Research Progress of CLE and Its Prospects in Woody Plants
Zewen Song, Wenjun Zhou, Hanyu Jiang, Yifan Duan

TL;DR
This review summarizes the role of CLE peptides in plant development, focusing on their signaling functions in various meristems and their potential in woody plants.
Contribution
The paper provides a comprehensive overview of CLE gene functions in different plant meristems and highlights their evolutionary conservation.
Findings
CLE genes regulate multiple meristems, including root and shoot apical meristems.
CLE peptides are involved in various signaling pathways and developmental processes.
The study emphasizes the need for further research on CLE functions in tall trees.
Abstract
The peptide ligands of the CLAVATA3/EMBRYO SURROUNDING REGION-RELATED (CLE) family have been previously identified as essential signals for both short- and long-distance communication in plants, particularly during stem cell homeostasis, cell fate determination, and growth and development. To date, most studies on the CLE family have focused on model plants and especially those involving stem and apical meristems. Relatively little is known about the role of CLE peptides in tall trees and other plant meristems. In this review, we summarize the role of CLE genes in regulating plant Root Apical Meristem (RAM), Shoot Apical Meristem (SAM), Procambium, Leaf and Floral Meristem (FM), as well as their involvement in multiple signaling pathways. We also highlight the evolutionary conservation of the CLE gene family and provide a comprehensive summary of its distribution across various plant…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
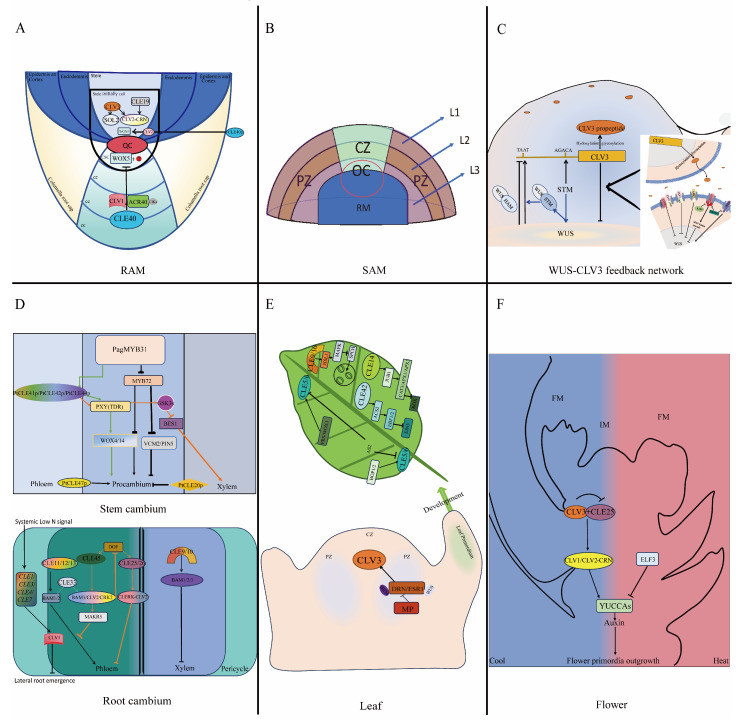 Figure 1
Figure 1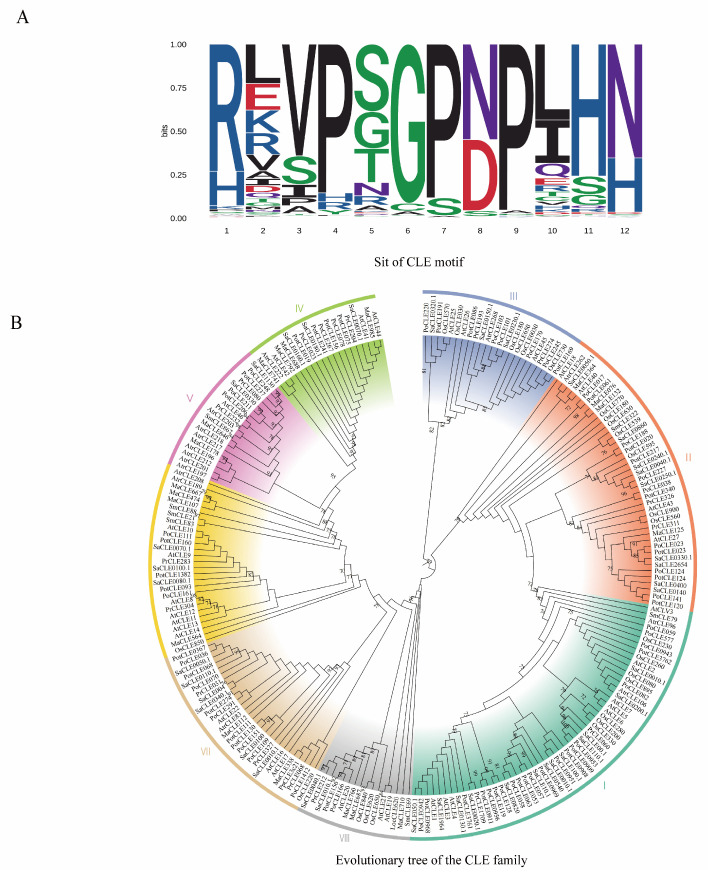 Figure 2
Figure 2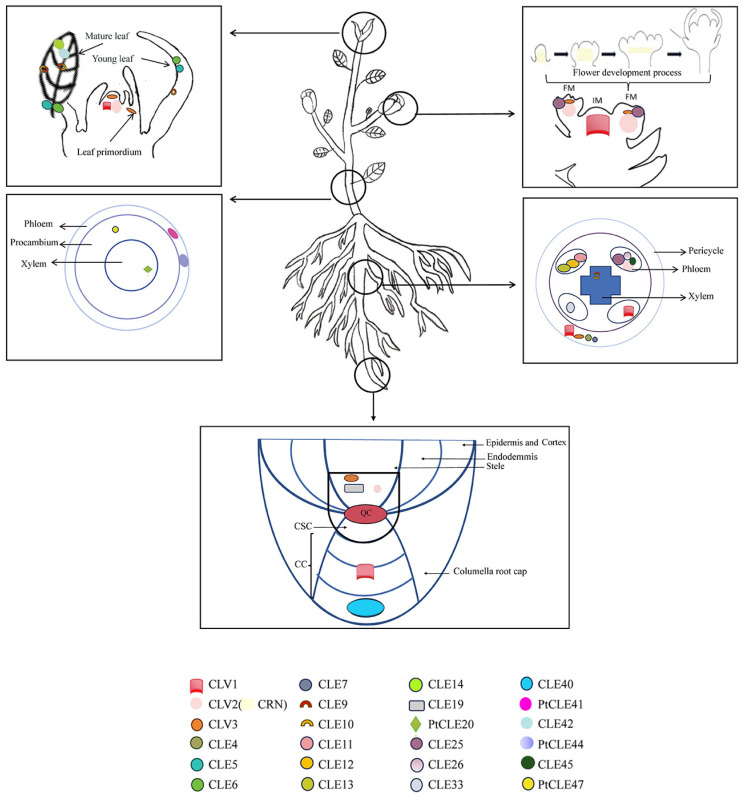 Figure 3
Figure 3- —Jiangsu Higher Education Institutions (PAPD)
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsPlant Molecular Biology Research · Plant Reproductive Biology · Plant Gene Expression Analysis
1. Introduction
The growth of trees originates from the development of embryonic stem cells within the seed. When plants have fully matured and sensed the appropriate external conditions, the differentiation and division of embryonic stem cells are precisely and stably regulated, which is the basis for the formation and maintenance of meristem tissues for sustaining development of plants after embryo [1,2]. The radicle firstly divides into RAM, and with the elongation of the plumular axis, the germ differentiated SAM to form stems and leaves, and then the procambium cells continued to increase differentiation. Eventually, the plant transitions from vegetative growth to reproductive development, marked by the formation of the FM [3,4,5]. In recent years, many genes have been concerned with the regulation of plant meristem, such as FASCIATA1 and FASCIATA2 (FAS1/FAS2), SHOOT MERISTEMLESS (STM), and CLAVATA3 (CLV3). Among these, the CLE gene family has been extensively studied as key genes [6,7,8,9,10].
