Impact of Temperature on the Biochemical Potential of Five Newly Isolated Strains of Microalgae Cultured in a Stirred Tank Reactor
Panagiotis Dritsas, George Aggelis

TL;DR
This study examines how temperature affects the growth and biochemical properties of five new microalgae strains, highlighting their potential as aquaculture feed additives.
Contribution
The novelty lies in the isolation and characterization of five new microalgae strains and their temperature-dependent biochemical potential.
Findings
VAS2.5, VON5.3, and PAT2.7 showed high biomass production at both 25 °C and 20 °C.
Temperature significantly influenced lipid composition, with glycolipids and sphingolipids dominating at 25 °C and neutral lipids at 20 °C.
Most strains exhibited high protein content, making them suitable for aquaculture feed.
Abstract
The microalgal strains Picochlorum costavermella VAS2.5, Picochlorum oklahomense SAG4.4, Picochlorum oklahomense PAT3.2B, Microchloropsis gaditana VON5.3, and Nephroselmis pyriformis PAT2.7 were cultured in a Stirred Tank Reactor at 25 °C or 20 °C in modified artificial seawater and their biotechnological potential was assessed. VAS2.5, VON5.3, and PAT2.7 were high in biomass production at both temperatures (i.e., 438.8–671.3 mg/L and 418.4–546.7 mg/L at 25 °C and 20 °C, respectively), though P. oklahomense strains grew only at 25 °C. The highest lipid percentage was recorded for the cultures of VAS2.5 (19.3 ± 0.7%) and VON5.3 (16.4 ± 1.5%) at 25 °C, notably rich in Δ5,8,11,14,17C20:5, while PAT2.7 proved a major producer of Δ9C16:1. The predominant lipid fraction was glycolipids and sphingolipids (41.3–57.4%) for VAS2.5, PAT2.7 at 25 °C and VON5.3 at 20 °C and neutral lipids…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
 Figure 1
Figure 1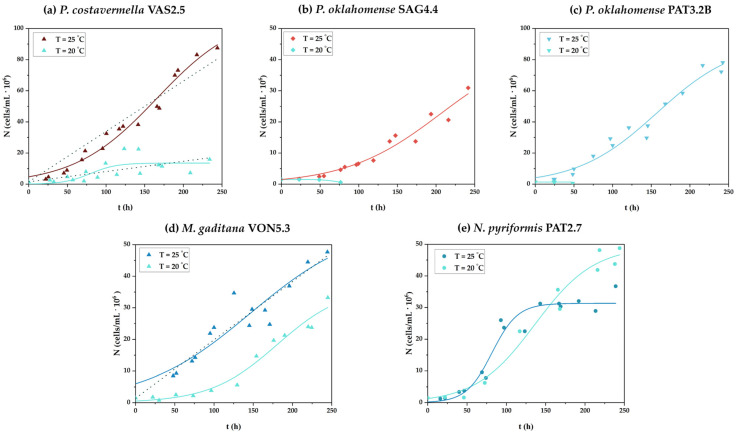 Figure 2
Figure 2- —Isolation of microalgae native of the Ionian Sea and their use in the production of high added value products IonianAlgae
- —Greek State (Greek General Secretariat for Research and Technology)
- —European Union
- —Hellenic Foundation for Research and Innovation (HFRI)
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsAlgal biology and biofuel production · Aquatic Ecosystems and Phytoplankton Dynamics · Marine and coastal ecosystems
1. Introduction
Microalgae are photosynthetic microorganisms with a global distribution, found not only in all types of water systems but in terrestrial ecosystems and the atmosphere as well [1,2,3]. Besides their obvious high ecological value, microalgae, through the conversion of solar energy and CO_2_, biosynthesize cellular materials of high nutritional interest (e.g., lipids, proteins, carbohydrates, pigments, etc.) for aquatic and terrestrial organisms in significant quantities [4,5]. Typically, green microalgae, rather than other groups of photosynthetic microorganisms, are considered as a superior oil source compared to conventional plants for a number of reasons. The major ones are related to their higher growth rates and their ability to grow under a wider range of pH and temperature values and valorize CO_2_ in a more efficient way comparing to plants [6,7,8].
