Sindbis Virus Platform Provides an Oncolytic-Virus-Mediated and Immunotherapeutic Strategy to Overcome the Challenging Microenvironment of Pancreatic Cancer
Silvana Opp, Christine Pampeno, Alicia Hurtado, Daniel Meruelo

TL;DR
A Sindbis virus platform shows promise in treating pancreatic cancer by reducing tumors and boosting immune responses in mouse models.
Contribution
The study demonstrates a novel oncolytic virus strategy combining IL12 expression and anti-OX40 to target pancreatic cancer's challenging microenvironment.
Findings
SV vectors reduced pancreatic tumors in three mouse models, including one with a highly immunosuppressive environment.
Combining SV with anti-OX40 increased immune cell infiltration into the tumor microenvironment.
The treatment prevented tumor recurrence in mice rechallenged with cancer cells.
Abstract
Background/Objectives: Our laboratory has been developing a Sindbis viral (SV) vector platform for treatments of several types of cancers. In this study, we assess treatment efficacy for metastatic and immunosuppressive pancreatic cancer. Methods: Orthotopic mouse models were generated by injection of tumor cells into the pancreatic parenchyma. Sindbis vectors were inoculated intraperitoneally. Imaging of tumors was performed by either MRI or in vivo imaging using luciferase. Flow cytometry, multi-immunofluorescence and elispot analysis were performed for certain tumors. Results: SV can infect and reduce pancreatic tumors in three mouse model systems: a model bearing human pancreatic tumors, a highly