The CLE family of plant-specific genes is named after its founding CLV3/ESR gene that is specifically expressed in maize (Zea mays) [11,12,13]. Furthermore, Cock and McCormick discovered 39 related protein sequences associated with the CLV3/ESR family, which they named the CLE family. These proteins were characterized by conserved 12-residue domains essential for ensuring the function of C-terminal signal peptides and N-terminal hydrophobic signaling peptide [11]. The similarity of the remaining sequences is very low except for the conserved motif and secreted signal peptide [14]. Subsequently, CLE is explicitly described as a signal peptide that is cleaved from a longer pre-peptide with a similar structure: small proteins (usually fewer than 150 amino acids) consisting of an N-terminal signal peptide, followed by a variable domain with significant sequence diversity, and a conserved C-terminal CLE motif. These pre-peptides are translated and modified one or more times [11,14,15,16]. As for CLE gene family function studies, as early as the mid-1990s, most CLV1/CLV3 mutants were found to affect the meristem activity of plant stems, roots, and flowers [17,18]. Subsequently, the WUSCHEL (WUS)-CLV3 regulatory network was discovered, which controls the activity of the apical meristem at the stem tip [19]. In 2002, CLV3-CLV1/CLV2 was found as a receptor ligand in plants to signal from the stem cell population [20]. This marked the beginning of further studies on the CLE family. However, this is only applicable to the CLV1/CLV2/CLV3 genes. In 2006, CLV3/ESR1-LIKE 41 (CLE41) was shown to repress xylem differentiation in cell culture [21]. In a later study, similar to WUS and CLV3, CLE40 and WUS-RELATED HOMEOBOX 5(WOX5) were found to play a role in regulating the root meristem [22,23,24]. As the research on various genes of the CLE family has been continuously deepened, it has been discovered that CLE gene family have different functions to control the development of plants.
Based on domain structure and functional analyses, Whitford classified the peptide types of the CLE family into two categories: A (CLAVATA3 (CLV3)-like) and B (TRACHEARY ELEMENT DIFFERENTIATION INHIBITORY FACTOR (TDIF)-like) [25]. A-type CLE peptides promote cell differentiation in root and shoot apical meristems, whereas the B-type peptides CLE41–CLE44 do not promote. B-type CLE peptides suppress differentiation into tracheary elements. B-Type CLE peptides are mainly Tracheary Element Differentiation Inhibitory Factor TDIF-like [26,27]. The synergistic interaction of these two peptides inhibits differentiation and promotes auxin-mediated cell proliferation in the secondary meristem (vascular cambium), suggesting that specific CLE genes have dual functions and cell type-specific responses [28].
