Functional Characterization of Anthocyanin Biosynthesis-Related Dihydroflavonol 4-reductase (DFR) Genes in Blueberries (Vaccinium corymbosum)
Yongyan Zhang, Sijian Guo, Zening Zhang, Ruide Li, Shitao Du, Siyi Hao, Chunzhen Cheng

TL;DR
This study identifies and characterizes DFR genes in blueberries, focusing on VcDFR11's role in anthocyanin production during fruit ripening.
Contribution
The study provides the first genome-wide analysis of blueberry DFR genes and identifies VcDFR11 as a key player in anthocyanin biosynthesis.
Findings
36 VcDFR genes were identified and categorized into five subfamilies.
VcDFR11 and VcDFR30 promote anthocyanin biosynthesis, with VcDFR11 being more effective.
VcMYB-1 and VcbHLHs transcription factors activate the VcDFR11 promoter, enhancing anthocyanin accumulation.
Abstract
Dihydroflavonol 4-reductase (DFR) genes contribute greatly to anthocyanin biosynthesis in plants. Up to now, however, research on the DFR gene family and the key anthocyanin-related DFR members in blueberries (Vaccinium corymbosum) has been limited. In this study, we performed a genome-wide identification of the blueberry DFR gene family, identifying 36 VcDFR genes categorized into five subfamilies. Gene expression analysis showed that three Subfamily III members (VcDFR11/29/34) and four Subfamily V members (VcDFR4/7/30/33) are highly expressed in blueberry fruits, particularly at late ripening stages. Transient overexpression analysis in apple fruits verified the contributions of VcDFR11 and VcDFR30 to anthocyanin biosynthesis, with VcDFR11 showing better promoting effects. Blueberry fruit-based transient overexpression further confirmed the promoting effects of VcDFR11 on anthocyanin…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
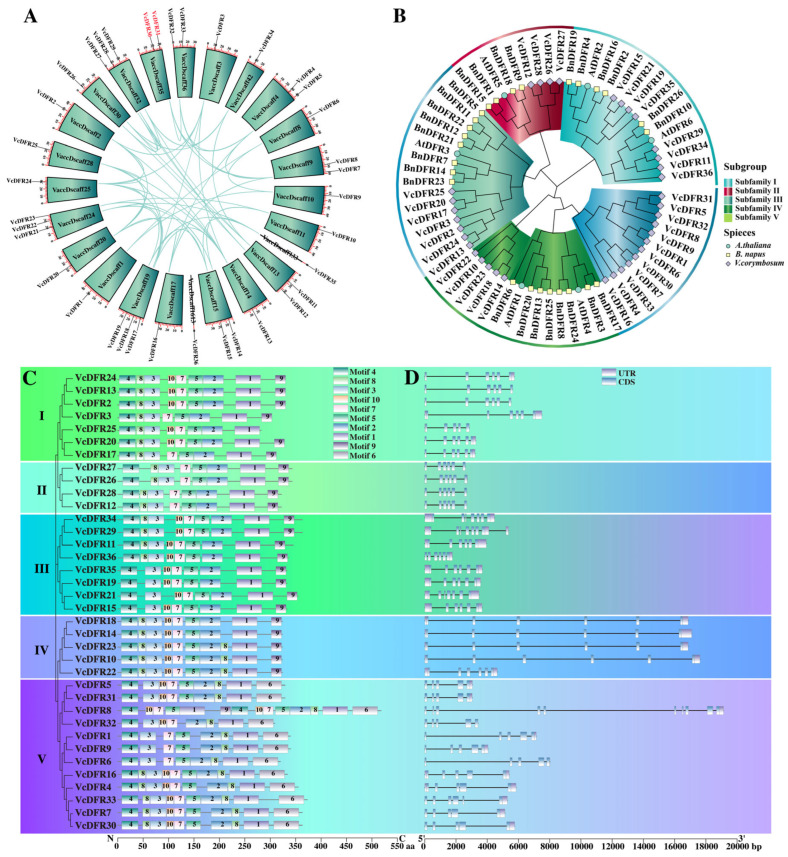 Figure 1
Figure 1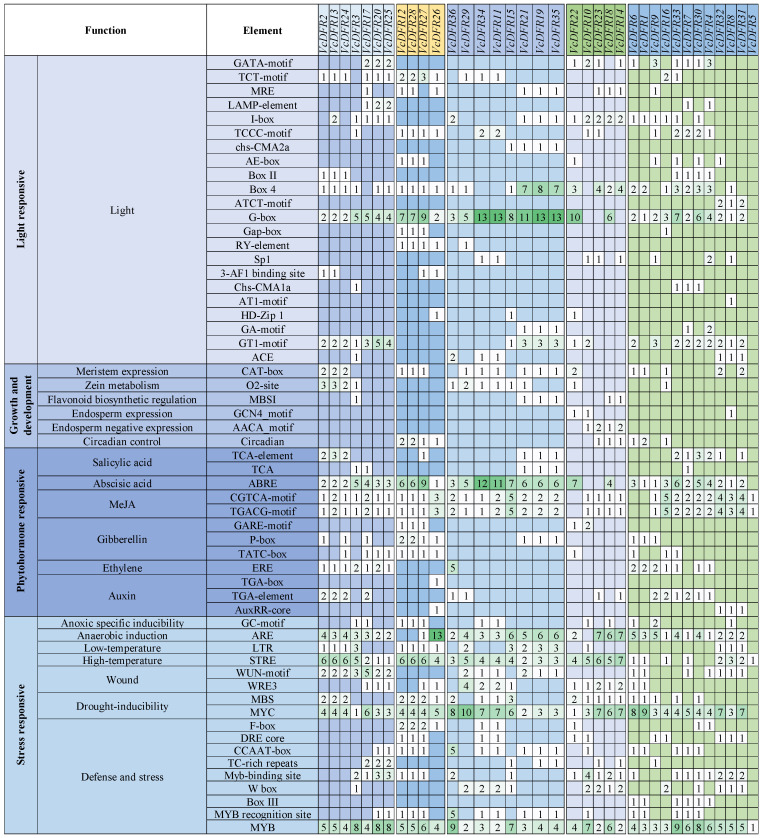 Figure 2
Figure 2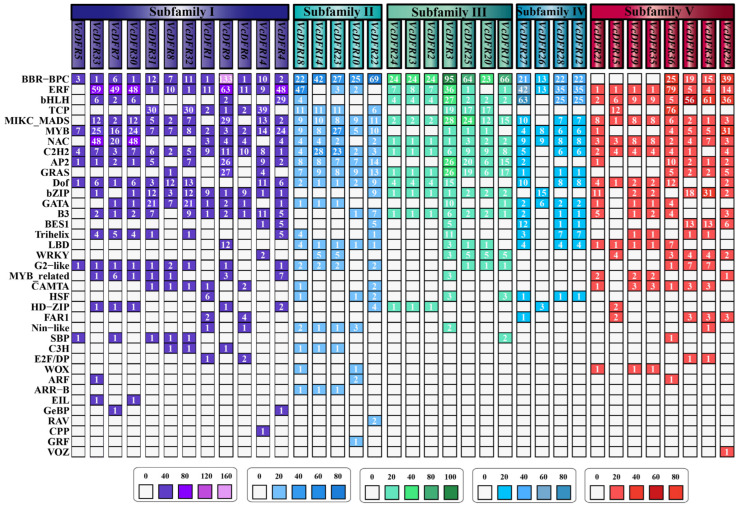 Figure 3
Figure 3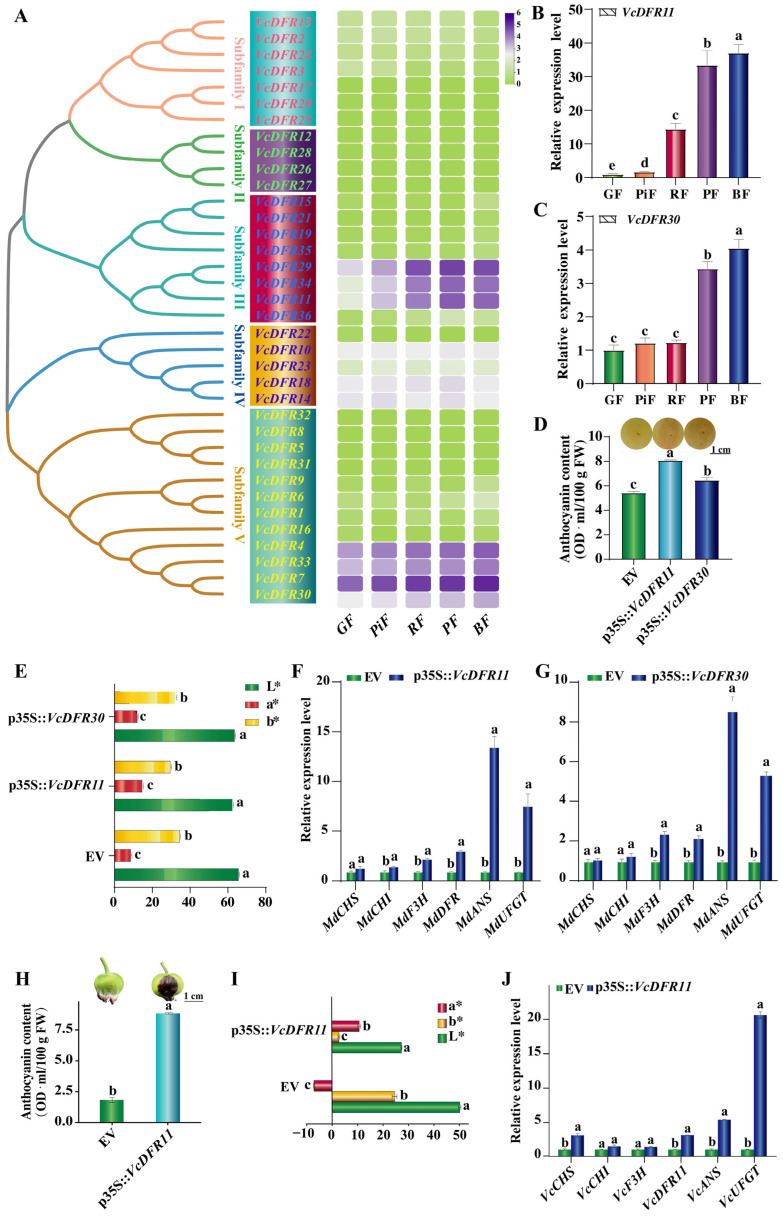 Figure 4
Figure 4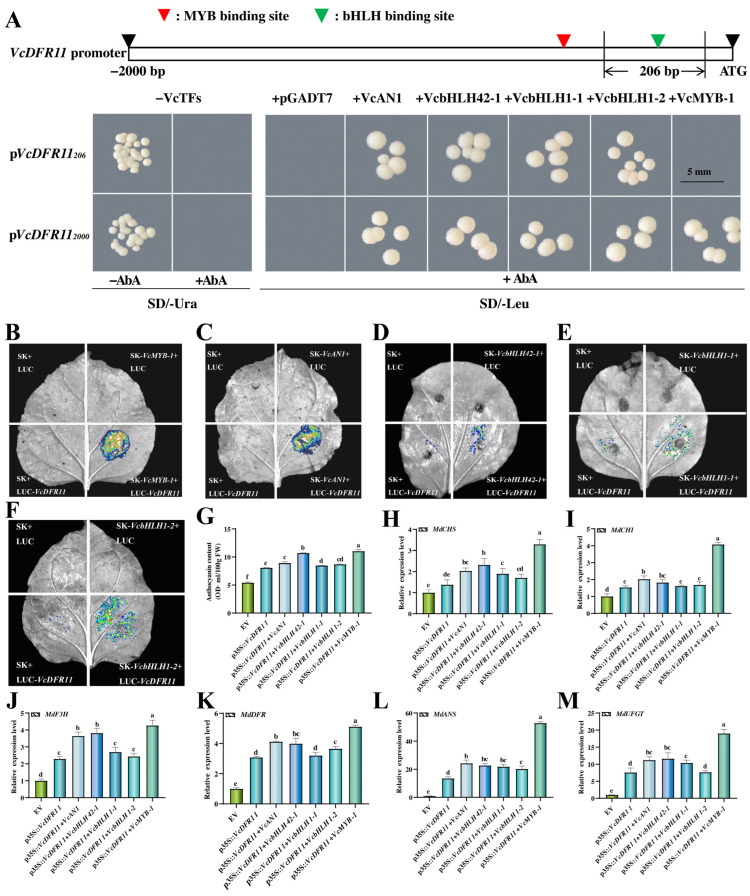 Figure 5
Figure 5- —Natural Science Basic Research Program of Shanxi Province
- —PhD Introduction Research Start-up Project of Shanxi Agricultural University
- —Fund for High-level Talents of Shanxi Agricultural University
- —Reward Fund for PhDs and Postdoctors of Shanxi Province
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsPlant Gene Expression Analysis · Phytochemicals and Antioxidant Activities · Plant biochemistry and biosynthesis
1. Introduction
