Nutritional, Phytochemical, and Biological Characterization of Peel, Pulp, and Seed Powder from the Fruits of Berberis mikuna and Berberis burruyacuensis: Potential as a Functional Ingredient
Enzo Agustín Matteucci, María Eugenia Orqueda, Mariana Leal, María Inés Isla, Mario Simirgiotis, Iris Catiana Zampini, Oscar R. Dantur, María Alejandra Moreno

TL;DR
This study explores the nutritional and health benefits of powders from two native Berberis fruits, showing their potential as functional food ingredients.
Contribution
The first evaluation of B. mikuna and B. burruyacuensis fruit powders as functional food ingredients.
Findings
Seed powders showed high nutritional value with protein, lipid, fiber, and ash content.
Pulp and peel powders contained high anthocyanin levels, especially in B. mikuna.
Powders exhibited antioxidant activity and enzyme inhibition relevant to metabolic syndrome.
Abstract
Berberis mikuna Job. (common name “mikuna”) and Berberis burruyacuensis O.R. Dantur, S. Radice, E. Giordani and Papini (common name “sacha mikuna”) are endemic native plant species from northwestern Argentina. The aim of this work was to evaluate, for the first time, the potential of the pulp, seed, and peel powders from B. mikuna and B. burruyacuensis fruits as functional food ingredients, with the purpose of adding value to these native resources and promoting their sustainable use. All powders exhibited nutritional value due to their protein, lipid, fiber, and ash content, especially the seed powder. Phenolic compounds (including xanthone, phenolic esters, coumarins, flavonoids, tannins, and anthocyanins), alkaloids, amino acids, lipids, and vitamins, totaling 33 compounds, were identified in the pulp, seed, and peel of both Berberis fruits through UHPLC-PDA-ESI-QT-MS/MS. High…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
 Figure 1
Figure 1 Figure 2
Figure 2 Figure 3
Figure 3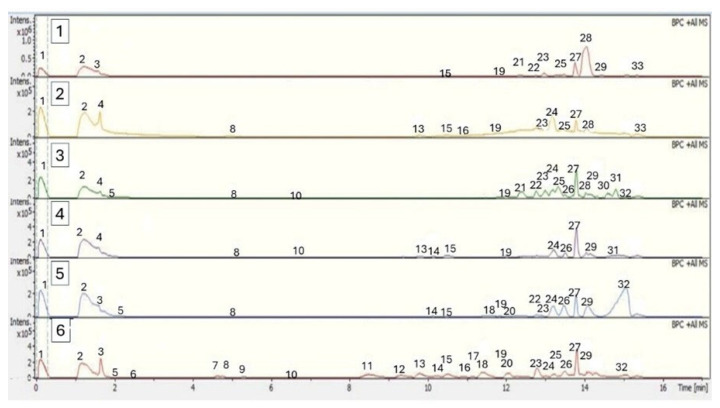 Figure 4
Figure 4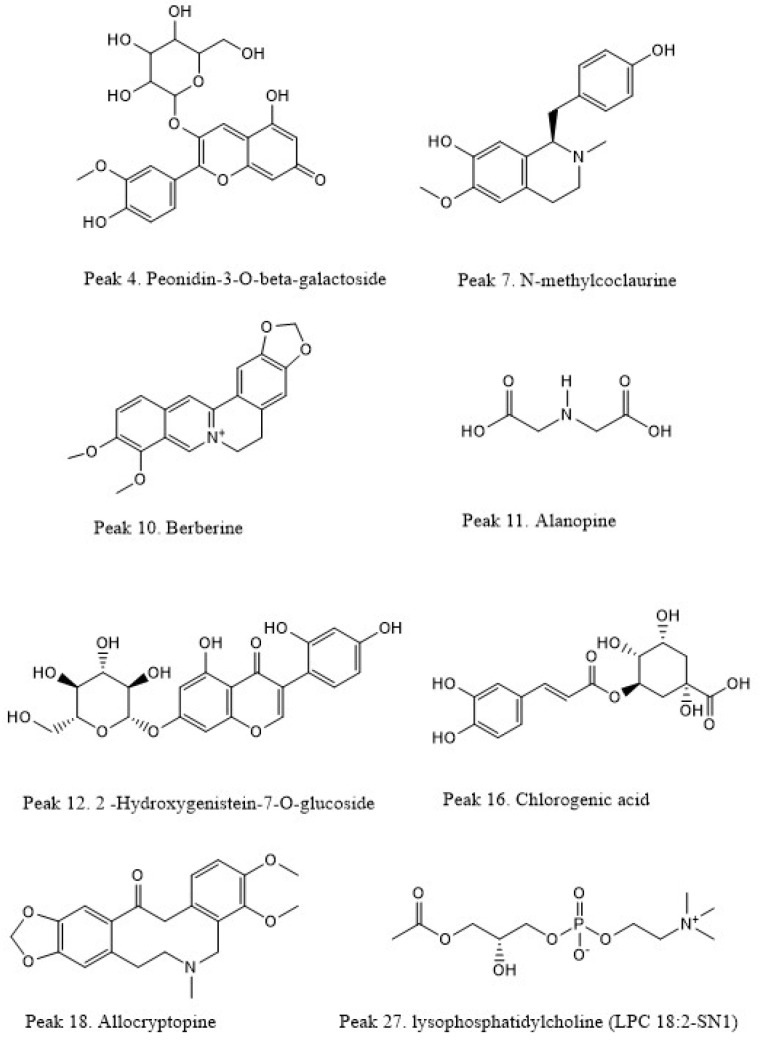 Figure 5
Figure 5- —Universidad Nacional de Tucumán
- —Agencia Nacional de Promoción Científica y Técnica
- —Consejo Nacional de Investigaciones Científicas y Técnicas
- —Fundación Williams 2023
- —CYTED
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsPhytochemicals and Antioxidant Activities · Biochemical Analysis and Sensing Techniques · Advances in Cucurbitaceae Research
1. Introduction
Berberis is the genus of the Berberidaceae family with the greatest number of species, totaling around 500, which are distributed throughout the globe. On the American continent, they extend from the north of the USA to the south of the Patagonian Andes of Argentina and Chile [1]. Approximately 26 species are recognized in Argentina and are distributed in two main areas, the Patagonian steppe and subantarctic forest and the Tucuman–Bolivian subtropical forest [2,3,4,5,6]. Berberis mikuna Job. (common name “mikuna”) and Berberis burruyacuensis O.R. Dantur, S. Radice, E. Giordani and Papini (common name “sacha mikuna”) are wild berry species endemic in the northwest of Argentina (NWA) [7]. Their dark purple or black fruits are edible (Figure 1), such as Berberis microphylla, known as calafate, and Berberis darwinii (michay). B. burruyacuensis fruits are smaller than those of B. mikuna (Figure 1). They are consumed fresh or prepared as jellies, marmalades, and wines [8]. B. mikuna was used by the Diaguita Calchaqui community as a dye, and today, their stem and roots are used by the “Warmi Pura” group in Tucumán (Argentina) and others for dyeing wool and natural fibers. Berberine, primarily stored in the roots and stem, is the alkaloid responsible for the yellow dye [8]. Most Berberis species have recognized medicinal properties due to the presence of berberine, berbamine, and phenolic compounds, as well as nutritional value, which is attributed to their fruits [3,9,10,11,12,13,14,15]. A previous study demonstrated that the fruits of B. mikuna from NWA exhibit high concentrations of anthocyanins and total phenols, comparable to those found in B. microphylla fruits from Tierra del Fuego, Argentina [16]. To date, there are no reports on the phytochemistry composition of B. burruyacuensis fruits or descriptions of the biological activities of the fruits of both species.