At present, the research on woody plants mainly focuses on the genetic transformation system [29], the molecular regulation related to wood formation [30,31,32], the molecular basis of forest economic traits [33,34], and the response control of plant stress resistance [35,36]. While the CLE gene family as a systemic regulatory hub governing stem cell dynamics across Arabidopsis thaliana tissues—particularly during organogenesis from embryogenesis to post-meristematic differentiation—the functional characterization of CLE networks in woody perennials remains critically understudied. The earliest research on CLE genes in forest trees was reported in Populus trichocarpa in 2016 [37]. Since then, research on the CLE gene family has focused on identifying the systematic classification of its family in woody plants or verifying a single function: for example, Camellia oleifera and some rosaceous plants, involving very few molecular regulatory networks [38,39]. Only Populus trichocarpa has been gradually studying the CLE family regulatory network during the development of vascular tissues, but the progress has been slow. Forest trees play a crucial role in water resource conservation, maintaining ecological balance, and providing medicinal compounds from their roots and leaves along with edible fruits, making them indispensable for both environmental sustainability and human well-being. For tall trees to grow healthily and vigorously, it is essential to maintain and properly differentiate the stem cells in the meristematic tissues. Here, we investigate the regulatory pathway of the CLE family by tracing the developmental sequence of plant organ formation, aiming to offer guidance for the growth and development of forest trees.
2. Development and Maintenance of CLEs in Root Apical Meristem
Root developmental plasticity is a critical determinant of plant fitness, enabling efficient acquisition of soil resources (water and nutrients) and systemic coordination of whole-plant growth. The establishment of RAM architecture begins with a stereotypical radial pattern at the root tip, where a small cohort of progenitor cells undergoes precisely oriented divisions to generate distinct tissue lineages [40]. Central to this process is the stem cell niche (SCN), a dynamic microdomain organized around the mitotically inactive quiescent center (QC) (Figure 1A). This niche contains the QC that is thought to be the initial cell that maintains the first surrounding cell in an undifferentiated state and gives rise to other stem cells; stem cells on the proximal (toward the shoot) side of the QC generate vasculature and pericycle; lateral stem cells of the QC give rise to endodermis, cortex, epidermis, lateral root cap; and distal columella stem cells (CSC) of the QC generate the protective cap of columella cells (CC) [41,42,43,44,45].
Unlike stems, root ecological niche restriction is mediated not by individual stem cells but by entire meristematic stem cell populations enveloped by the root cap [24,49]. CLE40 expression is localized to the basal region of the embryo during the globular embryo stage, where it initiates root meristem and vasculature formation. Post-germination, CLE40 is expressed in the CCs and localized at the distal end of QC [24,50]. Through the receptor-like kinase CLV1 and the CIKs (CLAVATA3 INSENSITIVE RECEPTOR KINASES)-assisted ARABIDOPSIS CRINKLY4 (ACR4) [51], CLE40 limits QC-derived signals whose activity or expression might depend on WOX5 function in the QC and act non-cell-autonomously to control CSC fate. CLE40 can positively regulate the promotion of WOX5 expression via the CLV2 receptor in the vascular initials [52]. In Solanum tuberosum, the homologous gene of CLV3 is StCLE4, which regulates stem cell activity and modulates both stem and root growth [53]. In lateral root apex meristem activity, CLV3 plays a central role in lateral root apical meristem activity. Under normal conditions, CLV3 is expressed in the pericycle of roots, and lateral root length is inversely correlated with CLV3 expression levels. However, CLV3 overexpression disrupts root tip meristem activity, leading to a short-root phenotype that is positively influenced by sucrose levels in the root [54]. Additionally, the CLV3-CLV2/SOL pathway regulates root meristem signaling, with SUPPRESSOR OF LLP1 2 (SOL2)/CORYNE (CRN) deficiency resulting in markedly reduced root length [55,56] (Figure 1A). Overexpression of CLE19 restricts root meristem cell size without directly affecting the QC or adjacent stem cells, instead acting on pericycle initiation cells via the CLV2 complex [57,58].
3. Development and Maintenance of CLEs in Shoot Apical Meristem
In forest trees, almost all above-ground tissues originate from the conserved dome-like SAM, which is actually a highly heterogeneous and highly organized structure controlled by stem cells [59,60]. Structurally, in monocotyledons like Oryza sativa L., SAM is organized into distinct layers: the L1 layer and L2 layer [61,62]. In dicotyledons, SAM forms between the two cotyledons and comprises three stem cell layers: the L1 layer, which generates the epidermis via anticlinal divisions; the L2 layer where cells undergo periclinal divisions in the meristem and produce mesophyll cells through vertical/peripheral divisions in leaf primordia; and the L3 layer, which differentiates into stem cell centers and vascular tissues via both anticlinal and periclinal divisions, as seen in Arabidopsis thaliana [59,63]. Functionally, stem cells organize the SAM into three domains: Central Zone, (CZ): including the organizing center (OC) which a central domain with slow division rates that maintains meristem integrity and supplies cells to the peripheral zone (PZ); the PZ, surrounding the OC, where rapid cell divisions generate organ primordia; the rib meristem (RM), located below the OC, which specifies central stem cell identity [64,65] (Figure 1B). Cells continuously proliferate, progressing through three cellular functional regions that are aimed to control different cells and, thus, regulate the differentiation, division, and formation of organ primordia and internal tissues [65].
