is available regarding their role in YFV transmission in this region.
Our findings confirm that the YFV viral loads varied across the Culicidae species, with Hg. janthinomys/capricornii and Hg. leucocelaenus having the highest viral loads. Notably, two pools of Ae. scapularis also had viral loads comparable to those of Hg. janthinomys/capricornii., indicating that this species may play a more relevant role in YFV transmission than previously thought. These pools were collected in Urupês on 15 February 2017, and in Araçatuba on 25 November 2016, at the municipal Zoo, yet neither location reported epizootic events at the time. The presence of YFV in these areas could be attributable to the different susceptibility of NTPs, as some Callithryx sp. may be less susceptible to the disease [2]. Ae. scapularis, which was the most abundant species collected in this study, is considered a generalist in its use of habitats, occurring in both sylvatic and human-dominated areas. Adult females are opportunistic in their behavior, feeding especially on mammals [21,22]. Considering the wide host breadth and feeding habitats, coupled with synanthropic adaptions, it is possible that Ae. scapularis may be an important bridge vector for human and animal viruses. Thus, our data suggest that this species may have played a secondary role in the YF outbreak.
Sabethes mosquitoes were observed to have low abundance, distribution, and infection rates, suggesting a local or secondary role during the 2016–2018 outbreak in the Brazilian southeastern region [18]. In our study, this genus accounted for 11.5% (n = 430 pools) of the collected mosquitoes and for 10.9% (n = 5/46) of the positive pools, all exhibiting high Ct values, indicative of low viral loads. Similarly, during the 2009 YF outbreak in São Paulo, YFV was only isolated from a single pool of Hg. leucocelaenus in Buri, despite the collection of Sa. chloropterus, Sa. purpureus, and Sa. undosus in the same area [19]. However, the absence of an RT-qPCR analysis in that study may explain the lack of positive detections among Sabethini mosquitoes. Conversely, during a YF epidemic and epizootic in Misiones, a northeastern province of Argentina, YFV was successfully isolated in a cell culture from pools of Sabethes albiprivus [23]. This viral isolation indicates high viral loads, contrasting with the low viral loads observed in the Sabethes specimens from the Atlantic Forest.
Sa. chloropterus has been identified as the primary YF vector during the dry season in the Cerrado biome of Minas Gerais [24]. In Espírito Santo, where the sylvatic YF cycle was first described in Brazil, Sa. chloropterus, Sa. soperi, Sa. identicus, Aedes aureolineatus, and Shannoniana fluviatilis were noted for their secondary roles in YFV transmission [25]. Additionally, Sa. albiprivus from Rio de Janeiro demonstrated high vector competence when inoculated with Brazilian YFV strains [26]. To better elucidate the role of Sabethes mosquitoes in the YF transmission cycle within São Paulo state, where the virus has now been established [27], additional studies are required.
Regarding the Aedes genus, earlier studies suggested that Brazilian Ae. aegypti mosquitoes might not favor the establishment of an urban cycle of YF [28]. However, a more recent study demonstrated that both anthropophilic mosquitoes, Ae. aegypti and Ae. albopictus, are highly susceptible to American and African YFV strains [26]. In 2018, in Minas Gerais state, a single Ae. albopictus mosquito pool tested positive for YFV [29]. In our surveillance study, Ae. albopictus was the second most frequent species collected, accounting for 21.66% of the total, with three pools testing positive for YFV, all of which exhibited low viral loads. No Ae. aegypti mosquitoes were found to be positive. Despite the high number of human infections during the outbreak, no urban YF cases were reported. Given that YFV has demonstrated potential for adaptation to Ae. albopictus and can be transmitted between NTPs [30,31] our findings underscore a potential threat to the endemic areas in South America where these mosquitoes are present. With its widespread distribution and ecological plasticity, Ae. albopictus could serve as a bridge vector, facilitating virus transmission between urban environments and rural areas.