Dihydroflavonol 4-reductase (DFR, EC1.1.1.219) catalyzes the conversion of dihydroflavonols to leucoanthocyanidins and is the first committed enzyme of the anthocyanin/proanthocyanidin biosynthesis pathway [1,2,3]. In recent years, DFR genes involved in anthocyanin biosynthesis have been identified and functionally characterized in numerous plant species [4,5,6]. For example, the expression level of pomegranate (Punica granatum L.) PgDFR is positively correlated with anthocyanin accumulation, and its expression in pomegranates with black skin is approximately 3- and 5-fold higher than in samples with red or green skin [4], respectively. In leaves of different Malus crabapple cultivars the expression of McDFR1 is positively correlated with the anthocyanin content [5]. In pink-leaved ornamental kale (Brassica oleracea var. acephala) the expression of BoDFR1 is very significantly higher than in the white-leaved line, and its silencing in the pink-leaved line leads to significantly reduced anthocyanin accumulation [6]. Evidence has revealed that mutations in DFR genes will cause abnormal anthocyanin accumulation in plants [6,7,8]. For instance, the Arabidopsis DFR mutant (tt3) plant shows no anthocyanidins accumulation in seeds and vegetative tissues [7]. In green-leaved kale, BoDFR1 and BoDFR2, have a 1 bp and 2 bp insertion compared to pink-leaved kale [9], respectively. Recently, the functions of several plant DFRs have been verified through genetic transformation analysis [6,10,11]. The heterologous overexpression of AtDFR in Brassica napus L. significantly increases anthocyanin accumulation [10]. Camellia sinensis CsDFRa and CsDFRc overexpression can restore the abnormal anthocyanin accumulation phenotypes of tt3 Arabidopsis plants [11]. Moreover, overexpression of B. oleracea BoDFR1 in white-leaved kale results in obvious anthocyanin accumulation [6].
Many transcription factors (TFs) have been reported to play important regulatory roles in plant anthocyanin biosynthesis and accumulation by regulating DFR transcription [12,13]. M. crabapple McMYB10 positively regulates anthocyanin biosynthesis by binding to and activating the promoter of McDFR1 and upregulating its expression [5]. In table grapes (Vitis vinifera L.), VvMYB23 and VvbHLH93 can bind to and activate the promoter of VvDFR, and upregulate its expression [14]. Dendrobium officinale DoMYB5 and DobHLH24 can both activate the promoter of DoDFR [15]. In apples, MdMYB1 has been reported to have the ability of binding to the MdDFR promoter and regulating anthocyanin biosynthesis in the pericarp [16], while MdMYB10 regulates anthocyanin biosynthesis in apple pulp by interacting with MdbHLH3/33 to improve the activity of the MdDFR promoter [17]. The interaction between MdERF38 and MdMYB1 greatly enhances the binding ability of MdMYB1 to the promoters of MdDFR and MdUF3GT, thereby increasing anthocyanin accumulation [18]. In peach (Prunus persica), PpMYB108 binds to the promoter of PpDFR and plays important roles in regulating anthocyanin accumulation in the flower [19]; and PpMYB75 functions in promoting flesh pigmentation by binding to the ‘CCGTTG’ sequence in the PpDFR promoter and upregulating its expression [20]. In addition to TFs acting as activators of anthocyanin biosynthesis, some TFs negatively regulate anthocyanin accumulation by downregulating DFR [21,22]. For example, potato (Solanum tuberosum L.) StMYB44 negatively regulates anthocyanin biosynthesis in tuber flesh via binding to and suppressing the promoter of StDFR, thereby downregulating its expression [21].