CLV3 is localized to the overlying cell layers of the stem cell niche, where it regulates cell division and organogenesis, while WUS, which specifies stem cell identity and controls meristem activity, resides at the base of the stem tip [23,63,66]. STM, which maintains stem cell pluripotency, works synergistically with WUS and CLV3 to form a WUS-CLV3-STM regulatory loop that governs SAM homeostasis [67,68]. During the proliferation and differentiation of SAM stem cells, a Homeobox (HB) family transcription factor WUS and a Class I KNOX transcription factor STM can upregulate CLV3 expression by binding to the CLV3 promoter cis-acting elements (TAAT and TGACA), respectively [69,70]. Furthermore, STM assists WUS in forming WUS-STM heterodimers, which enhance WUS binding affinity to the CLV3 promoter via protein–protein interactions. This promotes CLV3 expression in the central zone (CZ), ensuring stem cell population stability [69,70,71]. The CLV3 gene encodes a 96-amino acid precursor protein that is post-translationally modified to yield a mature arabinosylated glycopeptide. This peptide contains a conserved 12- to 13-amino acid motif, with Leu and Arg identified as critical residues for restricting SAM size [11,66,72,73,74]. Spatiotemporally, CLV3 expression is confined to the epidermal and subepidermal layers of the CZ in shoot and floral meristems but is absent in the RM [66,67,70] (Figure 1C).
When CLV3 promotes cell proliferation, elevated WUS levels recruit HAIRY MERISRTEM1/2 (HAM1 and HAM2 (members of the GRAS transcription factor-encoding HAM family)). These WUS-HAM heterodimers suppress CLV3 expression, thereby establishing apical polarity of the CLV3 expression domain along the SAM axis to regulate stem cell homeostasis [46,75,76,77,78,79]. In embryonic development, CLV3 expression is regulated exclusively by WUS, independent of STM [80]. However, during later developmental stages, STM and WUS jointly modulate CLV3 levels, with CLV3 responsiveness to WUS confined to the apical meristem. Sustained overexpression of WUS triggers exocytosis-dependent CLV3 signaling, which coordinates four distinct cellular pathways to repress WUS in the RM, forming a negative feedback loop [20,81]. Mechanistically, CLV3 inhibits WUS primarily via LEUCINE-RICH REPEAT RECEPTOR-LIKE KINASES (LRR-RLKs) and CIKs. These receptors act synergistically, where CIKs enhance LRR-RLK activity to amplify downstream signaling cascades that suppress WUS expression [74,82,83] (Figure 1C).
3.1. CLV3-CLV1
As a ligand–receptor pair, CLV3 undergoes proteolytic cleavage and directly binds to CLV1, an LRR-RLK, with a dissociation constant (Kd) of 17.5 nM. This interaction triggers CLV1 endocytosis to regulate its membrane trafficking [84,85,86]. The binding ability between CLV3 and CLV1 is mainly affected by the arabinosylation of CLV3 and the affinity of different amino acids in the extracellular domain of CLV1 [84,86,87,88] (Figure 1C).
3.2. CLV3-CLV2-CRN/SOL2
Both CLV2, a LEUCINE-RICH REPEAT (LRR, protein lacking a kinase domain) and SUPPRESSOR OF LLP1 2 (SOL2)/CORYNE (CRN) (a transmembrane pseudokinase devoid of LRRs) are synthesized on the endoplasmic reticulum (ER). The transmembrane (TM) domain of CRN binds specifically to CLV2, enabling the CRN-CLV2 complex to localize to the plasma membrane (PM). This interaction neutralizes an acidic inhibitory motif in the extracellular region of CLV2, which is essential for PM trafficking. Notably, CRN does not enhance CLV2 accumulation at the PM but facilitates its targeting. The mature CRN-CLV2 complex subsequently binds CLV3 to mediate signaling [84,89,90,91]. CLV2-CRN is parallel to CLV1 and co-responds to CLV3 signal transduction [92,93] (Figure 1C).
3.3. CLV3-RPK2
As a member of the RLKS family of receptor-like kinases, (RECEPTOR-LIKE PROTEIN KINASE2 (RPK2))/TOAD2 regulates the development of anther microspores and tapetum, and mutations in RPK2 cause anther breakage [94,95]. In CLV3-null backgrounds, RPK2 mutants exhibit reduced SAM size and increased carpel number, indicating that RPK2 participates in CLV3-dependent signaling within the SAM to repress WUS expression. While RPK2 does not directly bind CLV3 via its leucine-rich repeat (LRR) domain, the mechanism of interaction remains unclear [96]. We hypothesize that RPK2 participates in the CLV3 pathway not solely as a ligand but may act through alternative mechanisms in plant signaling (Figure 1C).