More importantly, some microalgal species, e.g., representatives of the genera Picochlorum and Microchloropsis, are of additional interest due to their ability to synthesize polyunsaturated fatty acids (PUFAs), such as the omega-3 fatty acids α-linolenic (^Δ9,12,15^C18:3, ALA), eicosapentaenoic (^Δ5,8,11,14,17^C20:5, EPA), and docosahexaenoic (^Δ4,7,10,13,16,19^C22:6, DHA). Such fatty acids are sought after by the food, feed, and pharmaceutical industries because of their proven beneficial effects against cardiovascular and neurodegenerative diseases, various forms of cancer, etc. [9,10,11,12]. On the other hand, microalgal lipids that contain more saturated fatty acids, like those that belong to the Nephroselmis genus, are used as feedstock for biodiesel production, because they are resistant to oxidation and yield more energy during combustion [13,14,15].
In general, the major advantage of microalgal oils against the fish oils, which are the main source of omega-3 fatty acids, is that they are free of undesirable odors and pollutants such as heavy metals and microplastics [16,17]. Additionally, the extensive use of microalgal oils will contribute decisively to the context of controlling and reducing intensive fishing globally. After all, microalgae farming incurs low production costs and, thus, can offer economic benefits by reducing dependency on imported fish oil in countries of need [18,19]. Yet even though >50,000 species of microalgae are described, only a few are cultivated on a large scale due to their high production costs [20,21,22].
Therefore, the need to use new strains with high efficiency rates when used in large-scale applications is of outmost importance. However, a crucial challenge that is associated with the yield of lipid production is the design of proper culture conditions. Environmental factors, such as temperature, irradiance, and nutrient availability majorly affect lipid content and fatty acid composition in microalgae cells [23,24,25]. For instance, at low temperatures, one might expect increased synthesis of unsaturated fatty acids, including PUFAs, as cells need to maintain the fluidity of their membranes, increasing their PUFA content. Evidently, in low temperatures, higher yields of these molecules of high biotechnological significance can be achieved [26,27].
The aim of the present study was to investigate the effect of temperature on the physiology and high added-value metabolite synthesis capability of five strains of microalgae, previously isolated and characterized with molecular techniques [13], that belong to genera Picochlorum, Microchloropsis, and Nephroselmis, due to the attractive features of representatives of the aforementioned genera and the relatively limited available literature for some of these strains [13,28,29,30,31,32], when cultured in a Stirred Tank Reactor (STR). One of the main reasons for using these strains to serve the cultures in the current investigation relates to the relatively limited literature available on most of these strains and the lack of data on the potential utilization of such microorganisms on a commercial scale. Even though the main focus was on lipid production, the fatty acid composition of total lipids and their lipid fractions, and the content of these strains in proteins, polysaccharides, and pigments were also analyzed. As a result of the aforementioned analyses, the biotechnological potential of the used strains when cultured in the STR was assessed.
2. Materials and Methods
2.1. Biological Material and Culture Conditions
The microalgal strains Picochlorum costavermella VAS2.5, Picochlorum oklahomense SAG4.4, Picochlorum oklahomense PAT3.2B, Microchloropsis (Nannochloropsis) gaditana VON5.3, and Nephroselmis pyriformis PAT2.7, which were previously isolated from coastal areas of the Ionian Sea of Greece and identified by the use of molecular techniques [13], were used as biological material.
The strains were maintained in 0.25 L Erlenmeyer flasks containing 0.05 L of modified Artificial Sea Water (mASW) and were regularly sub-cultured. The composition of mASW is presented in Table 1. The flasks containing mASW were sterilized at 121 °C for 20 min, inoculated with 10^5^ cells/mL of a fresh inoculum, derived from a 7-day pre-culture grown in sterilized mASW, and incubated at T = 25 ± 1 °C. Illumination was constant with an intensity of 300 μmol/m^2^∙s provided by linear fluorescent day light tubes (T5, 8 W, 6500 K). Before sterilization, the pH of the growth medium was adjusted to 8.5 ± 0.5 by adding 2.5 M NaOH (Sigma-Aldrich) solution and using an Orion 420A digital pHmeter (Thermo Scientific, Osterode, Germany) equipped with a glass electrode (HANNA Instruments, Athens, Greece). The pH of mASW was also measured after sterilization.