Blueberry (Vaccinium corymbosum) is a popular economic fruit crop celebrated for its anthocyanin-rich, highly nutritional and health-beneficial blue fruits [23,24,25]. There have been some studies on the role of DFR in blueberry anthocyanin biosynthesis [26]. For example, it was reported that both pre- and post-harvest UV irradiation can upregulate the expression of VcDFR and some other anthocyanin-related structural genes and increase anthocyanin accumulation in blueberry fruits [27]. The anthocyanin-related VcMYB1 can bind to and activate the promoter of VcDFR and positively regulate blueberry anthocyanin biosynthesis [28]. VcSnRK2.3 can interact with VcMYB1 and enhance the activation ability of VcMYB1 on the VcDFR promoter [29]. Research demonstrates that plant DFRs are encoded by a gene family and that different DFR members have various functions [30]. However, up to now, there has been no report focusing on the systematic analysis of the DFR gene family in blueberries. In 2015, Gupta et al. [31] published the draft genome of a blueberry, which can greatly facilitate the systematic identification and characterization of anthocyanin-related genes. In this study, we performed genome-wide identification and characterization analyses of the VcDFR gene family, and functionally characterized two VcDFRs (VcDFR11 and VcDFR30) that were highly expressed in blueberry fruits. Furthermore, the binding abilities of anthocyanin-related TFs (VcMYB-1 [32] and four VcbHLHs [33]) to the VcDFR11 promoter were studied by using yeast one-hybrid (Y1H) and dual-luciferase (LUC) assays. Additionally, to verify the possible ‘TF-DFR’ modules involving in anthocyanin biosynthesis, apple fruit-based transient overexpression was performed by co-expressing VcDFR11 with anthocyanin-related TFs. Our study will be helpful for understanding the characteristics of the blueberry DFRs and will provide a basis for clarifying the roles of DFR in anthocyanin biosynthesis.
2. Results
2.1. Identification and Physiochemical Properties Analyses Results of VcDFRs
In total, we identified 36 VcDFRs from the blueberry genome data. According to their chromosome (scaffold) localization information, they were named as VcDFR1VcDFR36 (Figure 1A). Their encoded proteins were of 282518 aa, with molecular weight ranging from 31.4456.33 kDa, and pI ranging from 5.368.59 (Supplementary Table S1). Subcellular localization prediction results revealed that 26 (72.22%), 6 (16.67%), 2 (5.56%), 1 (2.78%), and 1 (2.78%) VcDFRs were localized in Golgi apparatus, chloroplast and Golgi apparatus, cytoplasm, chloroplast and cytoplasm, and chloroplast, respectively.