3.4. CLV3-BAMs
BARELY ANY MERISTEM(BAM) is one of the leucine-rich repeat receptor-like kinases (LRR-RLK). Constitutive expression of BAM1/BAM2 partially rescues the CLV1 mutant phenotype, confirming their functional homology with CLV1. Unlike CLV1, BAM1/BAM2 exhibit broader expression patterns [97]. Photoaffinity labeling assays demonstrate direct binding between the BAM1 ectodomain and CLV3 peptide. While single mutants (BAM1, BAM2, or BAM3) show no obvious developmental defects, double (BAM1/BAM2) and triple (BAM1/BAM2/BAM3) mutants exhibit reduced SAM size due to stem cell depletion [96,98] (Figure 1C). Thus, the CLV3-BAMs regulatory pathway was identified.
Current understanding of CLV3 downstream signaling—particularly phosphorylation cascades involving kinases and phosphatases—remains limited. CLV3 activates a phosphorylation cascade mediated by MPK3/MPK6, which partially rescues the CLV1 mutant phenotype, implicating these kinases in dependent signaling CLV1 [99]. The kinase-associated protein phosphatase KAPP directly interacts with CLV1, dephosphorylating it to attenuate CLV1 activity. Additionally, the PP2C-type phosphatases POLTERGEIST (POL) and POL-LIKE1 (PLL1) act as negative regulators downstream of CLV1/BAM receptors, modulating WUS expression to control apical stem cell dynamics [83,100] (Figure 1C). Genetic evidence shows that CLV3-CLV1, CLV3-CLV2/SOR function independent of each other. However, studies on pathway crosstalk show that the first two pathways may be connected these pathways may converge-potentially compensating for each other to form a regulatory network that maintains stem cell homeostasis when one pathway is dysregulated [82].
4. Development and Maintenance of CLEs in Stem and Root Cambium
Stem and root apical meristem cells continue to divide and differentiate to form the procambium that have tissue with permanent meristematic activity. Procambium serves as the primary source of xylem and phloem cells, while also contributing to the structural framework of plant stems and roots [101].
In the stem, shoot apical localized procambium (PC) initials are described as the primary meristem that differentiates basally to produce primary vascular bundles that daughter cells of PC differentiate into protophloem (PPh) toward the outside of the stem and protoxylem (PXy) toward the inside of the stem. Moving basally toward developmentally older tissues, actively dividing meristematic cells within vascular bundles were described as metacambium (MC) that subsequently divide into the secondary vascular cambium meristematic cells that produce secondary phloem and secondary xylem [102,103,104]. In this review, we classify stem tissues into procambium, xylem, and phloem to elucidate CLE family-mediated regulatory mechanisms. Within the procambium, the TDIF predominantly regulates cambial activity [25]. In Populus trichocarpa, MYB31 located in the cambium layer regulates the PtCLE41/PtCLE42/PtCLE44 peptides produced by the phloem to translocate the cambium and through the TDIF RECEPTOR (TDR)/PHLOEM INTERCALATED WITH XYLEM (PXY) membrane protein kinase signaling pathway, the PtCLE41p/PtCLE42p/PtCLE44p combine with WOX4/WOX14 to promote procambial cell proliferation while suppressing xylem cell differentiation [105,106]. As a downstream transcription factor of TDIF-PXY, GLYCOGEN SYNTHASE KINASE 3 PROTEINS (GSK3s) inhibit BRI1-EMS SUPPRESSOR 1 (BES1), thereby inhibiting cambium-to-xylem cell differentiation [107]. In Populus trichocarpa, MYB31 could either promote cell proliferation through restraining the MYB31-MYB72-WOX4 module or inhibit cambial activity through restraining the MYB31-MYB72-VASCULAR CAMBIUM-RELATED MADS 2 (VCM2)/PIN-FORMED 5 (PIN5) modules (VCM2/PIN5) [108,109]. In gymnosperms, CLE41/CLE44 play a role not only in the phloem but also in the tracheary elements (TEs) [105,106]. PtCLE47 and PtCLE20, two poplar CLE polypeptides, respectively, promote and inhibit procambial cell proliferation [110,111] (Figure 1D).
In the root, the procambium can generate primary xylem and primary phloem which includes sieve elements (SEs), companion cells (CCs) and related cell types. The pericycle generates lateral roots and initiates vascular cambium (responsible for secondary phloem and xylem production) [47,112]. In xylem precursor cells, the receptor-like kinases BAM1, BAM2, and BAM3 collectively function as major receptors for CLE9/CLE10 peptides, negatively regulating periclinal cell division to control xylem file numbers [21,113] (Figure 1D). During protophloem development, CLE33 critically modulates BAM1/BAM2/BAM3 and CLV2/CRN complexes to regulate SE differentiation [114]. In the protophloem, CLE25/CLE26 are expressed early in the SE cells lineage and promote the initiation and development of phloem through the complex interaction with CLE-RESISTANT RECEPTOR KINASE-CLV2 (CLERK-CLV2) receptor to control the SE precursor cell (SPC) receptor-like protein [41,115,116]. A suppressor screen of BREVIS RADIX (BRX) mutants identified the CLE45-BAM3 axis as a compensatory pathway for SE differentiation [117]. MEMBRANE-ASSOCIATED KINASE REGULATOR 5 (MAKR5) acts as a post-transcriptionally regulated amplifier of the CLE45p signal that acts downstream of BAM3 [118,119]. However, this way of signaling antagonizes BAM1/BAM2-mediated CLE11/CLE12/CLE13 signaling in the phloem initials [119]. Additionally, phloem-Dofs not only enforce SE and CC formation but also activate the production of CLE25, CLE26, and CLE45 that reduce the level of phloem-Dofs by interacting with BAMs/CIKs, thereby inhibiting the excessive production of SEs and CCs [120]. Furthermore, CLE peptides (CLE1/CLE3/CLE4/CLE7) modulate lateral root growth and branching through the CLE-CLV1 signaling module in response to nitrogen availability, without affecting primary root development [121]. Collectively, these pathways fine-tune root architecture and elongation (Figure 1D).