The microalgal strains were cultured in monoalgal but non-axenic batch cultures for approximately 250 h in a closed system bioreactor, type Ralf Plus-System from Bioengineering (Wald, Switzerland), which is a Stirred Tank Reactor (STR), with a capacity of 3.7 L and a working volume (V_w_) = 2 L. The STR was converted into a light bioreactor after 8 fluorescent lamps (T5, 8 W, 6500 K) were placed circumferentially to the culture vessel, providing continuous illumination of 1071 μmol/m^2^∙s intensity (Figure 1). Prior to culture initiation, the culture vessel was cleaned with 70% ethanol (vol/vol), approximately 1.8 L of sterile mASW was added, and inoculation was performed with approximately 0.2 L of pre-cultured microalgae so that the initial cell density was 1.5 ∙ 10^6^ cells/mL. The cultures were maintained at T = 25 ± 1 °C or T = 20 ± 1 °C under 100 rpm agitation and natural air was supplied at a rate of 0.5 vvm, which was passed through a Whatman-type bacteriological filter with a pore size of 0.2 μm. Both the selected temperatures correspond to the sea temperatures from which the original samples were collected. The initial pH was 8.5 ± 0.3 and was maintained at this value by the addition of 2.5 M HCl (Sigma-Aldrich) automatically added to the medium. At the end of each culture, the biomass was collected for quantification and further chemical analyses. Cultures were carried out in duplicate. The monitoring of the culture conditions and the physicochemical parameters of the system was possible through a computer equipped with Wonderware InTouch 9.5.001 software (Bioengineering, Wald, Switzerland).
2.2. Cell Growth and Biomass Determination
Daily cell counting in a Neubauer improved cell counting chamber (Poly-Optik, Bad Blankenburg, Germany) was performed, similar to other studies [33,34], so as to assess microalgae growth. In brief, 1 mL of microalgae culture was subtracted from the culture vessel daily, diluted properly, and in each slot of the Neubauer chamber, 10 μL of microalgae sample was placed. Then, under an optical microscope Carl Zeiss Axiostar (GmBH, Gottingen, Germany), at total magnification of 400×, cell density was calculated following the equation provided below (Equation (1)):
where N_av._ per square is the average number of cells per square of the Neubauer chamber and DF is the dilution factor.
The cell counts were expressed as microalgal cell density (cells/mL). In addition, the integrated version of Verhulst’s model (i.e., Sigmoidal Logistic function) (Equation (2)) was applied in order to estimate the parameters of growth:
where N represents the number of cells/mL∙10^6^ at time t, b is a positive constant equal to (N_max_ − N0)/N0 (where N0 is the initial number of cells/mL∙10^6^), μ is the specific growth rate (1/d), and N_max_ is the carrying capacity of the system. The parameter values were determined by fitting Equation (1) to the experimental data, using the Levenberg–Marquardt method for parameter value optimization. The minimization of the residual root means square error between the experimental and model-predicted data, quantified by the values of the coefficient of determination R^2^, was used as a criterion for parameter optimization.
Biomass determination was held by gravimetric determination of dry biomass. Specifically, prior to the gravimetric determination, harvesting of the microalgal cells by centrifugation at 7455× g for 10 min at 4 °C (NÜVE NF 800R, Ankara, Turkey) occurred. Then, washing of the cells with deionized water and centrifugation under the aforementioned conditions occurred in duplicate and then the biomass was collected and dried at 80 °C until constant weight and expressed as g/L.
2.3. Lipid Extraction and Purification
Total lipids from the microalgal cells were extracted in chloroform (PENTA)/methanol (Fisher Chemical, Hampton, NY, USA) 2:1 vol/vol, in accordance with a modified version of the Folch et al. (1957) [35] method [36]. Then, the extracts were filtered through Whatman No. 1 paper, washed with a 0.88% (wt/vol) KCl (Merck) solution in order to remove non-lipid components (i.e., lipoproteins, pigments), and dried over anhydrous Na_2_SO_4_ (Sigma-Aldrich). Consequently, solvent evaporation under vacuum, using a Rotavapor R-210 evaporator (BUCHI, Flawil, Switzerland), occurred and the total cellular lipids were gravimetrically determined and expressed as the percentage of lipids in the dry biomass (L/x%, wt/wt).