In this context, the objective of this study was to assess the nutritional and functional properties of the peel, pulp, and seed powders of B. mikuna and B. burruyacuensis fruits with the aim of enhancing the value of these native resources and promoting their sustainable and comprehensive utilization.
2. Results and Discussion
This study utilized B. mikuna and B. burruyacuensis fruits collected from Altos de Medina, located at 1500 m.a.s.l. in the Burruyacú department, Tucumán, Argentina (Figure 2). The pulp, peel, and seeds were separated, dried to a constant weight, and ground into fine powders. These powders were analyzed for their chromatic characteristics and nutritional composition. Ethanolic extractions were obtained from each part of the fruit (seeds, pulp, and peel), which were chemically characterized, and their functional properties were evaluated.
2.1. Chromatic Parameters of Berberis Powder
Color is one of the first parameters to come into contact with the consumer, so it is one of the most influential factors in consumer food choices, providing an idea of freshness, taste, or composition of food products, which can lead to their acceptability or rejection. This indicator develops and intensifies due to the accumulation of natural pigments (anthocyanins, betalains, carotenoids, alkaloids, flavonoids, and chlorophylls) [17]. The pulp and peel powders of both Berberis showed a violet color (Figure 3). The chromatic characteristics of the powders are shown in Table 1. The values of the parameter L* indicate medium luminosity in all fruit parts (L* ≤ 42.07). The values of parameter a* (green to red) were positive (a* values between 2.7 to 8), indicating a preponderance of red over green. The values of parameter b* (blue to yellow) were positive (b* values between 0.07 and 9.58), suggesting the primacy of yellow over blue. The peel and pulp powders of both species show b* values that are lower than those in the seed powders, indicating higher shades of blue. B. mikuna showed b* values lower than B. burruyacuensis, indicating the highest shades of blue. A natural food colorant based on anthocyanins was demonstrated in other wild barberry (Berberis vulgaris L.) fruits [18].
2.2. Nutritional and Phytochemical Composition of the Pulp, Peel, and Seed Powders of B. mikuna and B. burruyacuensis
2.2.1. Nutritional Composition of Berberis Seed, Pulp, and Peel Powders
The nutritional composition and the moisture content of the peel, pulp, and seed powders of both Berberis species are shown in Table 2. To the best of our knowledge, no previous data about these parameters have been published for B. mikuna and B. burruyacuensis. As expected, the moisture content was higher in the pulp, with no significant differences observed between the two species. The pulp powder of B. mikuna fruits showed the highest amount of soluble total sugars (37.55 g GE/100 g of powder). The values are higher than those found in B. burruyacuensis pulp or those reported by other authors for native fruits pulp powders, such as chilto [19], and similar to those found in mistol pulp powder [20]. The pulp powder of B. mikuna fruits also showed the highest amount of reducing sugars (36.8 g GE/100 g), while its seeds contained the lowest level of reducing sugars. The highest amount of protein was found in the seed powders of both species, with maximum values of 12.8% for B. mikuna and 14.9% for B. burrayucuensis. A similar protein content was found in chickpea flour [21] and in the seeds of avocado, watermelon, and melon [22]. These values are comparable to those previously reported for Berberis microphylla fruits [23] and higher than those reported by Ruiz et al. (2010) [24] for B. microphylla fruits, while the protein content in the pulp was similar to that observed in other Berberis species, such as B. asiatica (pulp: 5.5%; seed: 5.9%), B. aristata (pulp: 6.3%; seed: 6.5%), B. lycium (pulp: 6.2%; seed: 7.1%), B. jaeschkeana (pulp: 4.7%; seed: 6.1%), and B. pseudumbellata (pulp: 7.2%; seed: 8.5%) [25].
As in the case of protein, the highest amount of fat was found in the seeds of both species, exhibiting maximum values of 1.5% for B. mikuna seeds and 2.4% for B. burruyacuensis seeds. The amount of fat found in the pulp and seeds of both Berberis species was lower than those of B. asiatica (pulp: 3.1%; seed: 5.3%), B. aristata (pulp: 3.7%; seed: 5.2%), B. lycium (pulp: 2.9%; seed: 4.8%), B. jaeschkeana (pulp: 2.6%; seed: 4.7%), and B. pseudumbellata (pulp: 4.0%; seed: 4.6%) [25]. The low lipid content of B. mikuna and B. burruyacuensis seeds was similar to avocado seeds [22].
The total fiber content in all powders of both Berberis species was higher than B. asiatica pulp (8.1%) and B. pseudumbellata seeds (5.2%) [25]. The seed and peel of B. mikuna and B. burruyacuensis showed the highest levels of fiber, similar to passion fruit seeds [22] (Table 2).
The ash content of foods is often utilized as a measure of mineral levels. The total ash content of B. burrayucuensis and B. mikuna fruits was higher than reported for calafate (0.64 to 1.3%) [24]. The B. mikuna seed powder showed an ash content (1.33 g/100 g) higher than that reported for several fruits (ranging from 0.20 to 0.69 g/100 g in fruits) and similar to minor levels reported in cereals (1.62 to 3.80 g/100 g) [22,26].
The results obtained on the proximal composition of these powders indicate that seeds, which are generally undervalued and considered waste, could be used as an important source of minerals, fiber, and proteins, and that these results can encourage their consumption.