5. Development and Maintenance of CLEs in Leaf
Leaf initiation and proper spatial orientation are essential for efficient photosynthesis, thereby ensuring plant survival. Within the SAM, the CZ harbors stem cells, while organogenesis initiates in the PZ [122]. During vegetative SAM development, CZ-derived stem cells undergo continuous division, with daughter cells migrating laterally into the PZ to form leaf primordia structures that are small and regularly spaced [123]. Cells in the PZ region divide rapidly and continuously, forming leaf protodermal cells, which can either directly divide into pavement cells (general epidermal cells) or become meristemoid mother cells (MMCs) that are stomatal lineage stem cells [124,125]. Following primordium initiation, leaves develop along three distinct polarity axes: axial-dorsal, proximal-distal, and central-lateral [126,127] (Figure 1E).
Auxin determines the fate of organ primordia in the peripheral region of PZ, and the formation of leaf primordia is dependent on the auxin maximum formed by the polar auxin transport mediated by the PIN-FORMED 1 (PIN1) gene [128,129]. Belonging to AUXIN RESPONSE FACTORs (ARFs), ARF5 (Mp) shows threshold expression in PZ to CZ and mediates auxin signaling by negatively regulating CLV3 by repressing ENHANCER OF SHOOT REGENERATION1/DORNROSCHEN (ESR1/DRN) that can combine BRAHMA(BRM) and WUS to form a ternary protein complex [80,130,131,132,133,134,135,136]. This mode can prevent the axillary meristem (AM) disturbance caused by high expression of CLV3. In Oryza sativa, NDL1 is the ortholog of Arabidopsis thaliana of ESR1/DRN and autonomously regulates leaf development [137]. This suggests that CLV3 affects the development of leaf initially, and CLE5/CLE6 are positively regulated by BLADE-ON-PETIOLE1/2 (BOP1/BOP2) at the petiole base so that their loss of function makes the petiole slightly wider. The transcription of CLE5/CLE6 is negatively regulated by ASYMMETRIC LEAVES2 (AS2) at the distal positions of petioles and leaves. But CLE5/CLE6 have little effect on the leaf. Referring to the CLE-WOX pathway in SAM, it was found that the expression of CLE5/CLE6 in leaves is also positively regulated by the WOX transcription factors, PRESSED FLOWER (PRS) and WOX1, which promote leaf growth and increase leaf margin cell-files [138]. In MCCs, CLE9/CLE10 bind to HAESA-LIKE1 receptor kinase (HSL1) to phosphorylate SPCH through a MAPK cascade to negatively control epidermal division [113]. By suppressing THE ENZYME 1-AMINOCYCLOPROPANE-1-CARBOXYLIC ACID SYNTHASE (ACS), CLE42 accumulates ETHYLENE-INSENSITIVE3 (EIN3)-binding F-BOX1/2 (EBF1/EBF2) protein, which degrades EIN3 (a master transcription factor in the ethylene pathway), a key component of the ethylene signaling pathway, through the ubiquitin–proteasome pathway, thereby delaying leaf senescence [139]. Additionally, CLE14 regulates age-dependent and stress-induced leaf senescence through promoting the expression of the JUB1-ROS scavenging gene (CAT3, APX1, APX3) to mediate ROS scavenging [139,140,141] (Figure 1E).
6. Development and Maintenance of CLEs in Floral Meristem
When the plant’s internal organs mature under a favorable external environment, SAM receives the flowering signal and transforms into the inflorescence meristem (IM), which marks the transition from vegetative growth to reproductive growth and then the formation of young flower primordia [142]. The young floral primordia retain apical stem cells that undergo lateral divisions within the IM, generating FMs. Each FM orchestrates the sequential development of floral organ whorls (sepals, petals, stamens, and carpels) to form a complete flower [143,144]. Floral organogenesis proceeds through a temporally and spatially regulated sequence, with partially overlapping phases ensuring precise whorl patterning [145]. Therefore, FMs are continuously produced by multiple developing organs, and unlike the SAM, which maintains expansive growth zones, FM activity occurs within spatially confined regions separated by narrow developmental boundaries [146].
CLV2 is expressed in IM, and CRN is expressed in the early flower primordium and even expresses in the whole flower primordia. The CLV2/CRN receptor complex promotes the growth and development of flower primordia [147]. Mutations in the CLV2 site lead to enlargement of stem and flower meristem and developmental defects in pistil, petals, and stamens [148]. CLV1 and CLV3 are expressed in the center and apex of FM. Compared with STM, CLV1/CLV3 has the same expression pattern but opposite function; that is, STM mutants fail to form undifferentiated cells in stem meristem during meristem development while CLV1/CLV3 mutants accumulate excessive undifferentiated cells in flower meristems, causing over proliferation of central floral tissues [17,63]. STM and KNAT-6 mutations have additive effects in regulating CLV3 inflorescence size [19,149]. In maize, the THICK TASSEL DWARF (TD1) and FASCIATED EAR2 (FEA2) genes encode CLV1-like LRR receptor kinase and the CLV2-like LRR receptor protein, respectively [150]. The TD1 and FEA2 double mutant exhibited a phenotype with an increased inflorescence size [151]. In Oryza sativa L, FON1 encodes a gene homologous to CLV1 and maize TD1, while FON2 encodes a CLE protein associated with AtCLV2. In FON1 and FON2 mutants, FMs are increased, resulting in an increased number of flower organs such as stamens and carpels [152]. This suggests that CLV1/CLV2/CLV3 genes affect flower development. Notably, WUS is not involved in floral meristem development and the CLV3-CLV1 regulatory pathway.