2.2. Phylogenetic and Sequence Similarity Analyses Results of VcDFRs
Phylogenetic analysis of the DFRs from the blueberries Arabidopsis thaliana and B. napus successfully categorized them into five subfamilies (Figure 1B). Subfamily I contains seven VcDFRs, eight BnDFRs, and AtDFR3. Subfamily II consists of four VcDFRs, four BnDFRs, and AtDFR5. Subfamily III contains eight VcDFRs, six BnDFRs, and two AtDFRs (AtDFR2 and AtDFR6), among which VcDFR11, VcDFR29, and VcDFR34 are most closely related to AtDFR6. Moreover, all three VcDFRs share more than 70% similarity with AtDFR6 (74.53%, 70.59%, and 70.59%, respectively). Subfamily IV consists of five VcDFRs, eight BnDFRs, and two AtDFRs (AtDFR1 and AtDFR4). However, Subfamily V contains only twelve VcDFRs, indicating that this subfamily is blueberry specific. Sequence alignment results showed that the nucleotide similarities among the 36 VcDFRs ranged from 34.86% to 100% (Figure S1A), with high similarities ranging from 91.76% to 100% and from 87.31% to 100% among Subfamily II and IV members, respectively. The protein similarities of VcDFR members belonging to the five subfamilies ranged from 72.26% to 100%, 92.71% to 100%, 55.31% to 100%, 90.43% to 100%, and 45.84% to 99.72% (Figure S1B), respectively.
2.3. Synteny, Conserved Motif, and Gene Structure Analyses Results of VcDFRs
Synteny analysis identified 51 duplicated gene pairs involving 34 VcDFRs (except the Subfamily III member VcDFR36 and the Subfamily IV member VcDFR22) (Figure 1A and Supplementary Table S3), including 1 tandem duplication gene pair (VcDFR30 and VcDFR31) and 50 segmental duplication gene pairs. The Ka/Ks values of duplicated gene pairs range from 0 to 0.7146. The divergence of these duplicated gene pairs was calculated to occur at 0 million years ago (Mya) to 3.30 Mya.
In total, we identified 10 kinds of conserved motifs in VcDFRs (Figure 1C). Of them, motif1, motif2, motif4, and motif7 are contained by all VcDFRs. There are 35 (except VcDFR8), 35 (except DFR32), 34 (except VcDFR25 and VcDFR8), and 32 VcDFRs (except VcDFR15, VcDFR19, VcDFR21, and VcDFR35) containing motif3, motif5, motif9, and motif8, respectively. All Subfamily III and IV members contain motif10. Interestingly, motif6 is found to be Subfamily V-specific.
Gene structure analysis results showed that the intron numbers of VcDFRs ranged from 49 (Figure 1D). Except for one Subfamily I member (VcDFR25, containing four introns) and three Subfamily III members (VcDFR21, VcDFR29, and VcDFR34, containing six introns), all other Subfamily IIV members contain five introns. Most of the Subfamily V members, however, contain four introns.
2.4. Promoter Analysis Results
Cis-acting elements prediction results revealed that VcDFRs promoters contained many light-, phytohormone-, stress-responsive-, and growth and development-related elements (Figure 2). In total, we identified 17 kinds of light-responsive elements in VcDFRs promoters. Of the light-responsive elements, G-box element (which is also a ubiquitous regulatory DNA element that can be bound by numerous TFs, such as bHLH and bZIP) was identified in 31 (86.11%) VcDFRs promoters. There were 33 (91.67%), 31 (86.11%), 18 (50%), 18 (50%), 15 (41.67%), and 15 (41.67%) VcDFRs containing methyl jasmonate (MeJA)-, abscisic acid (ABA)-, gibberellin (GA)-, auxin-, ethylene (ET)-, and salicylic acid (SA)-responsive elements in their promoters. Notably, all of the Subfamily I members contain the ET-responsive element ERE and all of the Subfamily I, II and III members contain MeJA- and ABA-responsive elements in their promoters. There are 32 (88.89%), 32 (88.89%), 28 (77.78%), 17 (47.22%), and 12 (33.33%) VcDFRs which contain anaerobic-induction-, high-temperature-, wound-, low-temperature-, and anoxic-specific-induction-related elements in their promoters, respectively. Additionally, meristem expression-, zein metabolism-, flavonoid biosynthetic regulation- and endosperm negative expression-related elements were found in the promoters of some VcDFRs.
The transcription factor binding sites (TFBSs) in VcDFRs promoters were further predicted (Figure 3). In total, the binding sites for 37 types of TFs are identified in VcDFRs promoters. Among them, C2H2 binding sites are identified in the largest number of VcDFRs promoters (34), followed by BCR-BPC (32), ERF (32), and MIKC_MADS (32). It is worth noting that all Subfamily II, IV, and V members have MYB binding sites, all Subfamily I and III members have bHLH binding sites, all Subfamily II and III members contain ERF binding sites, and all Subfamily I~III members have NAC binding sites in their promoters.
2.5. Gene Expression Analysis of VcDFRs
The expression patterns of VcDFRs in blueberry fruits at five ripening stages were first analyzed according to our transcriptome data (Figure 4A). Only three Subfamily III (VcDFR11/29/34, sharing nucleotide similarity > 94%), four Subfamily IV (VcDFR18/14/10/23, similarity > 85%), and four Subfamily V (VcDFR4/7/30/33, similarity > 86%, the similarity between VcDFR7 and VcDFR30 is 98.90%) members express in blueberry fruits at all stages. Of them, Subfamily III and V VcDFR members express much higher than Subfamily IV members, and their expression levels in red fruit (RF), purple fruit (PF), and blue fruit (BF) are much higher than that in green fruit (GF) and pink fruit (PiF) (Figure 4A).