metastatic model, and a model that reflects the highly immunosuppressive, desmoplastic microenvironment common to human pancreatic cancer. Conclusions: Combination of SV vector expressing…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
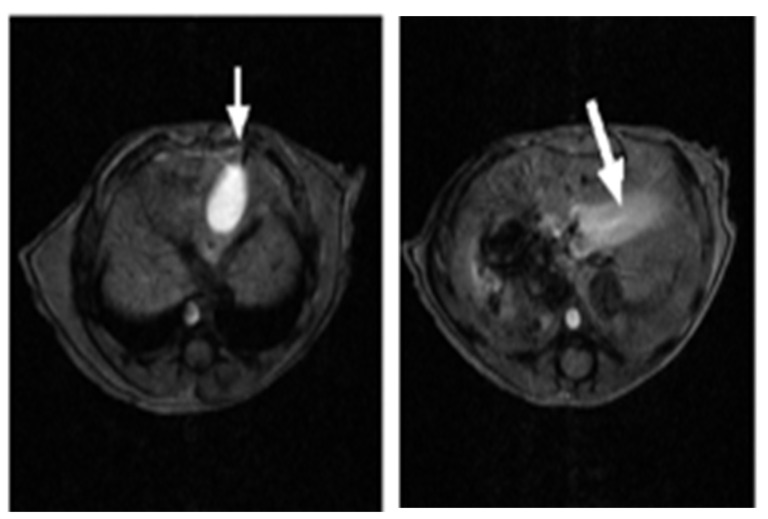 Figure 1
Figure 1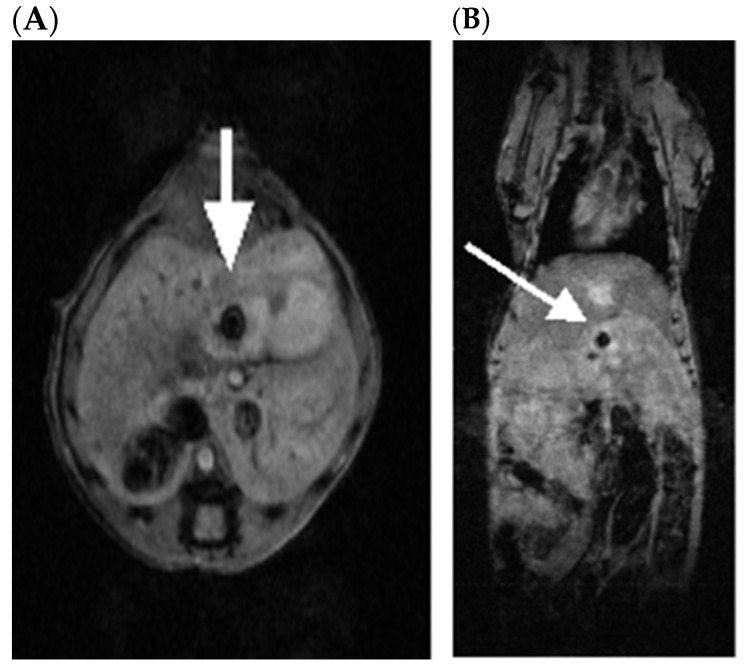 Figure 2
Figure 2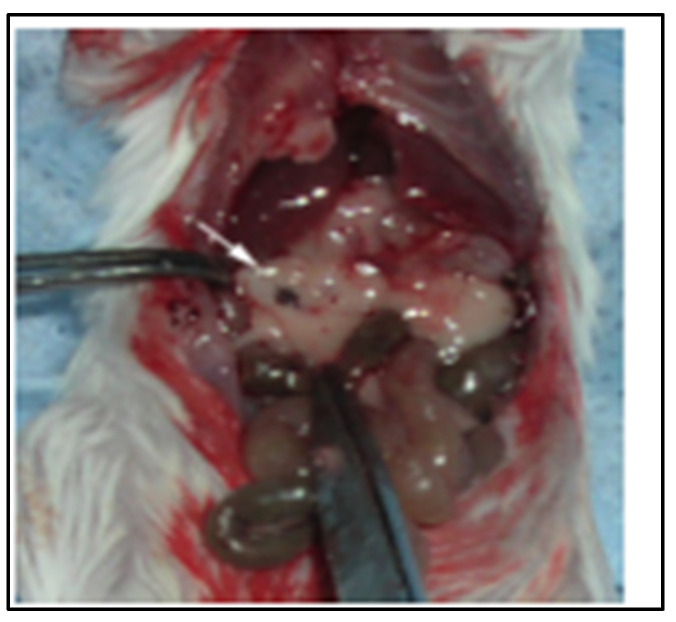 Figure 3
Figure 3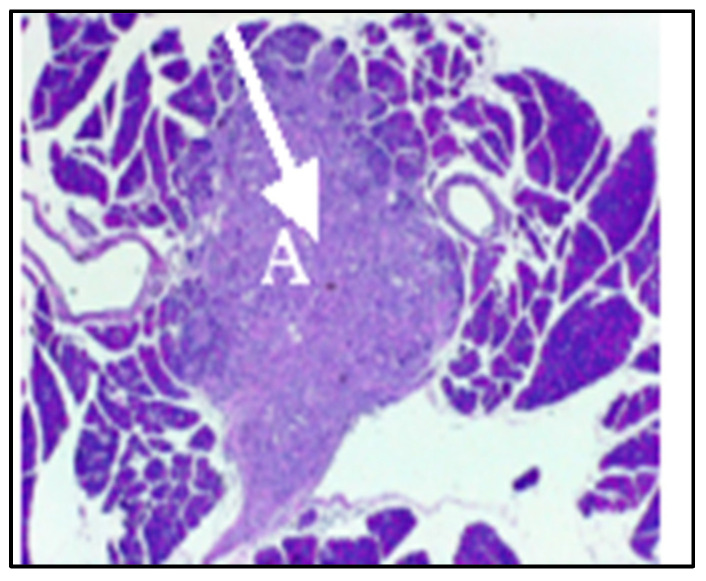 Figure 4
Figure 4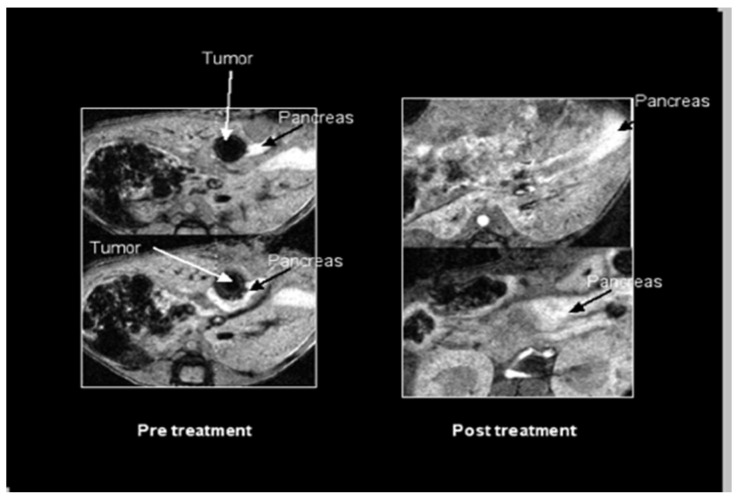 Figure 5
Figure 5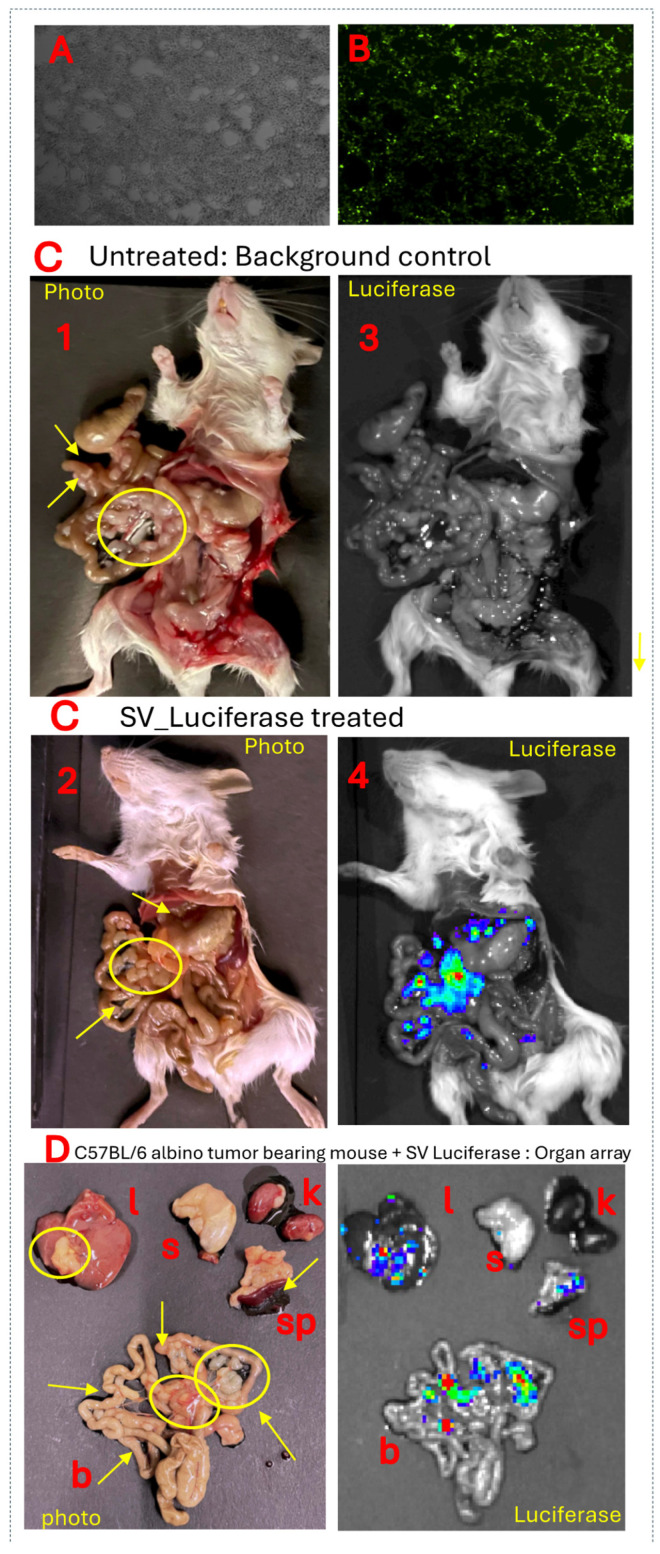 Figure 6
Figure 6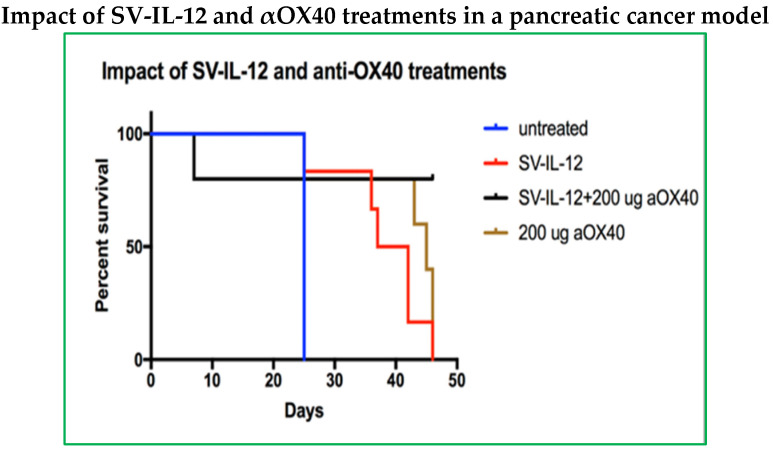 Figure 7
Figure 7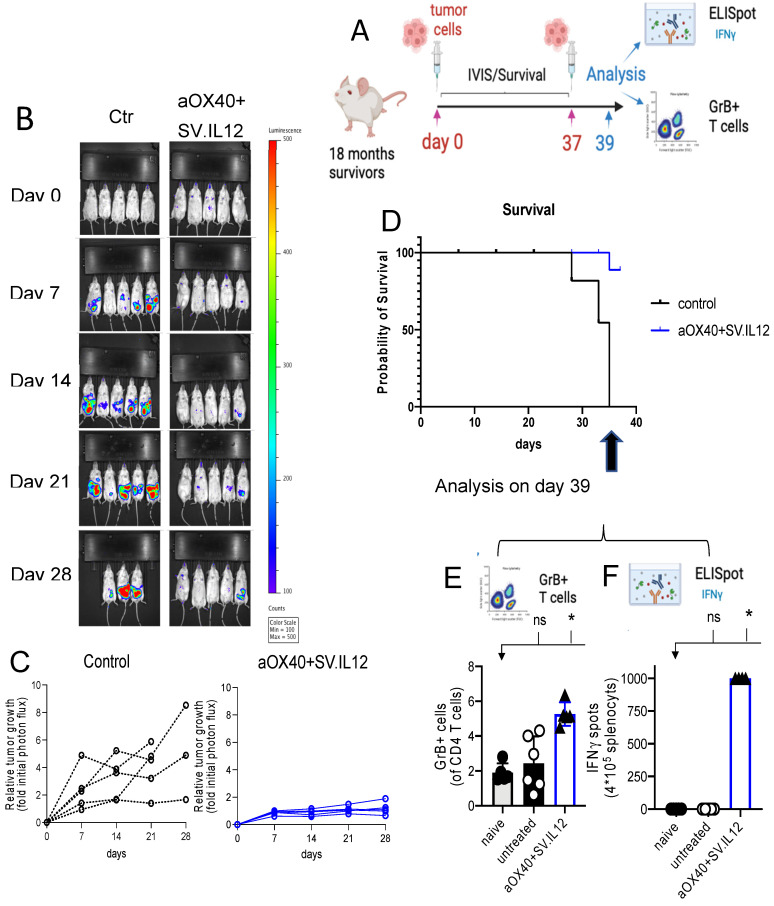 Figure 8
Figure 8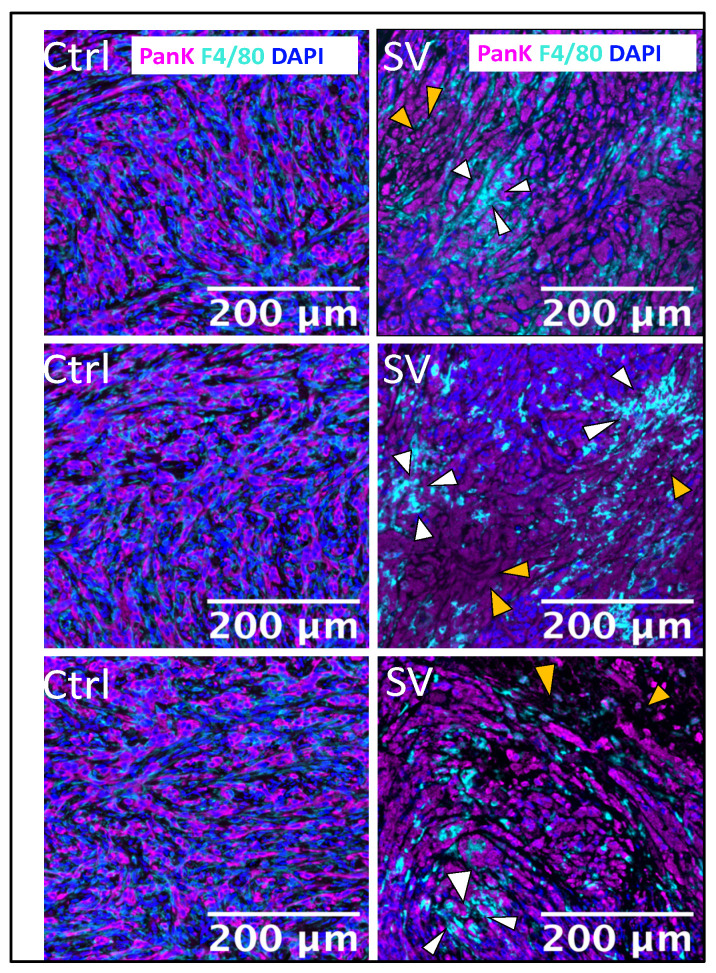 Figure 9
Figure 9- —NIH
- —The Experimental Pathology Research Laboratory at NYU Langone that is partially supported by the Cancer Center Support
- —Shared Instrument
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsVirus-based gene therapy research · Cancer Research and Treatments · CAR-T cell therapy research
1. Introduction
A Sindbis viral (SV) vector expressing an immunomodulatory molecule, agonistic OX40 antibody (αOX40), combined with the cytokine IL-12, has shown strong efficacy in the treatment of highly malignant ovarian cancer preclinical models [1,2]. The success of these studies prompted evaluation of this vector for the treatment of pancreatic cancer, another lethal malignancy with poor prognosis.