If the external environment temperature changes, CLE gene family, combined with auxin, play an irreplaceable role in responding to changes in flower primordia development [83]. At normal temperatures, the CLV3 pathway, like the thermal sensing ELF3 factor containing the Poly-Q structure, is functionally degraded by being sequestered by the YUCCA (YUC) complex [153,154,155]. Under lower temperatures, receptor complexes CLV1 and CLV2/CRN transduce the CLV3/CLE25 signal to promote normal flowering in plants by upregulating YUC-dependent auxin biosynthesis [48,156,157]. Although CLE25 is inhibited by CLV3, in the case of CLV3 mutation, CLV3 promoter can bind to CLE25 to compensate for the flower phenotype [157,158]. This suggests that the CLV3 promoter pathway is shown to be important in regulating the transition state of flower primordia during vegetative-to-reproductive growth, though the intermediate pathways and associated genes remain uncharacterized. Under high temperatures, ELF3 upregulates auxin to control flower development [48,155]. Therefore, the significance of temperature in regulating the CLE channel has also been given due attention [159] (Figure 1F).
7. Compensation Mechanism of CLEs in Plant
The compensation mechanism provides fault tolerance for plant development, enabling the maximization of growth along normal developmental trajectories [160,161,162]. Due to lineage-specific factors, the number, functional relationships, homologous retention, and diversity variation (including redundancy) of inbred family members differ significantly among distantly related species. However, the CLE protein family demonstrates remarkable structural conservation—particularly in the C-terminal CLE motif, which is critical for receptor binding [163]. In Arabidopsis thaliana, following CLV3 deletion, the CLE16 and CLE17 signaling pathways actively regulate WUS, limiting stem and floral stem cell accumulation and buffering infinite apical enlargement caused by CLV3 loss. These pathways are not sensed by CLV1 or CLV2 but exclusively by the BAM1/BAM2 receptor kinases, indicating their role as compensatory mechanisms for CLV3 deficiency [97,164]. In CLV1 mutants, ectopic BAM expression in the RM partially compensates for CLV1 loss [96,165,166]. Additionally, other CLE peptides may exhibit functional redundancy during SAM maintenance. This is evidenced by complete or partial CLV3 complementation when CLE1, CLE6, CLE9, CLE11, CLE12, CLE13, CLE19, CLE21, or CLE22 are expressed under the CLV3 promoter [47,167]. Notably, single and double mutants of CLE16, CLE17, and CLE27 show no detectable phenotypes in the SAM or IM.
In Solanum lycopersicum, SlCLE compensation is functionally active, with SlCLE9 partially restoring SlCLV3 stem cell homeostasis primarily via SlCLV1 [158]. However, in Arabidopsis thaliana, the CLE9-CLV1 regulatory pathway remains poorly characterized. CLE40, encoding a putatively secreted protein with functional similarity to CLV3, can fully substitute for CLV3 in the SAM. The CLV3 promoter drives CLE40 expression to compensate for CLV3 deficiency [47]. In Zea mays L., ZmFCP1 and ZmCLE1E5 partially rescue the enlarged inflorescence meristem phenotype caused by ZmCLE7 mutations [168].
8. Conclusions
When we review the research process of the CLE family, it is not difficult to find that although the CLE family has continuously evolved over millions of years, and in addition to parasitic nematodes, the CLE family is found in plants and is one of the largest families of expanded plant polypeptides [16,169] (Figure 2A). However, reports-of-CLE-in-non-pattern. woody plants are very limited, mainly for the following reasons.
As non-flowering woody plants continue to evolve, it remains unclear whether the CLE family have evolved into more refined branches, which seriously affects the search for CLE family homologous genes in trees. Here, we conduct extensive studies on mosses (such as P. patens [163]), ferns (such as S. moellendorffii), gymnosperms (such as P. abies), and angiosperms (such as O. sativa [170], Arabidopsis [11], P. trichocarpa [37] S. purpurea, P. deltoides, P. persica, A. trichopoda [171] (Figure 2B). We found that the CLE family between lower plants and higher plants has changed significantly, and more complex and precise branches have been differentiated. However, the amino acid structure of CLE was still conserved (Figure 2A). Therefore, we can use the research methods in Arabidopsis thaliana, such as molecular probes and gene editing, to locate the CLE gene family in forest trees.