Given the high similarities among the fruit expressed VcDFRs from the same subfamily, we further investigated the expression patterns of two fruit highly expressed VcDFRs, VcDFR11 and VcDFR30, in blueberry fruits using quantitative real-time PCR (qRT-PCR) (Figure 4B,C). Consistent with the transcriptome data, our qRT-PCR results showed that the expression levels of these two VcDFRs in PF and BF were significantly higher than those in GF, PiF, and RF. The expression of VcDFR11 in PiF, RF, PF, and BF was 1.73-, 14.47-, 33.46-, and 37.09-fold of GF, respectively. The expression of VcDFR30 in PF and BF was 3.44- and 4.05-fold of GF, respectively.
2.6. Effects of Transient Overexpression of VcDFR11 and VcDFR30 on Anthocyanin Accumulations
By using apple fruit-based transient overexpression, we further studied the functions of VcDFR11 and VcDFR30. Seven days post agrobacteria inoculation, apples overexpressing VcDFR11 and VcDFR30 showed obvious pigmentation in inoculated areas (Figure 4D). Compared to the apple fruit overexpressing the pBI121-GFP empty vector control (EV), the anthocyanin content in peels of apple fruit overexpressing VcDFR11 and VcDFR30 increased by 48.77% and 18.97%, respectively (Figure 4D). Notably, the anthocyanin content in apple peels overexpressing VcDFR11 was found to be significantly higher than that overexpressing VcDFR30. By determining the color parameters of apple fruits, we found that the L* and b* values of apple peels overexpressing VcDFR11 were the lowest, while their a* values were the highest (Figure 4E). VcDFR11 overexpression significantly upregulated the expression of MdCHI, MdF3H, MdDFR, MdANS, and MdUFGT in apple peels (accounting for approximately 1.54-, 2.29-, 3.08-, 13.51-, and 7.60-fold of EV, respectively) (Figure 4F). Meanwhile, VcDFR30 overexpression only upregulated the expression of MdF3H, MdDFR, MdANS, and MdUFGT in apple peels (accounting for approximately 2.40-, 2.94-, 12.57-, and 7.02-fold of EV, respectively) (Figure 4G). The promoting effects of VcDFR11 on anthocyanin accumulation and the expression of most anthocyanin-biosynthesis-related structural genes are better than that of VcDFR30. These results indicated that the VcDFR11 exhibited better fruit-pigmentation- and anthocyanin-biosynthesis-promoting effects than VcDFR30. It is worth noting that the two VcDFRs’ overexpression both led to a much larger multiple of upregulation of the downstream structural genes (MdANS and MdUFGT) than upstream genes of DFR, indicating that their overexpression greatly activated the downstream anthocyanin biosynthesis.
The function of VcDFR11 was further studied by using blueberry fruit-based transient overexpression analysis. Results showed that the gene’s overexpression accelerated greatly the fruit peel pigmentation in inoculated areas, and the anthocyanin content in fruit peels overexpressing VcDFR11 increased to about 4.88-fold of EV (Figure 4H). Similarly to the results obtained in apples, blueberry peels overexpressing VcDFR11 had lower L* and b* values and a higher a* value than that overexpressing EV (Figure 4I). Moreover, VcDFR11 overexpression upregulated the expression levels of VcCHS, VcCHI, VcF3H, VcDFR11, VcANS, and VcUFGT to 3.07-, 1.47-, 1.39-, 3.01-, 5.35-, and 20.68-fold of EV (Figure 4J), respectively. Consistent with the results obtained in apples, VcDFR11 overexpression also led to a much larger multiple of upregulation of VcANS and VcUFGT genes in blueberry peels.
2.7. Anthocyanin-Related VcbHLHs and VcMYB-1 Can Bind to and Active the VcDFR11 Promoter
Promoter sequencing results showed that there were bHLH and MYB binding sites on the VcDFR11 promoter, suggesting that its expression might be regulated by these two types of TFs. By using yeast one-hybrid assay (Y1H), the binding ability of anthocyanin-related VcAN1, VcbHLH42-1, VcbHLH1-1, VcbHLH1-2, and VcMYB-1 to the VcDFR11 promoter was first studied. Results showed that the growth of Y1HGold yeast strains transformed with pVcDFR11_206_/pVcDFR11_2000_ was inhibited on SD/-Ura plates supplemented with AbA (Figure 5A), indicating that pVcDFR11_206_ and pVcDFR11_2000_ could not be self-activated. The yeast strains co-transformed with pVcDFR11_206_ and pGADT7/pGADT7-VcMYB-1, and pVcDFR11_2000_ and pGADT7 could not grow on SD/-Leu medium containing AbA. Meanwhile, yeast strains co-transformed with pVcDFR11_206_/pVcDFR11_2000_ and pGADT7-VcAN1/VcbHLH42-1/VcbHLH1-1/VcbHLH1-2/VcMYB-1 grew well. These results indicated that VcMYB-1 and the four VcbHLHs can bind to the VcDFR promoter. Consistently, our dual-luciferase (LUC) assay results also showed that VcMYB-1 and all the four anthocyanin-related VcbHLHs can enhance the fluorescence of VcDFR11 promoter, indicating that they can bind to and activate the promoter of VcDFR11 (Figure 5B–F). Notably, the binding and activation activities of VcMYB-1 and VcAN1 to the VcDFR11 promoter were much stronger than other VcbHLHs. In one of our previous studies we have verified the interaction between VcMYB-1 and VcAN1 [25]. In this study, by using a firefly luciferase complementation imaging (LCI) assay the interactions between VcMYB-1 and VcbHLH42-1/VcbHLH1-1/VcbHLH1-2 were also verified (Figure S2), suggesting that the ‘MYB-bHLH-DFR’ module contributes greatly to the anthocyanin biosynthesis in blueberry.
2.8. Co-Overexpression of Anthocyanin-Related TFs with VcDFR11 Exhibited Better Anthocyanin Accumulaiton-Promoting Effects than Overexpressing VcDFR11 Alone
Transient overexpression experiments were further conducted to investigate the influences of anthocyanin-related TFs and VcDFR11 co-overexpression on anthocyanin accumulation. Results showed that their co-expression resulted in significantly higher anthocyanin accumulations in apple peels than that overexpressing VcDFR11 alone (Figure 5G). qRT-PCR analysis showed that the co-transformation of VcDFR11 with VcAN1, VcbHLH42-1, VcbHLH1-1, VcbHLH1-2, and VcMYB-1 further significantly upregulated the expression levels of anthocyanin-biosynthesis structural genes, particular MdANS and MdUFGT (with much higher fold changes than other structural genes), in apple peels (Figure 5H–M). Among them, the co-expression of VcMYB-1 and VcDFR11 exhibited the best promoting effect on anthocyanin accumulation and structural genes’ expression in apple fruit peels, followed by VcAN1/VcbHLH42-1.