Pancreatic cancer is typically diagnosed at a late stage when symptoms become apparent, and metastasis most likely occurs. Pancreatic ductal adenocarcinoma (PDAC), the most common type of pancreatic cancer (95%), has a five-year survival rate of only 13% [3]. Despite progress using chemotherapeutic agents, such as FOLFIRINOX and gemcitabine plus albumin-bound paclitaxel [4], <30% of patients respond to either regimen, and median progression-free and overall survival remain under 6 and 12 months, respectively [5].
A major challenge in PDAC treatment involves an immunosuppressive tumor microenvironment (TME) and a dense extracellular matrix known as desmoplasia [6,7,8,9]. The desmoplastic TME elevates interstitial fluid pressure that limits small-molecule perfusion and contributes to the restriction of nutrients and oxygen due to metabolic deregulation of tumor cells. Infiltrating T cells, requiring high energy demands, become dysfunctional in this environment [10,11].
Our studies involving multiple mouse tumor models indicate that combining an SV vector with agonistic antibodies to OX40 (αOX40) can confer complete remission and protection from recurrence by driving activated T cells selectively into the TME [1,12,13]. OX40 (CD134), a member of the tumor necrosis factor (TNFR) superfamily, is a co-stimulatory receptor that is expressed on activated T cells. Interaction between OX40 and its ligand, OX40L, promotes the clonal expansion, differentiation, and survival of CD4 Th1 helper cells, which produce IFNγ and IL-2 cytokines [14,15,16,17] that sustain the survival of primed CD8 T cells [18,19]. The