The tissue positioning of the CLE family during plant development in woody plants is currently in a very unclear state. Most woody plants are also limited to only one of the last few genes in the CLE family and are researchers unable to form an overall network structure. Our analysis revealed overlapping expression profiles of CLE gene family in diverse meristematic tissues, such as CLV1 being expressed in roots, stems, and cambium (Figure 3). CLV1/CLV2/CRN functions as a signaling transduction component extensively involved in plant organ developmental processes. This study shows that receptor kinases exhibit multi-organ distribution characteristics. The existence pattern of receptor kinase is highly conserved. It is very likely that the CLE gene family in woody plants are also located in the tissues of woody plants. For specifically expressed genes like CLV3, their regulatory pathways demonstrate shared features in root and shoot tissues (Figure 3). Based on evolutionary conservation analysis of the CLE family, CLV3 orthologous genes likely exert regulatory roles in both shoot and root apical meristems of woody plants. Comparative analysis of root and shoot procambium tissues revealed no functional redundancy among CLE family members such as CLE25 and PtCLE20 (Figure 3). These findings indicate that the CLE family exhibits both specific single-gene expression patterns and potentially unidentified functionally related genes. This suggests that CLE peptides can function both as individual initiation signals and as signaling molecules that coordinate with other genes to regulate plant development. This unique genetic function provides a very high degree of accuracy for the research on the cambium of woody plants.
The trees passed down from generation to generation are large in size and come in various shapes. The developmental regulatory networks existing in plant organs and tissues are far more complex than those in some model plants. Although Arabidopsis thaliana confers limited translational applicability for arboreal species, developmental genetic analyses of meristem regulatory networks have established that a phloem-specific CLE41-PXY/TDR-WOX4 regulatory circuit in Populus trichocarpa is discovered based on the WUS-CLV3 ligand–receptor module that is an evolutionarily conserved regulatory module (Figure 1). This suggests that the reference Arabidopsis regulatory network is crucial for elucidating the CLE family foundation in forest trees. This necessitates synergistic integration of pan-omics analyses (spatiotemporal proteomics, phospho-signaling mapping) with CRISPR-Cas9-mediated tissue-specific CLE knockout systems to resolve the mechanistic coupling between peptide ligand gradients and xylary differentiation trajectories in woody perennials. For floral and foliar organs in forest plants, auxin regulation could serve as a key entry point to elucidate the CLE signal transduction network in forest trees. Furthermore, it is imperative to integrate additional biological experiments to advance this research and address existing challenges for ultimately enhancing wood yield.
The CLE family involves more sophisticated compensation mechanisms during plant development. Not only for trees, but for all plants, there is even more profound room for the study of this compensation mechanism. The investigation into the compensation mechanism of the CLE gene family has revealed functional complementarity among its members. This signaling compensation fundamentally underpins meristem homeostasis, wherein developmental robustness is achieved via multilayered feedback control rather than isolated genetic components. Given this systems-level complexity, reductionist approaches focusing on single-gene characterization fail to capture the gene function. Therefore, it is imperative to explore diverse methodologies for a more scientific and comprehensive understanding of genes involved in forest tree development, such as protein interactome mapping, genome sequencing, and so on. The mechanistic insights derived from such multidimensional analyses hold significant potential for optimizing genome-informed silvicultural practices aimed at enhancing carbon sequestration efficiency and ecosystem service provisioning in managed forest stands.
In summary, as members of the polypeptide family, the CLE family genes act as signal regulators in meristems, maintaining the balance and transformation of stem cell homeostasis and thereby exerting regulatory effects on plant growth and development. Although extensive research has been conducted on model plants such as Arabidopsis thaliana and herbaceous plants, research on large, long-lived trees that play a key role in climate regulation and ecological balance is still limited. Therefore, it is more important to expand the research on how the CLE family regulates the meristems of forest trees.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Khavinson V. Linkova N. Diatlova A. Dudkov A. Peptide regulation of plant cells differentiation and growth BIO Web Conf.2024820200310.1051/bioconf/20248202003 · doi ↗
- 2Baskin C.C. Baskin J.M. The rudimentary embryo: An early angiosperm invention that contributed to their dominance over gymnosperms Seed Sci. Res.202333637410.1017/S 0960258523000168 · doi ↗
- 3Kathryn Barton M. Cell type specification and self renewal in the vegetative shoot apical meristem Curr. Opin. Plant Biol.19981374210.1016/S 1369-5266(98)80125-810066561 · doi ↗ · pubmed ↗
- 4Ali S. Khan N. Xie L. Molecular and hormonal regulation of leaf morphogenesis in Arabidopsis Int. J. Mol. Sci.202021513210.3390/ijms 2114513232698541 PMC 7404056 · doi ↗ · pubmed ↗
- 5Dinneny J.R. Benfey P.N. Plant stem cell niches: Standing the test of time Cell 200813255355710.1016/j.cell.2008.02.00118295573 · doi ↗ · pubmed ↗
- 6Scofield S. Murray J.A.H. KNOX gene function in plant stem cell niches Plant Mol. Biol.20066092994610.1007/s 11103-005-4478-y 16724262 · doi ↗ · pubmed ↗
- 7Scofield S. Dewitte W. Murray J.A. STM sustains stem cell function in the Arabidopsis shoot apical meristem and controls KNOX gene expression independently of the transcriptional repressor AS 1Plant Signal. Behav.20149 e 2893410.4161/psb.2893424776954 PMC 4091562 · doi ↗ · pubmed ↗
- 8Kwon C.S. Chen C. Wagner D. WUSCHEL is a primary target for transcriptional regulation by SPLAYED in dynamic control of stem cell fate in Arabidopsis Genes Dev.200519992100310.1101/gad.127630515833920 PMC 1080137 · doi ↗ · pubmed ↗