2.2.2. Phytochemical Composition
Wild fruits are known to be rich in various types of phenolic compounds, including flavonoids, anthocyanins, tannins, and phenolic acids. Phenolic compounds play a crucial role in the functional properties of fruits and contribute to the flavor, astringency, and bitterness of fruits and foods [27]. In this study, the content of total phenolic compounds (TPCs), flavonoids (F), condensed tannins (CTs), and anthocyanins in the seed, peel, and pulp powders of two Berberis species from NWA were determined for the first time (Table 3). The pulp showed TPC values of 0.78 g GAE/100 g powder (equivalent to 313.34 mg GAE/100 g FW) for B. burruyacuensis and 0.76 g GAE/100 g powder (equivalent to 268.31 mg GAE/100 g FW) for B. mikuna. Boeri et al. (2020) [23] reported a high phenolic compound content in the pulp of B. microphylla (1035.03 mg GAE/100 g FW). Similarly, TPC levels in the pulp of other Berberis species have been documented, including B. vulgaris (1052.3 mg GAE/100 g FW), B. amurensis (923.2 mg GAE/100 g FW), B. canadians (899.2 mg GAE/100 g FW), B. koreana (1362.8 mg GAE/100 g FW), and B. declinata (1243.2 mg GAE/100 g FW) [28]. The TPC content of B. mikuna seeds (24.54 g GAE/100 g DW) and B. burruyacuensis (29.83 g GAE/100 g DW) were considerably higher than the values reported for grape seeds from various cultivars, which range from 7.90 to 11.12 g GAE/100 g DW [29]. Similarly, the TPCs measured in the pulp of B. mikuna (1.48 g GAE/100 g DW) and B. burruyacuensis (1.77 g GAE/100 g DW) exceeded those previously reported for blueberry pulp from different varieties, which ranged between 0.15 and 0.69 g GAE/100 g DW [30]. In contrast, the TPC values recorded in the peel of B. burruyacuensis (1.61 g GAE/100 g DW) and B. mikuna (1.59 g GAE/100 g DW) were lower than those found in the peel of various blueberry cultivars, which showed a broader range from 2.5 to 6.9 g GAE/100 g DW [30]. The flavonoid content determined in different fruit anatomical parts of B. mikuna and B. burruyacuensis was notably higher than the values reported for grapes (Vitis vinifera) from various cultivars. In the seeds, B. mikuna and B. burruyacuensis presented values of 116.73 and 162.60 g QE/100 g DW, respectively, substantially exceeding the flavonoid content reported for grape seeds (4.00–5.20 g QE/100 g DW) [29]. In the pulp, the flavonoid content was 0.44 g QE/100 g FW in B. mikuna and 0.84 g QE/100 g FW in B. burruyacuensis, which are considerably higher than the values reported for grape pulp (0.12–0.13 mg QE/100 g FW) [31]. In the peel, B. mikuna and B. burruyacuensis exhibited values of 2.97 and 4.13 g QE/100 g DW, respectively, which greatly surpassed the flavonoid content reported for grape peels (9.8–40.0 mg QE/100 g DW) [32]. These findings underscore the high flavonoid content of Berberis fruits, highlighting their potential as valuable sources of antioxidant bioactive compounds.
The amounts of total phenolic compounds, flavonoids, and condensed tannins in the pulp and peels were similar between both species. The seeds of the fruits of both species had approximately 10 times higher TPC values than the pulp and peel, while flavonoid levels were 20 to 40 times higher than the pulp and peel (Table 3). The same trend was observed for tannins. The tannin levels were higher than those reported for other Berberis species, including B. jaeschkeana (1.4 mg/100 g), B. pseudumbellata (1.5 mg/100 g), B. aristata (1.9 mg/100 g), B. lycium (2.1 mg/100 g), and B. asiatica (2.4 mg/100 g) [27]. Tannins are commonly described as antinutritional compounds; however, they exhibit dual effects on health. On the one hand, they are considered antinutrients because they can reduce mineral absorption, inhibit digestive enzymes, and alter protein bioavailability, which may have a detrimental impact on nutrition, especially in diets high in tannins. Nevertheless, in recent years, tannins have been studied for their numerous biological benefits, including antioxidants, anti-inflammatory, neuroprotective, and anticancer properties. These benefits are mainly observed at moderate doses, demonstrating that tannins can act as functional compounds when consumed in appropriate amounts [33].
Anthocyanins were detected in all parts of both fruits; however, the pulp and peel powders of B. mikuna exhibited higher anthocyanin content than those of B. burruyacuensis. These findings align with the observed violet coloration of both powders (Table 1 and Figure 3). Similarly, previous studies have reported the incorporation of B. boliviana whole-berry powder into yogurt matrices, resulting in an attractive, stable anthocyanin-rich product that eliminates the need for industrial colorants [34]. B. mikuna exhibits a higher anthocyanin concentration compared to B. asiatica, which has been reported to contain 219 mg C3GE/100 g, highlighting its potential as a superior natural source of these bioactive pigments [35]. Levels of anthocyanins in the peel and pulp of B. mikuna and B. burruyacuensis were notably higher than the values reported for blueberries (Vaccinium corymbosum). In the pulp, B. burruyacuensis and B. mikuna presented values of 308.73 and 556.76 mg C3G/100 g DW, respectively, exceeding the anthocyanin content reported for blueberry pulp (5.8 mg C3G/100 g DW). Likewise, the anthocyanin content of B. burruyacuensis (164.95 mg C3G/100 g DW) and B. mikuna (503.80 mg C3G/100 g DW) peels were higher than the anthocyanin content reported for blueberry peel (188.5 mg C3G/100 g DW) [36].
2.2.3. Identification of Phytochemicals Using UHPLC-MS
To analyze the phytochemical composition of the Berberis powders, ethanolic extracts were prepared. Table 4 shows the retention time and MS/MS spectral data of the bioactive compounds present in the pulp, peel, and seed extracts of B. mikuna and B. burruyacuensis fruits characterized by UHPLC-MS. This study represents the first chemical characterization of these species using this methodology, resulting in the tentative identification of 33 compounds. The base peak chromatograms of Berberis extracts were obtained in the positive ion mode, as shown in Figure 4.
The identified metabolites in the pulp, seed, and peel of both Berberis species were mainly alkaloids, phenolic compounds (a xanthone, a phenolic ester, coumarins, flavonoids, anthocyanins, and tannins), amino acids, lipids, and vitamins.
Alkaloids
The main alkaloid found in Berberis was reticuline (peak 9, C_19_H_23_NO_4_), which is similar to trigonelline (peak 6, C_7_H_7_NO_2_), N-methylcoclaurine (peak 7, C_18_H_21_NO_3_), tetrahydropalmatine (peak 17, C_21_H_25_NO_4_), and alanopine (peak 11, C_6_H_11_NO_4_), which were exclusively detected in the seeds of B. mikuna.