co-expression of OX40 with ICOS on follicular T helper cells (Tfh) facilitates the differentiation of antibody-producing B cells and long-lived plasma cells from germinal center B cells [20]. In addition, OX40 signaling represses regulatory T cells (Treg) by downregulating the expression of Foxp3 [21].
The observation that IL-12 increases the presence of OX40 on the surface of CD4 T cells initiated the study of a combined anti-tumor capacity [13,22]. IL-12 activates T cells, stimulates the production of IFNγ and increases the expression of OX40 on effector CD4 T cells [22]. The combination of SV.IL-12 with an agonistic antibody to OX40 has been shown to exhibit strong therapeutic efficacy in CT26, colon, and MyC-CaP, prostate carcinoma, models [13]. SV expressing IL-12 combined with agonistic OX40 antibody changes the transcriptome and metabolic program of T cells, developing highly activated, terminally differentiated effector T cells, with enhanced tumor infiltration capacity that can overcome the repressive TME [1,12,13]. Intra-tumoral T cell immune responses showed increased granzyme B levels. Anti-tumor activity was observed by a decrease in Ki-67 proliferation marker in tumor cells. Early studies had shown that SV could significantly reduce subcutaneous tumors in a C.B-17-SCID mouse model bearing human pancreatic CFPAC cells [23] and in a syngeneic Pan02 C57BL/6 model [24]. The Pan02 cell line was also used to show that protein kinase R senses SV infection activating translational arrest, cellular stress, and apoptosis [25].