3. Discussion
In this study, we identified 36 DFR genes from the blueberry genome, which was more than most other plants, such as Arabidopsis, B. napus [34], tea plant [11], and Freesia hybrida [35]. In B. napus, the expansion of the DFR gene family was caused by segmental and tandem duplications [34]. Similarly, we identified 50 segmental and 1 tandem duplicated VcDFR gene pairs in blueberry, indicating that segmental and tandem duplications, especially segmental duplications, promoted the expansion of the blueberry DFR gene family.
Of the 36 VcDFRs, three Subfamily III and four Subfamily V VcDFRs were expressed highly in fruits. It is worth noting that the three Subfamily III VcDFRs shared a close relationship with the anthocyanin-related AtDFR6 [10,11]. Among them, VcDFR11 encodes a protein sharing the highest similarity with AtDFR6 (74.53%). The transcriptional levels of DFR genes are highly correlated with the accumulation of anthocyanin in plants [36,37]. Our study found that almost all fruit highly expressed VcDFRs had higher expression at late ripening stages. Moreover, the VcDFR11 and VcDFR30 overexpression significantly increased anthocyanin accumulation and upregulated anthocyanin-biosynthesis-related structural genes, particularly ANS and UFGT genes. The VcDFR11 reported in our study shared a 99.52% similarity with the VcDFR reported by Wang et al. [29], indicating that VcDFR11 is a key DFR involved in anthocyanin biosynthesis in blueberry. Although the promoting effects of VcDFR30 on anthocyanin biosynthesis are milder than that of VcDFR11, its overexpression also significantly improves anthocyanin accumulation and the corresponding structural genes’ expression. Therefore, it can be concluded that fruit highly expressed Subfamily III and V VcDFRs play important roles in the anthocyanin biosynthesis of blueberries.
The expression of anthocyanin-related genes is influenced by environmental factors and transcription factors [38,39]. Many studies have revealed that the high accumulation of flavonoids/anthocyanin under stress conditions, which results from activated DFR expression or its overexpression, is correlated with the stress tolerance of plants [10,40,41]. In this study, we identified many phytohormone- and stress-responsive elements in VcDFRs promoters, indicating that the expression of VcDFRs is influenced by a variety of phytohormones and environmental factors and that they might play important roles in blueberry stress responses.
Evidence has revealed that TFs, such as MYB and bHLH, participate in anthocyanin-biosynthesis regulation by binding to and activating/suppressing promoters of anthocyanin-biosynthesis-related structural genes [42,43], including DFR. Lu et al. [14] found that VvMYB3 and VvbHLH93 functioned in anthocyanin accumulation regulation in grape flesh by binding to the promoter of VvDFR and upregulating its transcription. Liu et al. [44] found that E3 ubiquitin ligase BoMIEL1 mediates the degradation of BoMYB4b, thereby alleviating the inhibitory effect of BoMYB4b on the BoDFR1 promoter and promoting anthocyanin accumulation in kale. In this study, it was found that all Subfamily II, IV, and V VcDFR members process MYB binding sites on their promoters. Additionally, all Subfamily I and III VcDFR promoters have bHLH binding sites. Notably, both bHLH and MYB binding sites are present in the VcDFR11 promoter. Our Y1H and LUC analysis results showed that anthocyanin-related VcbHLHs and VcMYB-1, especially VcAN1 and VcMYB-1, could bind to and activate the VcDFR11 promoter. The co-overexpression of their encoding genes with VcDFR11 led to a higher accumulation of anthocyanins than overexpressing VcDFR11 did alone. Specifically, the co-overexpression of VcMYB-1 and VcDFR11 showed the best promoting effect on anthocyanin accumulation, followed by VcAN1 and VcbHLH42-1. Future studies should further investigate their regulation on VcDFR11.
4. Materials and Methods
4.1. Plant Materials
Green, pink, red, purple, and blue fruits used in this study were harvested from two-year-old ‘FL03’ plants. For apple and blueberry fruit-based transient overexpression analysis, bagged ‘Gala’ apples and green ‘Legacy’ blueberry fruits were used. For LUC and LCI assays, six-week-old Nicotiana benthamiana plants were used.
4.2. Identification of Blueberry DFR Proteins
Arabidopsis DFR sequences (downloaded from TAIR, https://www.arabidopsis.org (accessed on 26 February 2024)) were used as queries to BLASTp against blueberry genome data (downloaded from GDV, https://www.vaccinium.org/analysis/49/ (accessed on 1 March 2024)) (E-value ≤ l × 10^−5^, identity ≥ 50%) [45]. Then, CDD (https://www.ncbi.nlm.nih.gov/Structure/cdd/wrpsb.cgi (accessed on 1 March 2024)) was used to verify the presence of the DFR domain (pfam01370) in candidate DFRs. Sequences containing complete DFR domain remained, with an exception of VaccDscaff12-snap-gene-66.28 whose coding sequence (CDS) was found to be quite different from the reference sequence in the genome data by gene cloning and sequencing validations (its CDS was only 1011 bp and had a very high similarity (>98.22%) with the VaccDscaff1613-processed gene-0.0 sequence).
4.3. Physiochemical Property, Subcellular Localization, and Phylogenetic Analyses of VcDFRs
The ExPASy—ProtParam tool (https://web.expasy.org/protparam/ (accessed on 1 March 2024)) and Plant-mPLoc (http://www.csbio.sjtu.edu.cn/bioinf/plant-multi/ (accessed on 1 March 2024)) [46] were used to predict the physiochemical properties and subcellular localization of VcDFRs, respectively. MEGA11 was first used to align the DFRs from blueberries, Brassica napus, and Arabidopsis. Then, a phylogenetic tree was constructed using the neighbor-joining method (bootstrap = 1000 and other parameters set as default). For the visualization of the phylogenetic tree, the ggtree package in R was used.