2.4. Lipid Fractionation
A column (25 × 100 mm) containing 1 g silicic acid (Fluka), which was activated by heating at 80 °C overnight, was used for the fractionation of approximately 100 mg of microalgal lipids which were priorly dissolved in 1 mL chloroform. The column was washed successively with 100 mL dichloromethane (Sigma-Aldrich) to obtain neutral lipids (N), 100 mL acetone (Fluka) to obtain glycolipids plus sphingolipids (G + S), and 50 mL methanol (Sigma-Aldrich) to obtain phospholipids (P) [33]. Following the evaporation under vacuum of the respective solvent of each fraction, all fractions (N, G + S, and P) were quantified gravimetrically and expressed as a percentage of total lipids.
2.5. Fatty Acid Composition of Cellular Lipids
The identification of total lipids and their lipid fraction composition in fatty acids was made attainable after their conversion into their fatty acid methyl-esters (FAMEs) and analysis with Gas Chromatography (GC). Specifically, an Agilent 7890A GC device (Agilent Technologies, Shanghai, China) equipped with a flame ionization detector at 280 °C and an HP-88 (J&W Scientific, Folsom, CA, USA) column (60 m × 0.32 mm) was used. The carrier gas was helium, at a flow rate 1 mL/min, and the analysis was run at 200 °C. Peaks of FAMEs were identified through comparison to authentic standards.
2.6. Polysaccharide Determination
Approximately 20 mg of fat-free biomass (x_f_) was hydrolyzed with 5 mL HCl (Sigma-Aldrich) 2.5 M at 100 °C for 60 min. When the hydrolysate was cooled at room temperature, it was neutralized with 2.5 M KOH (Sigma-Aldrich) and filtered through Whatman No. 1 paper so as to remove cell debris. The reducing sugars, expressed as glucose, were determined in the solution in accordance with the DNS method [37]. Intracellular polysaccharides, including storage and structural polysaccharides, were expressed (as glucose equivalents) as a percentage of dry biomass (S/x%, wt/wt).
2.7. Protein Determination
Total cellular protein was determined using approximately 10 mg of x_f_ following the biuret method, as described elsewhere [33]. Cellular protein was expressed (as albumin equivalents) as a percentage of dry biomass (P/x%, wt/wt).
2.8. Pigment Estimation
Approximately 0.5 g of wet biomass was solubilized with 10 mL of ethanol 95%, vol/vol (Fisher Chemical) solved with Milli Q water (Honeywell, Charlotte, NC, USA). The mixture was centrifuged (Hettich Mikro 200R, Föhrenstr, Germany) at 13,300× g for 15 min at 4 °C. Then, 0.5 mL of the supernatant was collected and mixed with 4.5 mL of the ethanol solution [38]. The resulting solution was spectrophotometrically analyzed for Chlorophyll-a, Chlorophyll-b, and carotenoids in a quartz cuvette (1 cm^2^), while ethanol solvent was used as a blank.
The Equations (3)–(5) used for the pigment quantification (in µg/mL) were the following [22]:
where A represents Absorbance and C_a_, C_b_, and C_x+c_ stand for Chlorophyll-a, Chlorophyll-b, and carotenoids, respectively. Pigments, evaluated as Total Chlorophylls (TCh) and Total Carotenoids (TC), were expressed as percentage in the dry biomass (TCh/x%, wt/wt, and TC/x%, wt/wt).
2.9. Data Treatment and Statistical Analysis
The graphing and analysis software OriginPro 2021 9.8.0.200 ^®^, 1991–2020 (OriginLab Corp., Northampton, MA, USA) was used for the treatment of the experimental data from microalgal cultures and growth kinetics.
3. Results
3.1. Cell Growth and Biomass Production
The growth curves of all strains under the examined growth conditions are collectively depicted in Figure 2, while the results regarding growth parameters are compiled in Table 2.