Additionally, berberine (peak 10, C_20_H_18_NO_4_) was found in the pulp of both species and in the seeds of B. mikuna. Oxyberberine (peak 24, C_20_H_17_NO_5_) was detected in almost all fruit parts, except for the peel of B. burruyacuensis. Columbamine (peak 29, C_20_H_20_NO_4_) was found in nearly all fruit parts, except for the peel of B. mikuna. Tetrahydrocolumbamine (peak 22, C_20_H_23_NO_4_) was exclusively present in the fruit parts of B. burruyacuensis. Allocryptopine (peak 18, C_21_H_23_NO_5_) was detected in the seeds of both species. Some of these compounds, such as berberine, oxyberberine, tetrahydropalmatine, and columbamine, have been previously reported in other Berberis species [15,37]. Alkaloids are responsible for various pharmacological activities, such as antioxidants, anti-inflammatory, antiviral, antibacterial, antiparasitic, anticancer, and antiarrhythmic properties, among others. Among other compounds, berberine and its derivatives, such as oxyberberine, have been evaluated and found effective in the prevention and treatment of metabolic syndrome [15,38,39].
Amino acids
Three amino acids were tentatively identified, L-histidine (peak 2, C_6_H_9_N_3_O_2_), L-glutamic acid (peak 3, C_5_H_9_NO_4_), and tryptophan (peak 13, C_11_H_12_N_2_O_2_). L-histidine was found in all parts of the fruit and in both species, while L-glutamic acid was found in the peel of B. burruyacuensis and in the seeds of both species. Finally, tryptophan was found in all parts of the fruits of B. mikuna. These amino acids have also been reported in other Berberis fruits, such as B. heteropoda fruits [40]. In addition to their nutritional value, some biological properties, such as antioxidant activities, were attributed to these amino acids [41,42,43].
Phenolic compounds
Hydroxycinnamic acids
Chlorogenic acid (peak 16, C_16_H_18_O_9_) was detected in the peel and seeds of B. mikuna fruits, while its related compound, caffeic acid (peak 5, C_9_H_8_O_4_), was found in the seeds of both species and in the pulp of B. burruyacuensis. These compounds have also been previously identified in other Berberis fruits, such as B. microphylla (calafate), from four different locations in southern Chile [44]. The main pharmacological effects of these compounds include antioxidant, anti-inflammatory, antiviral, antibacterial, hypoglycemic, lipid-lowering, cardio-protective, anticancer, antimutagenic, and immunomodulatory effects, among others [45,46].
Coumarins
5,7-Dihydroxy-4-methylcoumarin (peak 21, C_10_H_8_O_4_) was detected in the peel and pulp of B. burruyacuensis, while mellein (peak 33, C_10_H_10_O_3_) was found exclusively in the peels of both species collected in Tucumán. B. lycium is a source of several coumarins, including 7-hydroxycoumarin (C_10_H_8_O_3_) and 4-methyl-7-hydroxycoumarin, as well as feselol. Coumarins are known for their potential therapeutic effects, including anti-inflammatory and anticoagulant properties [47].
Xanthones
Xanthones are natural polyphenolic compounds found in various plant species, as well as in fungi and lichens. Their significance in the medical and pharmaceutical fields lies in their ability to interact with multiple biological targets, endowing them with antioxidant, anti-inflammatory, and neuroprotective properties. Among their main biological activities, they exhibit anticancer, antimicrobial, anti-inflammatory, and neuroprotective effects. Specifically, they have been shown to inhibit cell proliferation in various cancer cell lines, exert activity against pathogenic microorganisms, modulate COX-2 enzyme activity to reduce inflammatory processes, and block cholinesterases, suggesting a potential application in Alzheimer’s treatment [48]. Moreollic acid (peak 32, C_34_H_41_O_9_), a xanthone, was identified for the first time in B. mikuna and B. burruyacuensis and was detected in the seeds of both species, as well as in the pulp of B. burruyacuensis. As reported by Sukpondma et al. (2005) [49], moreollic acid, derived from Garcinia hanburyi fruits, displayed antibacterial activity against methicillin-resistant Staphylococcus aureus. Additionally, Jin et al. (2019) [50] demonstrated that moreollic acid also exhibited inhibitory effects on α-glucosidase in Garcinia hanburyi fruits.
Flavonoids
Quercetin-4-O-glucoside (peak 14, C_21_H_20_O_12_) was detected in the seed extracts of both species and in the pulp of B. mikuna. Rutin (peak 15, C_27_H_30_O_16_) was identified in almost all fruit parts of both species, except for the pulp of B. burruyacuensis, whereas isorhamnetin-3-O-rutinoside (peak 23, C_28_H_32_O_16_) was present in all fruit parts of both species, except for the pulp of B. mikuna. Alpinetin (peak 25, C_16_H_14_O_4_) was identified in the peel and pulp of B. burruyacuensis and in the peel and seeds of B. mikuna.
Epicatechin (peak 20, C_15_H_14_O_6_) was detected in the seeds of both fruits. This compound was previously reported to occur in B. vulgaris [51]. Some biological activities attributed to these chemical compounds are antioxidant, anti-inflammatory, antiviral, antimicrobial, antihypertensive, vasorelaxant, neuroprotective, anticancer, hepatoprotective, cardio-protective, anti-proliferative, and anti-atherosclerotic effects, among others [52,53,54,55,56].
Isoflavones
2-Hydroxygenistein-7-O-glucoside (peak 12, C_21_H_20_O_11_), an isoflavone, was identified for the first time in B. mikuna seeds (Table 4, Figure 5).
Anthocyanins
Anthocyanins are a class of flavonoids within the polyphenol family that are responsible for the red, purple, and blue colors in many fruits and vegetables.
Peonidin-3-O-β-galactoside (peak 4, C_22_H_22_O_11_) was detected in the pulp of both Berberis species and in the peel of B. mikuna (Table 4, Figure 4). This pigment gives cherry red hue shades in plants. It has also been reported that the consumption of anthocyanin-rich foods can reduce the risk of cardiovascular diseases, improve cognitive function, and protect against cancer [56].
Procyanidins
Procyanidin B1 (peak 19, C_30_H_26_O_12_), a tannin, was detected in all anatomical parts of the fruit in both species. Tannins are polyphenolic compounds synthesized by higher plants as specialized metabolites for defense. These compounds are widely distributed in plants, particularly in roots, stems, bark, leaves, seeds, and fruits. In foods, tannins contribute to astringency and bitterness and are present in products such as coffee, cocoa, tea, red wine, nuts, and legumes. Additionally, tannins have been extensively studied for their health benefits, including antioxidant, anti-inflammatory, and antimicrobial properties, as well as their potential role in the prevention of cardiovascular diseases and certain types of cancer [57,58].