4.4. Synteny, Gene Structure and Conserved Motif Analyses
Based on the genome sequence and annotation data of blueberries, MCScanX was used to analyze the syntenies among the 36 VcDFRs, and ‘Circos’ of TBtools software v2.056 was used for figure drawing [47]. The simple Ka/Ks calculator (NG) program in TBtools software v2.056 was used to calculate the divergence time (T) of duplicated VcDFR gene pairs [48]. GSDS (http://gsds.cbi.pku.edu.cn/ (accessed on 1 March 2024)) and MEME (https://meme-suite.org/meme/index.html (accessed on 1 March 2024)) were used to analyze the gene structures and conserved motifs in their encoded proteins, respectively. TBtools was used for the figure drawing of gene structures and conserved motifs analyses results [49].
4.5. Promoter Analysis
The 2000 bp sequences upstream of the start codon (ATG) of VcDFRs were extracted from blueberry genome data using TBtools, and used as promoter sequences. For the prediction analysis of cis-acting elements and transcription factor binding sites in VcDFRs promoters, PlantCARE (http://bioinformatics.psb.ugent.be/webtools/plantcare/html/ (accessed on 1 March 2024)) and PlantTFDB (http://planttfdb.cbi.pku.edu.cn/ (accessed on 1 March 2024)) were used (p-value ≤ l × 10^−5^), respectively. Subsequently, an R package was utilized to show the distribution of cis-acting elements and transcription factor binding sites across the VcDFRs promoters.
4.6. RNA Isolation and Gene Cloning
Using an RNAprep pure plant kit (Cat#GDP432) (Tiangen, Beijing, China), we isolated total RNA samples of ‘FL03’ blueberry fruits at five ripening stages. The RevertAid first-strand cDNA synthesis kit (Catalog# K1622) (Thermo Scientific, Shanghai, China) was used for the biosynthesis of cDNA used for gene cloning, with equally weighted mixed fruit RNA as the template. Then, VcDFR11 and VcDFR30 CDSs were amplified using reverse transcription PCR (RT-PCR). The 20 μL RT-PCR system contains 10.0 μL Dream Taq™ green PCR master mix (2×), 7.0 μL ddH_2_O, 1 μL cDNA, and 1 μL each of forward and reverse primers. The amplification conditions were initial denaturation at 95 °C for 3 min, followed by 35 cycles (95 °C for 30 s, 56 °C for 30 s, 72 °C for 1 min), and final extension at 72 °C for 10 min. Target PCR products were gel purified and sent to Qingke Biotechnology (Fuzhou, China) Co., Ltd. for sequencing verification.
4.7. Gene Expression Analysis
To study the expression patterns of VcDFRs during fruit ripening, transcriptome data of ‘FL03’ blueberry fruits at green, pink, red, pink, and blue stages were first used. Then, quantitative real-time PCR (qRT-PCR) was used to verify the expression patterns of two highly expressed VcDFRs, VcDFR11 and VcDFR30, in blueberry fruits at the five ripening stages. An RNAprep pure kit (Tiangen, Beijing, China) was used to isolate total RNA from fruits. High-quality blueberry fruit RNA was reverse transcribed into complementary DNA (cDNA) using the PrimeScript RT reagent kit with gDNA eraser (perfect real-time) kit (Cat# RR047A) (Takara, Dalian, China). For the designing of the primers used for the qRT-PCR analysis of VcDFR11 and VcDFR30, Primer3web version 4.1.0 (https://primer3.ut.ee/) was used. qRT-PCR reactions were conducted on a QuantStudio 3 real-time PCR system (Applied Biosystems, Shanghai, China) with GAPDH (AY123769) as the internal reference gene [25]. The relative expression levels of VcDFR11 and VcDFR30 in different fruit samples were calculated using the 2^−ΔΔCt^ method [50]. For the gene expression analysis, three biological replications were used.
4.8. Vector Construction and Transient Overexpression Assays in Apple and Blueberry Fruits
According to Zhang et al. [32], the full-length CDSs of VcDFR11 and VcDFR30 (1038 bp and 1092 bp, respectively) were introduced into the pBI121-GFP vector to obtain p35S::VcDFR11 and p35S::VcDFR30 recombinant vectors, respectively. Primers used for vector construction are all listed in Supplementary Table S2. The VcMYB-1 and VcbHLHs overexpression vectors used for co-expression experiments were provided by our lab [33,51].
After transforming overexpression vectors into Agrobacterium tumefaciens GV3101 component cells, infiltration solutions were prepared and injected into ‘Gala’ apple fruits [51,52] and green ‘Legacy’ blueberry fruits. For the transient overexpression of each vector, at least eight apple/blueberry fruits were used. After injection, fruits were kept in dark for 2 d and then removed to normal light condition at 25 °C for 5 d. The influences of the transient overexpression of selected VcDFR genes or gene combinations on fruit coloration changes were observed and photographed. A CR8 colorimeter (3nh, Guangzhou, China) was used to measure the color parameters of apple and blueberry fruits.
The anthocyanin contents in apple and blueberry fruit peels around the injection areas were determined. Briefly, fruit peels were homogenized with acidified ethanol, placed in the dark at approximately 25 °C for 1 d, and centrifuged at 10,000 rpm for 15 min to collect the supernatant. Then, the absorbance value of the supernatant solution was measured at 535 nm using a spectrophotometer (UV-1800, Shanghai Meixi Instrument Co., Ltd., Shanghai, China) and used for the calculation of the anthocyanin contents according to Yang et al. [53]. To investigate the influences of the VcDFRs overexpression on the expression of anthocyanin-biosynthesis-related structural genes, the relative expression levels of MdCHS, MdCHI, MdF3H, MdDFR, MdANS, and MdUFGT in apple peels and VcCHS, VcCHI, VcF3H, VcDFR11, VcANS, and VcUFGT in blueberry peels were analyzed using qRT-PCR [54]. Three biological replications were used for the anthocyanin content determination and qRT-PCR analysis.
4.9. DNA Isolation, VcDFR11 Promoter Cloning, and Yeast One Hybrid (Y1H)
By using a plant genomic DNA kit (Cat# GDP304) (Tiangen, Beijing, China), genomic DNA (gDNA) was isolated using ‘FL03’ blueberry leaves as materials. By using ‘FL03’ blueberry gDNA as template, a 206 bp (only containing bHLH binding site) and an approximately 2000 bp (containing binding sites for both bHLH and MYB TFs) long VcDFR11 promoter sequences were individually amplified, gel purified, and subjected to sequencing verifications. Then, the gel-purified short and long VcDFR11 promoter sequences, pVcDFR11_206_ and pVcDFR11_2000_, were individually ligated into the pAbAi vector to obtain recombinant pVcDFR11_206_ and pVcDFR11_2000_ vectors and transformed into yeast strain Y1HGold. For Y1H assay, pGADT7-VcAN1/VcbHLH42-1/VcbHLH1-1/VcbHLH1-2/VcMYB-1 (provided by our lab, Zhang et al. [25,51]) vectors were separately transformed into yeast strains Y1HGold-pVcDFR11_206_/VcDFR11_2000_ according to Zhang et al. [51].