One objective of this study was to assess whether seasonal variation (rainy vs. dry) influenced YFV viral loads in mosquitoes. Despite previous reports showing seasonal peaks in mosquito abundance and transmission during rainy periods [32] our results indicate that season was not a significant predictor of Ct values, suggesting that once a mosquito is infected, viral replication remains stable. Sacchetto and collaborators reported viral persistence during the non-epidemic dry season in NTPs collected in Belo Horizonte, Minas Gerais state [33]. These results show the importance of continuous surveillance, regardless of seasonal variations.
Our study has some limitations. Specifically, our study involved triturating whole mosquitoes instead of processing solely the salivary gland. Additionally, the contents of the mosquitoes’ digestive systems—whether engorged or not—were assessed solely through visual examination, and some of the positive results could have come from residual blood feeding. Nevertheless, the data obtained in the present study are relevant, as monitoring of virus circulation and characterizing vectors are fundamental elements for understanding the dynamics of vector-borne viruses, providing new insights for the establishment of control strategies and to prevent the risk of the re-urbanization of YFV. More studies of vectorial competence, mainly in Ae. scapularis, are needed, as our results suggest a possible role of Ae. scapularis in the YFV cycle in the Atlantic Forest.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Vasconcelos P.F.C. Monath T.P. Yellow Fever Remains a Potential Threat to Public Health Vector Borne Zoonotic Dis.20161656656710.1089/vbz.2016.203127400066 · doi ↗ · pubmed ↗
- 2de Azevedo Fernandes N.C.C. Guerra J.M. Díaz-Delgado J. Cunha M.S. Saad L.D. Iglezias S.D. Ressio R.A. Cirqueira C.d.S. Kanamura C.T. Jesus I.P. Differential Yellow Fever Susceptibility in New World Nonhuman Primates, Comparison with Humans, and Implications for Surveillance Emerg. Infect. Dis.202127475610.3201/eid 2701.19122033350931 PMC 7774563 · doi ↗ · pubmed ↗
- 3Monath T.P. Vasconcelos P.F.C. Yellow fever J. Clin. Virol.20156416017310.1016/j.jcv.2014.08.03025453327 · doi ↗ · pubmed ↗
- 4Vasconcelos P.F. Costa Z.G. Travassos D.R.E.S. Luna E. Rodrigues S.G. Barros V.L. Dias J.P. Monteiro H.A. Oliva O.F. Vasconcelos H.B. Epidemic of jungle yellow fever in Brazil, 2000: Implications of climatic alterations in disease spread J. Med. Virol.20016559860410.1002/jmv.207811596099 · doi ↗ · pubmed ↗
- 5Ministerio da Saúde Brasília-Df 2017 Guia de Vigilância de Epizootias em Primatas não Humanos e Entomologia Aplicada à Vigilância da Febre Amarela Ministério da Saúde 2 a Edição Atualizada 2017 Available online: https://www.gov.br/saude/pt-br(accessed on 11 December 2024)
- 6Li S.L. Acosta A.L. Hill S.C. Brady O.J. de Almeida M.A.B. Cardoso J.d.C. Hamlet A. Mucci L.F. de Deus J.T. Iani F.C.M. Mapping environmental suitability of Haemagogus and Sabethes spp. mosquitoes to understand sylvatic transmission risk of yellow fever virus in Brazil P Lo S Neglected Trop. Dis.202216 e 001001910.1371/journal.pntd.001001934995277 PMC 8797211 · doi ↗ · pubmed ↗
- 7Silva-Inacio C.L. Ximenes M.d.F.F.d.M. Haemagogus spegazzinii Brèthes, 1912 (Diptera: Culicidae) in Brazilian semiarid: Resistance in eggs and scale color variation in adults Rev. Bras. Entomol.202165 Epub ahead of print 10.1590/1806-9665-rbent-2021-0078 · doi ↗
- 8Cunha M.S. da Costa A.C. de Azevedo Fernandes N.C.C. Santos F.C.P.D. Nogueira J.S. D’Agostino L.G. Komninakis S.V. Witkin S.S. Ressio R.A. Maeda A.Y. Epizootics due to Yellow Fever Virus in São Paulo State, Brazil: Viral dissemination to new areas (2016–2017)Sci. Rep.20199547410.1038/s 41598-019-41950-330940867 PMC 6445104 · doi ↗ · pubmed ↗