Fatty acids
The analysis of the fruits allowed the identification of various fatty acids, such as erucic acid (peak 30, C_22_H_42_O_2_), which was found exclusively in the pulp of B. burruyacuensis. Among the identified compounds, two phosphatidylcholines (PC) were detected: lysophosphatidylcholine (LPC 18:3-SN1) (peak 26, C_26_H_48_NO_7_P), identified in the pulp and seeds of both fruits, and lysophosphatidylcholine (LPC 18:2-SN1) (peak 27, C_26_H_50_NO_7_P), which was detected in all anatomical parts of both fruits. These molecules are essential components of cell membranes, contributing to their structure and functionality. Furthermore, the detection of essential fatty acids, such as linolenic acid (peak 28, C_18_H_30_O_2_) and linoleic acid (peak 31, C_18_H_32_O_2_), is particularly noteworthy.
These essential fatty acids were also reported in other Berberis species [59]. Linoleic acid is an essential fatty acid found in foods such as sunflower oil and nuts. It has anti-inflammatory properties and can improve cardiovascular health [60].
Vitamins
Ascorbic acid (peak 8, C_6_H_8_O_6_) was identified in all fruit except for the peel of B. burruyacuensis. It was also found in other Berberis species, such as B. vulgaris [61]. Some biological activities attributed to this compound include antioxidant, anti-inflammatory, and anticancer effects [62].
2.3. Functional Properties of Berberis burruyacuensis and Berberis mikuna Seed, Pulp, and Skin Powders
The biological activities of extracts obtained from Berberis powders were determined.
2.3.1. Antioxidant Activity
Table 5 shows the antioxidant activity values, as measured by ABTS^•+^ and the hydrogen peroxide scavenging values of the different fruit parts (peel, pulp, and seed) of the two Berberis species. All the powder extracts showed a dose–response effect with regard to their ABTS cation radical scavenging capacity, with SC_50_ values between 0.71 ± 0.01 and 1.80 ± 0.14 μg GAE/mL. Overall, the scavenging activity was similar to quercetin, a commercial natural antioxidant (SC_50 quercetin_ = 0.85 ± 0.04 μg/mL).
All extracts showed a high H_2_O_2_ scavenging effect (Table 5). The seed extracts of both Berberis species were more effective than the other extracts, with SC_50_ values of 24.8 and 32.4 μg GAE/mL. The B. mikuna peel extract, with an SC_50_ value of 22.4 µg GAE/mL, was able to scavenge only 37.4% of H_2_O_2_. Although hydrogen peroxide itself is not very harmful, it can generate other highly reactive species, such as HO•, which is responsible for diseases linked to oxidative, infectious, and inflammatory processes [63].
The antioxidant activity of extracts obtained from B. burruyacuensis and B. mikuna powders could be attributed to the presence of phenolic compounds identified in the present study, such as chlorogenic acid, caffeic acid, isorhamnetin derivatives, moreollic acid, rutin, procyanidin B1, and other compounds (Table 5), which demonstrated antioxidant activity [45,54,55,58]. Alkaloids such as berberine have been reported in various Berberis species and could also contribute to the antioxidant effect [15,38].
These results demonstrate that B. mikuna and B. burruyacuensis powders, or the extracts obtained from them, could be used as functional ingredients to improve the oxidative state.
2.3.2. Effect of Berberis Fruit Powder Extracts on Enzymes Involved in the Development of Metabolic Syndrome
Recent reports suggest that the development of metabolic syndrome is associated with various clinical health disorders, including oxidative stress, proinflammatory states, and non-alcoholic fatty liver disease [64]. Metabolic syndrome is characterized by the development of hypertension, hyperglycemia, and hypercholesterolemia [65]. Previous reports suggest that bioactive compounds present in so-called “red fruits”, particularly polyphenols such as phenolic acids and anthocyanins, may have beneficial effects against the development of obesity and other diseases [66]. In this context, the activity of extracts from B. mikuna and B. burruyacuensis fruit powders against key enzymes involved in metabolic syndrome was evaluated, and the results are presented in Table 6.
The search for natural products capable of inhibiting pancreatic α-amylase and α-glucosidase activity represents a therapeutic strategy for the control of postprandial hyperglycemia.
All extracts obtained from Berberis fruit powders actively inhibited both enzymes, with IC_50_ values between 2.1 and 51.6 µg GAE/mL for α-glucosidase and 3.5 and 33.6 µg GAE/mL for α-amylase (Table 6). The powdered seed extracts exhibited the highest inhibitory activity against both enzymes. This potency may be due to the greater diversity of polyphenols and alkaloids found in this study in seed powders (Table 4). The polyphenols present in these powders, such as caffeic acid, quercetin, rutin, procyanidin, isorhamnetin, and xanthones, could be responsible for this effect. Polyphenols can reach the intestine, even when diluted with other foods and digestive fluids, where they interact with digestive enzymes and modulate glycemic responses by inhibiting carbohydrate digestion [67]. Several alkaloids, such as berberine and oxyberberine, had also demonstrated inhibitory activity against these enzymes and were found in the seed powder of both plant species [15,38,39]. It is important to note that the seeds, in addition to presenting a greater diversity of phenolic compounds (Table 4), have the highest content of total phenolic compounds, flavonoids, and condensed tannins (Table 3).
The pulp and peel extracts were also active against both enzymes, although with less potency. The weaker inhibitory effect is likely due to the lower polyphenol diversity (Table 4). The anthocyanins contained in the pulp of B. mikuna and B. burruyacuensis (Table 3 and Table 4) probably influence the activity of these enzymes. Previous work indicated the inhibitory effect of anthocyanin isolates from other Berberis fruits on α-glucosidase and α-amylase [68]. Furthermore, in vivo studies report that anthocyanins present in fruits improve insulin sensitivity through complex biochemical mechanisms, thus potentially preventing diabetes. Animal models of diabetes, as well as cross-sectional studies in humans, revealed that anthocyanins reduce blood glucose levels and peripheral insulin resistance [69,70].