We have developed a Sindbis viral (SV) vector platform for the treatment of multiple types of tumors [1,12,13,24,26,27]. Sindbis is a positive single-strand RNA oncolytic alphavirus that can infect and specifically replicate in tumor cells, eliciting cell death and stimulating anti-tumor immune responses. The utility of alphaviruses as vector systems and oncolytic-virus-mediated therapy has been reviewed [28,29,30]. Reviews of their structure, expression, replication, and evolution have also been presented [31,32,33].
In this study, we demonstrate that SV vectors can effectively treat PDAC in three different preclinical model systems: human pancreatic cancer, a metastatic cancer model, and a model with an immunosuppressive TME.
2. Results
2.1. SV Vectors Can Generate Regression of Human Pancreatic Tumors in an SCID Mouse Model
To assess the ability of SV to infect and treat human pancreatic tumors, 2.5 × 10^6^ CFPAC-1 cells were injected directly into the pancreatic parenchyma of SCID mice following a modified protocol from Tomioka et al. [34]. Magnetic resonance imaging (MRI) was used to facilitate monitoring of tumor growth and therapy in these mice. At 21 days post-surgical implantation, animals were examined 30 minutes after IV injection of an MRI contrast medium, MnDPDP [manganese (II) N,N′-dipyridoxylethylene-diamine-N,N′-diacetate-5,5′-bis (phosphate)] (0.5 µmol/kg). Figure 1 shows that in an untreated control animal, contrast accumulated in the gallbladder as anticipated (right) and the pancreas was visualized as a bright band of signal in the expected location in the upper abdomen (left).
In tumor-bearing mice, each tumor appeared as a dark focus in the pancreatic neck (Figure 2). Following the initial imaging experiment, one animal was sacrificed and the pancreas was inspected for gross tumors and evaluated by histopathology with hematoxylin and eosin staining. On gross inspection, a nodular mass could be recognized in this location (Figure 3). Histologic evaluation confirmed this to be adenocarcinoma (Figure 4). Having established the model, we tested a second set of animals to determine whether SV vectors could prove therapeutic. As before, the pancreas was surgically exposed and CFPAC-1 cells were injected into each of five SCID mice. Animals were imaged 21 days after surgical implantation. Tumors were detected in all the injected mice. Following imaging, mice were treated daily for 2 weeks with SV vectors (~10^8^ TU per day/5 days per week) to determine whether tumor regression could be induced in the pancreas and whether such regression was detectable on re-imaging. In this model, treatment with SV vectors caused complete tumor regression of orthotopically implanted human pancreatic cancer cells growing in the pancreas of SCID mice (Figure 5).
2.2. SV.IL-12 Can Reduce Metastasis in a Pan02 Syngeneic Mouse Model
To address the impact of our therapy on metastasis, we used the Pan02 cell line derived from a pancreatic tumor that developed in a C57BL/6 mouse following implantation of 3-methylcholanthrene-soaked cotton threads into pancreas tissue [35]. Pan02 tumors have intrinsically high resistance to a wide range of chemotherapeutic agents and significant metastatic burden [35,36]. At 5 weeks post-Pan02 tumor implantation, when metastases were observed to occur mice were treated daily, five times per week, for 4 weeks with 10^8^ TU of SV vector expressing IL-12 (SV.IL-12). A comparison of the anti-tumor response seen against Pan02 metastases is shown on Table 1. SV.IL-12 was found to significantly reduce metastatic growth; however, complete remission was not observed. Previously, it has been shown that the IL-12 cytokine alone, i.e., without the vector, offers very little if any protection [13].
2.3. Effect of SV Vector on PDAC Models with Immunosuppressive TME
A limitation of the Pan02 model is that the orthotopic implants do not develop as pronounced a desmoplastic reaction as seen in human PDAC tumors [37,38]. KC and KPC models are more analogous to human PDACs. Histological analyses of implanted KC and KPC cell lines demonstrate similar levels of fibrosis and leukocyte infiltration to those that arise in genetically engineered mouse models of spontaneous PDAC but express far fewer neoantigens [37,38].
UN-KC-6141 cells were derived from a pancreatic tumor of a KrasG12D;Pdx1-Cre (KC) mouse engineered to express gene mutations commonly observed in pancreatic cancer patients. These mice spontaneously produce tumors with features of human PDAC [39]. Figure 6 shows that SV expressing firefly luciferase (SV.Luc) was observed by IVIS imaging to specifically infect C57BL/6 albino orthotopic UN-KC-6141 tumors. This model, therefore, provides a means to test SV combination immunotherapy for the treatment of the immunosuppressive TME and desmoplasia characterizing PDAC. The results indicate that the combination of SV with IL-12 and αOX40 increases the therapeutic potential (Figure 7). In this experiment, UN-KC-6141 cells expressing firefly luciferase were used for IVIS imaging to monitor tumor growth. Four days after tumor inoculation, groups of mice (n = 25) were treated for one month with the indicated SVs (10^6^ TU, 4 days/week) and OX40 antibody (200 µg), as indicated, 3 days per week. Control mice (n = 10) were untreated.