The strain P. costavermella VAS2.5 grew satisfactorily at both incubation temperatures tested, especially at T = 25 °C (Figure 2a), leading to higher biomass production as well (x = 671.3 ± 17.2 mg/L with p_x_ = 67.1 ± 1.7 mg/L∙d and μ = 0.45 ± 0.05 1/d), compared to 418.4 ± 17.7 mg/L of the other culture condition (Table 2). It is worth mentioning that due to the similarity of the conceivable curve that could express the trend of the cytometry points with a straight line when this strain was cultured at T = 25 °C, the linear model was also applied to the data, showing a very good fit (R^2^ = 0.98) (see respective dοtted line plot in Figure 2a). Similar observations were made regarding the culture at T = 20 °C (see respective dotted line plot in Figure 2a) as well as for the other two strains that belong to the Picochlorum genus and were used in the current study. Intriguingly, though, the other two Picochlorum isolates, namely, P. oklahomense SAG4.4 and P. oklahomense PAT3.2B, showed growth capacity only when incubated at T = 25 °C (Figure 2b,c). Under these culture conditions, both strains reached similar biomass production levels. Specifically, P. oklahomense SAG4.4 biomass production reached 224.8 ± 5.9 mg/L (p_x_ = 22.5 ± 0.7 mg/L∙d and μ = 0.39 ± 0.08 1/d) and P. oklahomense PAT3.2B produced 242.7 ± 13.6 mg/L (p_x_ = 24.3 ± 1.4 mg/ L∙d and μ = 0.47 ± 0.07 1/d) (Table 2).
Regarding the cultures of M. gaditana VON5.3, this strain grew satisfactorily at both examined temperatures, but clearly more efficiently when incubated at T = 25 °C (Figure 2d). As mentioned before for P. costavermella VAS2.5 cultured at T = 25 °C, the growth curve of the M. gaditana VON5.3 culture at the same temperature was also well-fitted to the linear model as shown by the dotted line (Figure 2d) (R^2^ = 0.90 and R^2^ = 0.88 for linear model and the integrated version of Verhulst’s model, respectively). As regards the biomass production, M. gaditana VON5.3 gained practically the same level of biomass at both incubation temperatures (Table 2). In particular, it was x = 527.4 ± 15.0 mg/L (p_x_ = 52.7 ± 1.5 mg/L∙d and μ = 0.35 ± 0.11 1/d) and x = 509.0 ± 77.8 mg/L (p_x_ = 50.9 ± 7.8 mg/L∙d and μ = 0.56 ± 0.12 1/d) for the culture at T = 25 °C and T = 20 °C, respectively.
The strain N. pyriformis PAT2.7 also grew satisfactorily at both incubation temperatures (Figure 2e). Interestingly, in this set of experiments, the biomass produced was lower when the microalga grew at T = 25 °C, i.e., x = 438.8 ± 89.3 mg/L (with p_x_ = 43.9 ± 8.9 mg/L d and μ = 1.47 ± 0.34 1/d) compared to x = 546.7 ± 1.2 mg/L (with p_x_ = 54.7 ± 0.1 mg/L∙d and μ = 0.64 ± 0.12 1/d) at T = 20 °C (Table 2).
3.2. Accumulation of Storage Materials
The results of storage material accumulation levels at the end of each culture performed are compiled in Table 2.