Pancreatic lipase breaks down triglycerides into absorbable glycerol and fatty acids. Its inhibition by drugs such as orlistat has been used to treat obesity, with numerous undesirable side effects. However, several studies have shown that polyphenols present in fruits have lipase-inhibiting effects and could constitute a healthy way to regulate fat digestion, and therefore energy intake and obesity [71]. All extracts obtained from Argentine Berberis fruit powders were potent inhibitors of pancreatic lipase, with IC_50_ values between 0.26 and 1.16 µg GAE/mL, particularly the seed extracts (Table 6). Sosnowska et al. (2022) [72] indicated that extracts obtained from chokeberry fruits rich in proanthocyanidins were potent inhibitors of the lipase enzyme, while those fractions containing mainly low-molecular-weight phenolic compounds, such as phenolic acids, flavonols, and anthocyanins, were less active. In this framework, we can conclude that the seed extracts of both Berberis were more active in inhibiting pancreatic lipase due to the presence of compounds such as condensed tannins in their composition. Polyphenol-rich extracts from Berberis fruits showed greater potency in inhibiting pancreatic lipase than extracts from Solanum betaceum fruits, both red and orange varieties, as well as from beverages made with Zuccagnia punctata extracts, blueberry juice, and lemon honey [19,73,74].
2.4. Toxicity Analysis of Berberis burruyacuensis and Berberis mikuna Seed, Pulp, and Skin Powders
The results obtained from the A. salina assay showed that B. mikuna and B. burruyacuensis powder extracts were not toxic below a concentration of 125 μg GAE/mL. Table 7 shows the results obtained in the assay evaluating mutagenic activity against Salmonella Typhimurium strains TA98 and TA100. The mutagenicity assay using S. Typhimurium strains TA98 and TA100 indicated that, up to a concentration of 500 μg GAE/plate, B. mikuna and B. burruyacuensis extracts did not induce an increase in the number of spontaneous revertants, showing, in all cases, an RM < 1.5. It can be concluded that the extracts evaluated are neither toxic nor genotoxic at the concentrations tested, which would guarantee their safe use.
3. Materials and Methods
3.1. Chemicals and Reagents
Ethanol, phenol, sulfuric acid, and H_2_O_2_ were purchased from Cicarelli (Santa Fe, Argentina). Folin–Ciocalteau reagent, AlCl_3_, gallic acid, quercetin, procyanidin B2, and ABTS^•+^ were acquired at Sigma Aldrich, St. Louis, MO, USA. α glucosidase, α amylase, lipase, p-nitrophenyl-α-D-glucopyranoside, p-nitrophenyl palmitate, and acarbose were from Sigma-Aldrich (Darmstadt, Germany). Amilokit ^®^ was purchased from Wiener Lab Group, Rosario, Argentina, Kit No. 1504163370. Orlistat (tetrahydrolipstatin, ATC code: A08AB01) was purchased from Elea Laboratory, Ciudad Autónoma de Buenos Aires, Argentina.
3.2. Plant Material
B. mikuna fruits were collected in January at the coordinates 26°23′52.8″ S 65°04′22.2″ W and B. burruyacuensis fruits were collected in February at the coordinates 26°23′52.8″ S 65°04′20.3″ W (Figure 2). Voucher specimens were deposited at the Herbarium of Fundación Miguel Lillo, Tucumán, Argentina (LIL 618421 and 618422, respectively). The identification of plant materials was carried out by the botanist Dr. Ana Soledad Cuello. The harvesting stage of fruits was selected as ripe grade as commonly consumed (Figure 1). Once transferred to the laboratory, the fruits of both species were washed with tap water and separated into peel, pulp, and seed. Subsequently, each part of the fruit was dried by freeze-drying and then ground in a Helix mill (Numak, F100 Power 1/2 HP-0.75 Kw, Brusque, Brazil) to obtain a fine powder (Figure 3). The powders were stored in sealed plastic bags at −20 °C.
3.3. Quality Parameters of Fruits
The chromatic parameters of powders from each part of the fruit (pulp, peel, and seed) were measured with a Chroma meter NR110 (3NH TECHNOLOGY Co., Ltd., Zengcheng, China) using the CIELab system. The color space was chosen to obtain the results expressed in the chromaticity coordinates L*, a*, and b* for the selected illuminant. The L* coordinate represents the lightness (black or white contribution varying between 0 and 100), a* represents the green or red contribution (negative or positive), and b* represents the blue or yellow contribution (negative or positive). The L* coordinate is perpendicular to the plane containing the chromaticity coordinates a* and b*. Considering the L*, a*, and b* coordinates, the color is expressed through L*, C*, and H, where L* is the brightness, C* is the chroma or saturation, and H is the hue (or hue angle, indicating the color variation in the plane formed by the a* and b* coordinates). These parameters were determined considering: C* = (a2 + b2) 1/2, H° = arc tang (b/a), where 0° = red-purple, 90° = yellow, 180° = bluish-green, and 270° = blue and the CIRG index= 180-h/(L* + C*) [75].
3.4. Nutritional Analysis
To 0.2 g of each one of the powders, 3 mL of 80% ethanol was added and heated at 80 °C for 10 min. The extractions were carried out until exhaustion. Then, they were centrifuged and the total soluble sugars in the supernatant were quantified using the method proposed by Dubois et al. (1956) [76]. The results were expressed in g of glucose equivalents in 100 g of powder (g GE/100 g of powder). Reducing sugars were quantified by Somogyi (1945) [77] and Nelson (1944) [78]. The results were expressed in g of glucose equivalents in 100 g of powder (g GE/100g of powder). Total protein content was determined by measuring total nitrogen (N) using the Kjeldahl method with a conversion factor of 6.25 (AOCS, 1989) [79]. The total lipid content in each of the powders was determined gravimetrically after continuous extraction of crude fat with petroleum ether at 40–60 °C in a Soxhlet for 4 h. The results were expressed in g/100 g powder.
The moisture content was determined by evaluating the difference in weight between the fresh sample and the sample dried by freeze-drying. The dried sample was placed in a muffle furnace (500 °C) until the ashes were obtained (AOAC, 2005) [80]. The results were expressed in g/100 g powder.
Crude fiber content was determined according to AOAC (2005) [80]. The powder (0.4 g) was weighed and transferred to a beaker containing 4 mL of 1.25% H_2_SO_4_. The mixture was incubated at 100 °C for 30 min. Subsequently, 10 mL of NaOH 3.52% was added, and the mixture was incubated again at 100 °C for 30 min. The sample was filtered and subsequently washed with hot distilled water until the alkali was eliminated. It was then washed with H_2_SO_4_ 1.25% and again with distilled water until the acid was eliminated. Finally, the sample was placed on ash-free filter paper in a crucible. The sample was transferred to ash-free filter paper in a pre-weighed porcelain crucible and dried at 110 °C until a constant weight. The sample was then incinerated in a muffle furnace at 500 °C, and the crude fiber content was determined by difference, with the result expressed in g/100 g powder.
3.5. Functional Phytochemicals
3.5.1. Extract Preparation
The ethanolic extractions were performed using 1 g of each part of the fruit (seeds, pulp, and peel) powders with 10 mL of 96° ethanol, assisted by ultrasound in 3 cycles of 20 min each. The extracts were then filtered and stored in caramel-colored glass bottles. The dry weight (DW) of each extract was determined by evaporating the solvent at a constant temperature of 40 °C. Dry extracts were stored frozen at −20 °C until analysis.