Figure 8 demonstrates that SV.IL-12 vectors with αOX40 therapy confer long-lasting protection. A schematic of the experiment is shown in Figure 8A. UN-KC-6141-Luc tumor-bearing mice, surviving 18 months after treatment with SV.IL12 + αOX40, were re-challenged with 5 × 10^5^ UN-KC-6141-Luc tumor cells. Age-matched naïve mice received similar inoculation of tumor cells. Figure 8B,C show that by both photon flux and relative growth, animals previously cured by treatment with the SV.IL12 and αOX40 therapy are more protected than age-matched naïve mice. Survival of re-challenged mice was seen in 80% of those previously cured compared with naïve untreated mice (Figure 8D). To analyze immune-cell-mediated anti-tumor responses associated with long-term protection in the same re-challenged survivors, we reinjected them with UN-KC-6141 tumor cells on day 37, and then excised spleens and stained a single cell suspension for flow cytometry analysis on day 39. Naïve (mice not previously challenged with tumor or treated) and untreated mice (mice challenged with tumor cells on day 37) were used as controls. The results showed that the re-activation of granzyme B-expressing T cells (Figure 8E) and upregulation of IFNγ secretion (Figure 8F) helped to prevent tumor regrowth. No treatment with SV.IL12 + αOX40 was administered in this experiment. The observed protection results from memory T cell responses. Mechanistically, SV.IL-12 + αOX40 therapy achieves regression-free survival by driving activated T cells selectively and measurably into the cold, immunosuppressed tumor [12]. Contributing to this effect is that the combination of systemically administered SV vectors and αOX40 markedly changes the transcriptome signature and metabolic program of T cells, driving the development of highly activated, terminally differentiated, effector T cells. These reprogrammed T cells demonstrate enhanced tumor infiltration capacity, as well as anti-tumor activity throughout the body, overcoming the repressive TME [13,40].
SV tumor targeting and apoptosis can be similarly observed in the UN-KC-6141-pancreatic cancer model by use of multicolor immunohistochemistry. Staining with Pan cytokeratin (PanK) shows the presence of tumor cells that are killed by treatment with SV. Macrophage migration into the tumor TME induced by SV is revealed by staining with F4/80 antibody (Figure 9).
3. Discussion
Pancreatic cancer treatment requires targeting of primary and metastatic tumor cells as well as the desmoplasmic TME. Combinatorial treatment strategies that target both the TME and tumor cells of PDAC have been studied and multiple clinical trials are underway [9]. These include combining immune checkpoint blockade antibodies with drugs that target stromal elements or immunosuppressive myeloid cell inhibitors. Like OX40, another co-stimulatory TNFR, CD40, has been combined with chemotherapy. CD40 activates antigen-presenting cells (APCs) that can elicit an immune response to tumor antigens. When combined with chemotherapy, CD40 antibody agonists activate APCs to destroy tumor stroma and drive anti-tumor T cell responses in mouse models [41]. CD40 immunotherapy, in combination with gemcitabine and nab-paclitaxel chemotherapy, is currently in phase II clinical trials (ClinicalTrials.gov, NCT03214250) [42].
Oncolytic viruses (OVs) have been recognized as an immunotherapy that can overcome challenges presented by the TME. By selectively infecting and lysing tumor cells, OVs release tumor-associated antigens and danger-associated molecular patterns that trigger IFN type I responses, promoting antigen-presenting cell maturation that activates T cells. Preclinical and ongoing clinical trials have been described that employ OVs as monotherapy, with some expressing transgenes that target the TME or enhance the anti-tumor immune response. Combinations of OVs with immune checkpoint blockades and/or chemotherapy are also under investigation (reviewed in [43,44]).
Investigations related to our SV.IL12, αOX40 therapy have been performed. A long-term follow-up phase I clinical trial (2017–2024) of an adenovirus expressing IL12 has shown promising results for the treatment of metastatic cancer (ClinicalTrials.gov, NCT02555397) [45]. The adenovirus vector, inoculated intra-tumorally, was combined with chemotherapy in this study. Patients showed increased overall survival and elevated serum levels of immunostimulatory cytokines IL12 and IFNγ, and the chemokine CXCL10. The results of this study have encouraged upcoming phase II/III clinical trials.
Two TNFR family superfamily member co-stimulatory ligands, CD40L and 4-1BBL, have been encoded within an adenovirus, LOAd703, for intra-tumoral injection into PDAC tumors. LOAd703 treatment, combined with chemotherapy, was evaluated in a phase I clinical trial (ClinicalTrials.gov, NCT02705196) [46]. The results showed enhanced immune response and the study will continue with the addition of anti-PD1 inhibitor, atezolizumab.
In a preclinical study, OX40 ligand (OX40L) was inserted into the herpes simplex OV (HSV-1) and treatment of PDAC was evaluated in the KPC syngeneic mouse model [47]. The results support our data showing that OX40 stimulation suppresses tumor growth and prolongs survival (Figure 8). HSV-1 OV-mOX40L activated CD4 and CD8 T cells, as evidenced by increased IFNγ and granzyme b production, and increased migration of immune cells into the TME, similar to our observations with the UN-KC-6141 tumor model (Figure 9). In another study, the addition of αOX40 to an anti-PD1 checkpoint blockade was also shown to greatly augment the efficacy of treatment in the KPC PDAC model in the absence of OV therapy [48].
This report demonstrates that our SV platform promises to be an effective treatment for PDAC. SV was shown to reduce CFAC-1 human tumors in a xenotrophic SCID mouse model.