Regarding P. costavermella VAS2.5, it was observed that the lipid content was clearly higher at T = 25 °C (L/x% = 19.3 ± 0.7%, wt/wt), whereas at T = 20 °C, the lipid yield was relatively low (L/x% = 5.2 ± 0.3%, wt/wt). The other two Picochlorum strains, which were able to grow only at T = 25 °C, synthesized fewer lipids, i.e., L/x% = 4.1 ± 0.4%, wt/wt and L/x% = 11.5 ± 0.3%, wt/wt, for P. oklahomense SAG4.4 and P. oklahomense PAT3.2B, respectively (Table 2). Additionally, the percentages (%, wt/wt) of lipid fractions (N—neutral lipids, G + S, glycolipids, and sphingolipids, P—phospholipids) to total lipids (TLs) are also presented in Table 2. Herein, though, such investigation occurred only for P. costavermella VAS2.5 cultured at T = 25 °C, as it was not possible to investigate the contribution of individual lipid fractions to total lipids of the biomass collected from the lower temperature, due to the small amount of lipids produced. Likewise, the aforementioned results apply to the two P. oklahomense strains as well. In this respect, the predominant lipid fraction of P. costavermella VAS2.5 was that of G + S (57.4 ± 5.4%, wt/wt), followed by the N fraction (29.6 ± 3.4%, wt/wt) and P (13.1 ± 2.0%, wt/wt). M. gaditana VON5.3 produced biomass in which the reserve lipid content was slightly higher at T = 25 °C (L/x% = 16.4 ± 2.1%, wt/wt) compared to the culture at T = 20 °C (L/x% = 13.8 ± 0.0%, wt/wt). The dominant lipid fraction was that of N (55.6 ± 1.6%, wt/wt), followed by the G + S fraction (24.7 ± 1.4%, wt/wt) and P (19.8 ± 0.3%, wt/wt) when the microalgae was grown at T = 25 °C. On the contrary, the highest percentage was recorded for the G + S fraction (41, 3 ± 2.0%, wt/wt) after culture at T = 20 °C, with P also following at high percentages (36.3 ± 2.1%, wt/wt), and finally, the N fraction (22.5 ± 0.2%, wt/wt) (Table 2). Lastly, as regards the biomass content of N. pyriformis PAT2.7 to reserve lipids in the two incubation conditions, a higher percentage was observed in the culture at T = 20 °C (L/x% = 9.3 ± 6.1%, wt/wt) compared to that at T = 25 °C (L/x% = 3.3 ± 1.7%, wt/wt) (Table 2). When this strain was grown at T = 25 °C, the highest percentage belonged to the lipid fraction G + S (48.6 ± 5.0%, wt/wt) which was marginally higher than the N fraction (45.8 ± 4.3%, wt/wt), and the last one was the P fraction (6.0 ± 1.0%, wt/wt). In culture at 20 °C, there was a clear dominance of the N fraction (63.5 ± 6.0%, wt/wt), followed by G + S (34.8 ± 4.8%, wt/wt) and P (2.3 ± 1.2%, wt/wt) (Table 2).
Regarding the ability of P. costavermella VAS2.5 to store polysaccharides and proteins under the examined culture conditions, a significant difference was recorded in the accumulation of polysaccharides (S/x% = 13.5 ± 0.8 and S/x% = 6.7 ± 0.1, for cultures at T = 25 °C and T = 20 °C, respectively), but not in protein content (P/x% ranged in the 25.0–26.7%, wt/wt range) (Table 2). Similar levels of polysaccharides and protein synthesis were recorded for P. oklahomense SAG4.4 (S/x% = 11.3 ± 0.4%, wt/wt and P/x% = 28.8 ± 1.0%, wt/wt) cultured at T = 25 °C (Table 2). On the other hand, despite polysaccharide accumulation being similar to the above for P. oklahomense PAT3.2B (i.e., S/x% = 11.2 ± 1.2%, wt/wt), the exhibited protein content of the biomass was significantly higher (P/x% = 50.6 ± 3.0%, wt/wt) (Table 2). Regarding the ability of M. gaditana VON5.3 to store polysaccharides and proteins under the two culture conditions, similar percentages with little variation were also recorded (S/x% = 8.5–9.4% and P/x% = 16.4–17.5%) (Table 2). In the same manner, small variations were recorded in the case of N. pyriformis PAT2.7, though the recorded protein content was high (S/x% = 8.0–10.7% and P/x% = 41.0–45.2%) (Table 2).