3.5.2. Total Polyphenols and Flavonoids
The content of total phenolic compounds was quantified using the technique described by Singleton et al. (1999) [81]. Each extract was mixed with 0.1 mL of Folin–Ciocalteu reagent (previously diluted with distilled water, 1:1; v:v) and 0.4 mL of 15.9% sodium carbonate solution. The mixture was then incubated at room temperature for 20 min. After incubation, the absorbance was measured at 765 nm using a UV–visible spectrophotometer (UV-2400 PC). The results were expressed in g of gallic acid equivalents (GAE) in 100 g of powder (g GAE/100 g powder). The flavonoid content was estimated using the method of Zhishen et al. (1999) [82]. Each extract was mixed with 0.03 mL of 5% sodium nitrite solution and allowed to stand for 5 min. Subsequently, 0.03 mL of 10% aluminum chloride solution was added, followed by a 5 min incubation period. Then, 0.2 mL of 1% sodium hydroxide solution was added. The reaction mixture was brought to the final volume with distilled water. Absorbance was then measured at 510 nm using a UV–visible spectrophotometer (UV-2400 PC). The results were expressed in g of quercetin equivalents in 100 g of powder (g QE/100 g powder).
3.5.3. Condensed Tannins and Pigments
The seed, pulp, and peel extracts were used to determine the condensed tannin and anthocyanin contents. The total condensed tannin content was quantified as described by Prior et al. (2010) [83]. The condensed tannin content was determined by mixing 30 μL of each sample with 900 μL of 0.1% DMAC (4-Dimethylaminocinnamaldehyde) in acidified ethanol and 270 μL of 80% ethanol. The mixture was incubated for 20 min at 25 °C. Absorbance was then measured at 640 nm. Results were expressed as g of procyanidin B2 equivalent (PB2E) in 100 g powder (g PB2E/100g powder) of each fruit part.
The anthocyanin content was quantified using the differential pH method according to the technique described by Lee et al. (2005) [84]. This spectrophotometric method is based on the structural transformation of anthocyanins with a change in pH (pH 1.0—colored and pH 4.5—uncolored). The samples were diluted in a solution of potassium chloride (0.025 M pH 1.0) and sodium acetate buffer (0.4 M pH 4.5) in a 4:1 ratio (buffer/sample). The solution was incubated for 20 min in the dark, and the absorbance readings were taken at 520 and 700 nm. The total anthocyanin concentration was calculated using the following formula:
where A (absorbance) = (A_520nm_ − Abs_700nm_)pH1 − (A_520nm_ − A_700nm_)pH4.5; MW = the molecular weight of cianidin-3-glucoside: 449.2 g/mol; DF = the dilution factor; ɛ = 26,900; the molar extinction coefficient for cyanidin-3-glucoside (L/mol/cm); l = the path length (cm). The results were expressed in mg of cyanidin-3-glucoside equivalents in 100 g of powder (mg C3GE/100 g of powder).
3.5.4. Identification of Phytochemicals Using UHPLC-PDA-ESI-QT-MS/MS
The separation and identification of specialized metabolites from B. mikuna and B. burruyacuensis extracts were carried out on a UHPLC-MS system equipped with a UHPLC Ultimate 3000 RS with Metaboscape 4.0 software, and a Bruker maXis ESI-QTOF-MS. The chromatographic equipment consisted of a quaternary pump, an autosampler, a thermostated column compartment, and a photodiode array detector. The elution was performed using a binary gradient system with eluent (A), 0.1% formic acid in water, and eluent (B), 0.1% formic acid in acetonitrile, and the following gradient: isocratic 12% B (0–1 min), 12–99% B (1–11 min), isocratic 99–99% B (11–13.5 min), 99–12% B (13.5–14 min), and 12–12% B (14–15 min). The separation was carried out with an acclaim Kinetex C18 1.7 μm (2.1 mm × 100 mm) column at a flow rate of 400 mL/min. ESI-QTOF-MS experiments in negative ion mode were recorded, and the scanning range was between 50 and 2000 m/z.
3.6. Antioxidant Activity of Polyphenolic Extracts
3.6.1. ABTS●+ Scavenging Assay
The total antioxidant activity of each extract was measured using the enhanced ABTS radical cation (ABTS^●+^) method, as described by Re et al. (1999) [85]. Results were expressed as the concentration of phenolic compounds in gallic acid equivalent per mL (µg GAE/mL) required to remove 50% of ABTS (SC_50_). Quercetin was used as a reference compound.
3.6.2. Hydrogen Peroxide Scavenging Assay
The ability of polyphenolic extracts from Berberis fruits to scavenge hydrogen peroxide was measured according to Fernando and Soysa (2015) [86] with some modifications. Briefly, the reaction mixture containing the polyphenol-enriched extracts (7–70.4 µg GAE/mL), H_2_O_2_ (0.7 mM), and horseradish peroxidase (1 U/mL) was preincubated for 3 min at 37 °C. The mixture containing the phenolic extract (2–50 µg GAE/mL) and H_2_O_2_ (0.7 mM) was incubated for 3 min at 37 °C. Then, a solution of phenol (12 mM) and 4-aminoantipyrine (0.5 mM) was added to the mixture. The absorbance was measured at 504 nm (Spectrophotometer Jasco v-630, Thermo Fisher Scientific, Tokyo, Japan) by the production of a colored quinone. The values were expressed as the concentration of phenolic compounds in gallic acid equivalent per mL (µg GAE/mL) required to remove 50% of hydrogen peroxide (SC_50_). Quercetin was used as a control.
3.7. Antihyperglycemic and Antihyperlipidemic Activity of Berberis Extracts
3.7.1. α-Glucosidase Inhibition
The inhibitory potential of the extracts on the glucosidase enzyme was performed according to Costamagna et al. (2013) [87] with some modifications. First, preincubation of the α-glucosidase enzyme (0.0273U) and the polyphenolic extracts of the seeds, peel, and pulp (0.5–70 µg GAE/mL) was performed in 160 µL of 0.1 M sodium phosphate buffer (pH 6.9) at 4 °C for 15 min. Then, 5 μL of 25 mM p-nitrophenyl α-D-glucopyranoside was added to start the reaction. After 15 min of incubation at 37 °C, 80 μL Na_2_CO_3_ 0.2 M was added to stop the reaction and the absorbance at 405 nm was recorded. The IC_50_ values were calculated by the interpolation of dose–response curves. Results were expressed as the concentration of phenolic compounds in gallic acid equivalent per mL (µg GAE/mL) required to inhibit 50% of enzymatic activity. Acarbose was used as a positive control.