SV.IL12 also targeted and reduced tumors in a metastatic Pan02 syngeneic mouse model. Most notably, SV.IL12 combined with αOX40 was shown to infiltrate the desmoplasia of UN-KC-6141 pancreatic tumors resulting in tumor lysis, long-term protection against recurrence, and increased infiltration of macrophage cells.
The SV platform affords certain advantages over other oncolytic viruses: (1) Alphaviruses, like SV, are known to target lymph nodes and infect monocytes, macrophages, and dendritic cells, thus promoting the activation and priming of T cells [1,49,50,51,52]. Consequently, SV vectors can take advantage of the existing T cell repertoire, obviating prior knowledge of tumor-associated antigens (TAAs). (2) Sindbis virus is transmitted via mosquito bites to mammals [32]. As a blood-borne vector, SV can be administered via intravenous (i.v.) and intraperitoneal (i.p.) routes. The hematogenous delivery property of Sindbis virus enables it to reach tumor cells throughout the circulation, avoiding more complicated intra-tumoral injections [24]. (3) Studies show that repeated administration of vectors remains efficacious [53,54,55]. (4) SV has the safest profile among alphaviruses, with mostly asymptomatic infections in the wild [56,57]. SV is an RNA virus without replicative DNA intermediates that pose any risk of chromosomal integration or insertional mutagenesis [58]. To avoid even transient adverse effects, our vectors have been attenuated by splitting the SV genome and by removing the packaging signal from the genomic strand that encodes the structural genes [59,60].
In summary, we have shown in colon, prostate, and ovarian cancer models [1,13], and here in a pancreatic cancer model, that SV expressing IL12 in combination with stimulatory αOX40 can effectively reduce tumor burden, provide long-term protection, and stimulate anti-tumor immune responses. An SV has been generated that expresses both IL12 and αOX40.
4. Materials and Methods
4.1. Cell Lines
Human CFPAC-1 PDAC were obtained from the American Type Culture Collection (Manassas, VA, USA) (ATCC, CRL-1918); and cultured in DMEM (low-glucose modified, JRH Biosciences; Lenexa, KA, USA) supplemented with 10% FBS. Pan02 cells were obtained from the NCI–Frederick Cancer Research Facility (Frederich, MD, USA) and were maintained in DMEM supplemented with 10% FBS. The UN-KC-6141 cell line, a gift from the Surinder K. Batra laboratory (Univ. of Nebraska Medical Center, Omaha Nebraska, NE, USA), was cultured in DMEM medium supplemented with 10% FBS.
For IVIS imaging, a firefly luciferase (Luc)-expressing UN-KC-6141 cell line (UN-KC-6141-Luc) was generated by stable transfection of the pGL4.20_Fluc plasmid (Promega, Madison, WI, USA). All media were supplemented with 100 μg/mL of penicillin–streptomycin and 0.5 μg/mL of amphotericin B (Mediatech, Inc., Manassas, VA, USA). Cells were incubated at 37 °C, 5%CO_2_.
4.2. SV Production
The SV vectors used in these studies are propagation-defective and were produced as previously described [13,23,61]. Briefly, plasmids carrying the replicon or Sindbis helper RNAs were linearized with PacI, NotI, or Xhol, and in vitro transcribed using mMessage mMachine T7 or SP6 kits (InVitrogen, Thermo Fisher, Fairlawn, NJ, USA) according to protocol. RNA quality was assessed by gel electrophoresis and 18 µL of helper and replicon reactions was co-electroporated (25 µF, 1.2 kV) into 6 × 10^6^ BHK-21 cells. Electroporated cells were incubated in 10 cm dishes at 37 °C overnight. Cells were washed with PBS and 8 mL of Opti-MEM (minus FBS) (Thermo Fisher, Fairlawn, NJ, USA) supplemented with 100 mg/L CaCl_2_. After 24 h, supernatant was separated by centrifugation at 3000 rpm, for 15 min, and aliquoted for storage at −80 °C. Vectors were titrated in BHK cells, expressed as transducing units (TU) per ml. In SV.IL12, the IL-12 sequence is a synthetic construct of bioactive single-chain murine interleukin 12 mRNA [Genbank Sequence ID: AP411293.l]. SV.Luc expresses firefly luciferase excised from the pGL3 plasmid (Promega, Madison, WI, USA) [61].