4.10. Dual-Luciferase Assay (LUC)
By using a ready-to-use seamless cloning kit (Cat# B632219) (Sangon Biotech, Shanghai, China), the 2000 bp VcDFR11 promoter sequences were introduced into the pNC-Green-LUC vector to obtain the LUC-pVcDFR11_2000_ recombinant vector. CDSs of VcMYB-1 and VcbHLHs were introduced into pNC-Green-SK to obtain the SK-VcMYB-1 and SK-VcAN1/bHLH42-1/bHLH1-1/bHLH1-2 recombinant vectors. These recombinant vectors were individually transformed into A. tumefaciens GV3101 (pSoup-p19) cells and cultured till the OD_600_ reached 1. Then, agrobacteria carrying SK-VcMYB-1/VcAN1/bHLH42-1/bHLH1-1/bHLH1-2 were individually equally volume mixed with agrobacteria carrying LUC-VcDFR11_2000_, kept still at room temperature for 20 min, and inoculated into fully expanded healthy mature leaves of six-week-old N. benthamiana plants. pNC-Green-LUC + pNC-Green-SK, SK-VcMYB-1/VcAN1/bHLH42-1/bHLH1-1/bHLH1-2 + pNC-Green-LUC, and pNC-Green-SK + LUC-VcDFR11_2000_ inoculation combinations were also made. After inoculation, tobacco plants were kept in the dark at 25 °C for 2 d, removed to normal light conditions for another 2 d, and sprayed with 1 mM luciferin substrate solution and kept in the dark for 5 min. Then, tobacco leaves were harvested and observed under a Tanon 5200 system (Tanon Science & Technology Co., Ltd., Shanghai, China).
4.11. Firefly Luciferase Complementation Imaging (LCI)
In one of our previous studies [25], LCI was successfully applied to verify the interactions between VcMYB-1 and VcAN1. In this study, VcbHLH42-1/bHLH1-1/bHLH1-2 was individually introduced into the pCAMBIA1300-cLUC vector to obtain the cLUC-VcbHLH42-1/bHLH1-1/bHLH1-2 recombinant vectors, transformed into agrobacteria GV3101 (pSoup-p19), and subjected to LCI assay to investigate the interactions between VcbHLH42-1/bHLH1-1/bHLH1-2 and VcMYB-1 according to Yan et al. [55].
4.12. Statistical Analyses
The anthocyanin contents and relative gene expression levels in different samples were presented as mean ± standard deviations of at least three replications. SPSS 22.0 (IBM corporation, Armonk, NY, USA) was used to analyze the difference significance of measured parameters among different samples at the p < 0.05 level. For figure drawing, GraphPad Prism 8 was used.
5. Conclusions
In this study, we functionally characterized two members (VcDFR11 and VcDFR30) of the 36 blueberry VcDFR genes. The two of VcDRFs’ overexpression both significantly promote anthocyanin accumulation and upregulate corresponding structural genes’ expression (especially the downstream ANS and UFGT genes), with the former one showing much better anthocyanin-biosynthesis-promoting effects. Moreover, we found that the ‘MYB-bHLH-DFR’ module contributed greatly to anthocyanin biosynthesis in blueberries. Our study demonstrated the role of VcDFR11 in anthocyanin biosynthesis and can provide basis for its application in blueberry genetic breeding.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Johnson E.T. Ryu S. Yi H. Shin B. Cheong H. Choi G. Alteration of a single amino acid changes the substrate specificity of dihydroflavonol 4-reductase Plant J.20012532533310.1046/j.1365-313x.2001.00962.x 11208024 · doi ↗ · pubmed ↗
- 2Cheng C.-Y. Krishnakumar V. Chan A.P. Thibaud-Nissen F. Schobel S. Town C.D. Araport 11: A complete reannotation of the Arabidopsis thaliana reference genome Plant J.20178978980410.1111/tpj.1341527862469 · doi ↗ · pubmed ↗
- 3Liu H. Shi G. Ye W. Behera J.R. Kilaru A. Wang L. Functional role of DFR genes in various blue Iris for the regulation of delphinidin synthesis Plant Physiol. Biochem.202521910935510.1016/j.plaphy.2024.10935539708701 · doi ↗ · pubmed ↗
- 4Sepideh R. Bahman Z. Farhad N.-F. Ali S. Expression analysis of anthocyanin biosynthesis key regulatory genes involved in pomegranate (Punica granatum L.)Sci. Hortic.20151868488
- 5Tian J. Chen M.-c. Zhang J. Li K.-t. Song T.-t. Zhang X. Yao Y.-c. Characteristics of dihydroflavonol 4-reductase gene promoters from different leaf colored Malus crabapple cultivars Hortic. Res.201741707010.1038/hortres.2017.7029263792 PMC 5727492 · doi ↗ · pubmed ↗
- 6Feng X. Zhang Y. Wang H. Tian Z. Xin S. Zhu P. The dihydroflavonol 4-reductase Bo DFR 1 drives anthocyanin accumulation in pink-leaved ornamental kale Theor. Appl. Genet.202113415916910.1007/s 00122-020-03688-933011819 · doi ↗ · pubmed ↗
- 7Shirley B.W. Hanley S. Goodman H.M. Effects of ionizing radiation on a plant genome: Analysis of two Arabidopsis transparent testa mutations Plant Cell 19924333347135400410.1105/tpc.4.3.333PMC 160133 · doi ↗ · pubmed ↗
- 8Wang X. Chen X. Luo S. Ma W. Li N. Zhang W. Tikunov Y. Xuan S. Zhao J. Wang Y. Discovery of a DFR gene that controls anthocyanin accumulation in the spiny Solanum group: Roles of a natural promoter variant and alternative splicing Plant J.20221111096110910.1111/tpj.1587735749258 · doi ↗ · pubmed ↗