3.7.2. α-Amylase Inhibition Assay
The inhibitory activity of α-amylase using starch as a substrate was assayed using an Amilokit^®^ according to the manufacturer’s instructions, as reported by Costamagna et al. (2016) [88]. The inhibitory capacity of polyphenolic extracts from the seeds, peel, and pulp (0.5–50 µg GAE/mL) was evaluated and the results are reported as IC_50_ values. The IC_50_ values indicate the µg GAE/mL of extract required to inhibit the enzyme by 50%. Acarbose was used as a positive control.
3.7.3. Lipase Inhibition Assay
The lipase inhibitory activity of Berberis extracts was assayed by measuring the enzymatic hydrolysis of p-nitrophenyl palmitate to p-nitrophenol according to Costamagna et al. (2016) [88]. The enzyme lipase (1 mg/mL) was preincubated with different concentrations of the extracts (final concentration between 0.1 and 4 µg GAE/mL) for 10 min. Then, sodium phosphate buffer 0.1M pH 7 supplemented with 0.6% (w/v) Triton X-100, 0.15% (w/v) gum arabic, and 20 µL of 10 mM p-nitrophenyl palmitate (substrate) was added, and the reaction was incubated at 37 °C for 20 min. The absorbance was measured in a microplate reader at 400 nm using a BiotekELx808 microplate reader. IC_50_ values were determined as µg GAE/mL of extracts required to inhibit the enzymatic activity by 50%. Orlistat was used as a positive control.
3.8. Toxicity Assessment
3.8.1. Acute Toxicity Assay
The acute toxicity level of extracts from different parts of Berberis fruits was evaluated using the crustacean Artemia salina as a test organism [89]. Artemia salina cysts were incubated in artificial seawater. After 24 h incubation at 25 °C, nauplii were transferred to microplates containing seawater and 15.6–250 µg GAE/mL of each extract. A solvent control (dimethyl sulfoxide, DMSO) without extract and a positive control of potassium dichromate (10–40 µg/mL) were included in the experiment. All plates were incubated for 24 h at 25 °C. At the end of the incubation period, dead larvae were counted for each polyphenolic extract concentration tested.
3.8.2. Mutagenicity
The mutagenic effect of Berberis extracts on two strains of Salmonella Typhimurium (TA98 and TA100) was evaluated. The plate incorporation assay was performed according to Maron and Ames (1983) [90], adding 0.1 mL of the overnight bacterial culture, 0.1 mL of each extract at different concentrations (125–500 µg GAE/plate), and 2 mL of top agar to minimal agar plates. The plates were then incubated at 37 °C for 48 h. After incubation, revertant colonies were counted and compared with the number of revertant colonies in the controls. The positive control used was 4-nitro-O-phenylenediamine (4-NPD; Aldrich Chemical Co., St. Louis, MO, USA), 10 µg/plate. Solvent control was performed by adding 0.1 mL of DMSO/plate. An extract was considered mutagenic when the mean number of revertant was two-fold or more than two-fold higher than that of the negative control. Three plates were tested per experiment and two separate experiments were carried out for each concentration tested and for the positive and negative controls.
3.9. Statistical Analysis
The statistical analysis was performed using Infostat software (Student Version, 2015) [91]. All measurements were carried out in triplicate or more, and the data are expressed as mean values accompanied by their respective standard deviations. To evaluate differences among experimental groups, a one-way analysis of variance (ANOVA) was conducted. When significant differences were detected (p < 0.05), Tukey’s post hoc test was used to identify specific group differences. A p-value below 0.05 was considered indicative of statistical significance.
4. Conclusions
This study highlights that fruit powders from B. mikuna and B. burruyacuensis, two plant species native to Tucumán, Argentina, are promising sources of nutrients (proteins, lipids, and sugars), fiber, and minerals. The nutritional attributes varied considerably among parts of fruits and between species. The powders obtained from seeds, peel, and pulp can also be considered valuable sources of functional compounds, with potential applications as dietary supplements or food additives aimed at preventing diseases associated with oxidative stress and metabolic syndrome. In addition, it is possible to achieve comprehensive use of the fruits using the seeds, which are usually discarded, since it has been shown that they have biological activities such as antioxidant and hypolipidemic effects, which can be attributed to the identified bioactive compounds. The metabolites detected in the pulp, seed, and peel powders of both Berberis species were mainly alkaloids and phenolic compounds, including phenolic acids, coumarins, flavonoids, isoflavonoids, xanthones, anthocyanins, and condensed tannins.
Overall, this work provides a foundation for promoting the cultivation and sustainable use of Berberis species in NWA.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Ahrendt L.W.A. Berberis and Mahonia: A taxonomic revision Bot. J. Linn.196157141010.1111/j.1095-8339.1961.tb 00889.x · doi ↗
- 2Job M.M. Los berberis de la región de Nahuel Huapi Rev. Mus. Plata 194252172
- 3Orsi M.C. Berberidaceae Flora Patagónica Correa M.N. INTA Buenos Aires, Argentina 1984325348
- 4IBODA Flora del Cono Sur Catálogo de Plantas Vasculares 2019 Available online: http://www.darwin.edu.ar/Proyectos/Flora Argentina/Generos.asp(accessed on 28 April 2025)
- 5Zuloaga F.O. Morrone O. Belgrano M.J. Catalogue of the Vascular Plants of the Southern Cone (Argentina, Southern Brazil, Chile, Paraguay and Uruguay). Volume 2: Dicotyledoneae: Acanthaceae-Fabaceae (Abarema-Schizolobium)Missouri Botanical Garden Press St. Louis, MO, USA 20089852286
- 6Ayarde H. Bulacio E. Una nueva especie de Berberis (Berberidaceae) de las montañas del noroeste de Argentina y sur de Bolivia Bol. Soc. Argent. Bot.20155059560010.31055/1851.2372.v 50.n 4.12918 · doi ↗
- 7Gori M. Biricolti S. Pedrazzani S. Giordani E. Papini A. Dantur O.R. Arena M.E. Radice S. Berberis burruyacuensis OR Dantur, S. Radice, E. Giordani, A. Papini sp. nov.(Berberidaceae): A new species Genet. Resour. Crop Evol.2021681799180810.1007/s 10722-020-01096-y · doi ↗
- 8Pedrazzani S. Scali E. Radice S. Giordani E. WARMIPURA: Recovery of ancestral techniques for dyeing wool and natural fibers in North-West Argentina J. Univ. Int. Dev. Coop.202044352