4.3. Animal Experiments and Tumor Models
All experiments were in accordance with the Institutional Animal Care and Use Committee of New York University Langone Health [Approval Record: IA17-00796, 7/31/23]. Mice were used at 8 weeks of age. For imaging experiments, isoflurane was used as an anastatic. The CFPAC-1 human PDAC model was established by injection of 2.5 × 10^6^ cells into the pancreatic parenchyma of SCID mice (Jackson Labs, Bar Harbor, ME, USA) by a modified protocol from Tomioka et al. [34]. Groups of 5 mice were used for control and experimental treatments. Imaging was performed at the Animal Facility and the Preclinical Imaging Laboratory at NYU Langone Health as described in the text. The metastatic Pan02 model was established by orthotopic injection of 2.5 × 10^6^ cells into the pancreatic parenchyma of C57 BL/6 mice (Jackson Labs, Bar Harbor, ME, USA). Treatment with SV.IL12 (10^8^ TU) began 5 weeks after tumor cell inoculation; i.p., 5 times per week for 4 weeks. Groups of 5 mice were used for control and experimental treatments. The KPC orthotopic model was established by intra-pancreal injection of 5 × 10^5^ UN-KC-6141 cells into C57BL/6 albino mice, B6 (Cg)-Tyr-21/J, (Jackson Labs, Bar Harbor, ME, USA) to facilitate noninvasive bioluminescent imaging using the IVIS (in vivo imaging system) Spectrum Imaging System (Caliper Life Sciences, Hopkinton, MA, USA). Mice were randomized into untreated and treated groups for survival experiments. Tumor growth was quantified using the Living Image 3.0 software (Caliper Life Sciences, Hopkinton, MA, USA) as previously described [27]. Briefly, mice were injected with 150 mg/kg D-luciferin firefly and potassium salt (GoldBio, St. Louis, MO, USA), and luminescence was assessed 5 min later. Relative tumor growth was calculated individually for each animal by dividing the total body bioluminescence signal of a given day by the first one registered before treatment. As per the indicated treatment strategy, mice were injected i.p. with indicated doses of SV vectors and/or a commercial αOX40 antibody (dose—250 μg/mouse, BioXcell, Lebanon, NH, USA BE0031).
4.4. Histochemistry and Multiplex Immunofluorescence (MIF)
Tumors of mice were collected, fixed in 4% paraformaldehyde (PFA) for 2 days, embedded in paraffin, sectioned, and hematoxylin and eosin-stained. For MIF staining and imaging, 5 μm paraffin sections were stained with an Akoya Biosciences Opal multiplex automation kit (Akoya Biosci., Marlborough, MA, USA; 01752) on a Leica BondRX autostainer (Leica Biosystems, Lincolnshire, IL, USA), according to the manufacturer’s instructions as previously described [1]. Pancytokeratin (C-11) and macrophage F4/80 (EPR26545-166) were obtained from Abcam, Cambridge, UK.
4.5. Flow Cytometry Analysis of Granzyme B on CD4 T Cells and Elispot Analysis of IFNγ
For flow cytometry and elispot analysis, spleens were mashed though a 70 µm strainer before red blood cells were lysed with ammonium–chloride–potassium (ACK, Gibco, Thermo Fisher, Fairlawn, NJ, USA). Cells were washed with PBS with 1% FBS and surface receptors were stained as previously described [12,13]. Fluorescence-conjugated antibodies for CD4 and granzyme B were purchased from BioLegend (San Diego, CA, USA). Mouse IFNg ELISPOT was performed according to the manufacturer’s protocol (BD Biosciences, Franklin Lakes, NJ, USA). T cells (8 × 10^4^) were plated per well overnight in RPMI 1640 supplemented with 10% FCS. No additional stimulus was used in the elispot. As a positive control, the cells were stimulated with 5 ng/mL phorbol myristate acetate (PMA) + 1 mg/mL ionomycin (Mediatech, Inc., Manassas, VA, USA).
4.6. Statistical Analysis
Analysis was performed using GraphPad Prism 9.0. Treated groups were compared, with a one-way analysis, to naive mice. Differences with a p value of <0.05 were considered significant as determined with the Kruskal–Wallis test followed by the Dunn or Mann–Whitney test.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Opp S. Hurtado A. Pampeno C. Lin Z. Meruelo D. Potent and Targeted Sindbis Virus Platform for Immunotherapy of Ovarian Cancer Cells 2022127710.3390/cells 1201007736611875 PMC 9818975 · doi ↗ · pubmed ↗
- 2Pampeno C. Opp S. Hurtado A. Meruelo D. Sindbis Virus Vaccine Platform: A Promising Oncolytic Virus-Mediated Approach for Ovarian Cancer Treatment Int. J. Mol. Sci.202425292510.3390/ijms 2505292538474178 PMC 10932354 · doi ↗ · pubmed ↗
- 3Siegel R.L. Giaquinto A.N. Jemal A. Cancer statistics, 2024 CA Cancer J. Clin.202474124910.3322/caac.2182038230766 · doi ↗ · pubmed ↗
- 4Ferrone C.R. Ryan D.P. Pancreatic Cancer: A Time to Change Ann. Surg.20202711003100410.1097/SLA.000000000000391032398612 · doi ↗ · pubmed ↗
- 5Guo S. Contratto M. Miller G. Leichman L. Wu J. Immunotherapy in pancreatic cancer: Unleash its potential through novel combinations World J. Clin. Oncol.2017823024010.5306/wjco.v 8.i 3.23028638792 PMC 5465012 · doi ↗ · pubmed ↗
- 6Bonaventura P. Shekarian T. Alcazer V. Valladeau-Guilemond J. Valsesia-Wittmann S. Amigorena S. Caux C. Depil S. Cold Tumors: A Therapeutic Challenge for Immunotherapy Front. Immunol.20191016810.3389/fimmu.2019.0016830800125 PMC 6376112 · doi ↗ · pubmed ↗
- 7Ebelt N.D. Zamloot V. Manuel E.R. Targeting desmoplasia in pancreatic cancer as an essential first step to effective therapy Oncotarget 2020113486348810.18632/oncotarget.2774533014284 PMC 7517960 · doi ↗ · pubmed ↗
- 8Vonderheide R.H. Bayne L.J. Inflammatory networks and immune surveillance of pancreatic carcinoma Curr. Opin. Immunol.20132520020510.1016/j.coi.2013.01.00623422836 PMC 3647365 · doi ↗ · pubmed ↗