Finally, the values of total chlorophyll and carotenoids in dry biomass were recorded (Table 2). The obtained results presented similar levels of chlorophyll biosynthesis (the percentage of which ranged between 2.1 and 2.6%, wt/wt) and slightly higher levels of carotenoids when P. costavermella was grown at T = 20 °C (0.8% at t = 20 °C versus 0.2%, wt/wt at T = 25 °C). On the contrary, the cultures of both P. oklahomense strains at T = 25 °C led to higher total chlorophyll (TCh/x% = 4.9 ± 0.1%, wt/wt and TCh/x% = 6.1 ± 0.7%, wt/wt, for SAG4.4 and PAT3.2B, respectively) and carotenoid (TC/x% = 0.8 ± 0.0%, wt/wt and TC/x% = 1.1 ± 0.1%, wt/wt, for SAG4.4 and PAT3.2B, respectively) content of the cells. Respective analyses on M. gaditana VON5.3 biomass showed higher levels of chlorophyll biosynthesis when the microalga was grown at T = 20 °C (TCh/x = 3.2% vs. 1.4%) and similar rates of carotenoid biosynthesis when grown at T = 25 °C (i.e., TCh/x = 0.3–0.9%, wt/wt and TC/x = 0.1–0.3%, wt/wt). Finally, total chlorophyll and carotenoid contents were at low levels in both culture conditions tested for N. pyriformis PAT2.7 (in the range of TCh/x% = 1.0–1.7%, wt/wt, and TC/x% = 0.3%, wt/wt).
3.3. Fatty Acid Composition of Total Lipids and Lipid Fractions
The composition of total lipids and lipid fractions in fatty acids at the end of the cultures under both culture conditions are listed in Table 3, Table 4 and Table 5.
Regarding the culture of P. costavermella VAS2.5 at T = 25 °C, it is observed that in TLs the predominant fatty acids are C16:0 (29.0 ± 0.2%, wt/wt) and ^Δ9^C16:1 (28.6 ± 0.3%, wt/wt), followed by ^Δ5,8,11,14,17^C20:5 presenting in a significant proportion (17.0 ± 0.2%, wt/wt). The N lipid fraction showed a similar fatty acid composition to that of the TLs, except that the proportion of ^Δ5,8,11,14,17^C20:5 in it was significantly lower (5.4 ± 0.1%, wt/wt). The percentages of fatty acids within the G lipid fraction were similar to those of TLs. With regard to the P fraction, a decrease in the percentage of C16:0 and ^Δ9^C16:1 was observed, which was accompanied by a significant increase in the percentage of ^Δ5,8,11,14,17^C20:5 to 30.3 ± 4.9% (wt/wt), compared to TLs. The three aforementioned fatty acids were also dominant in the TL cultures at T = 20 °C; however, in this case, limited biosynthesis of C16:0 was observed (14.3 ± 0.6%, wt/wt), in contrast to ^Δ5,8,11,14,17^C20:5, whose concentration was higher (35.9 ± 0.7%, wt/wt), compared to that of lipids produced at T = 25 °C.
Regarding the fatty acid composition of total lipids produced by P. oklahomense SAG4.4, the predominant fatty acid is the rare fatty acid ^Δ6,9,12,15^C18:4, (18.9 ± 3.6%, wt/wt), followed at similar levels by ^Δ9^C18:1 (18.4 ± 2.4%, wt/wt), while C16:0 (12.2 ± 2.1%, wt/wt) came in third place (Table 3). Also, Table 3 shows that the fatty acid ^Δ9,12,15^C18:3 was also present at remarkable levels (6.3 ± 0.6%, wt/wt). On the other hand, in the case of P. oklahomense PAT3.2B, the fatty acid detected in the highest percentage was ^Δ9,12^C18:2 (25.3 ± 0.0%, wt/wt), followed by the fatty acids ^Δ9,12,15^C18:3 (17.9 ± 1.0%, wt/wt), C16:0 (15.2 ± 0.1%, wt/wt), and ^Δ9^C18:1 (14.4 ± 0.8%, wt/wt) (Table 3). Other fatty acids detected, but at significantly lower percentages, were ^Δ9^C14:1 (9.3 ± 0.3, wt/wt), C18:0 (6.4 ± 0.7, wt/wt), ^Δ9^C16:1 (4.4 ± 0.4, wt/wt), and C14:0 (2.3 ± 0.2, wt/wt) (Table 3).
In the case of M. gaditana VON5.3, it was observed for the culture at T = 25 °C that in TLs the dominant fatty acids were ^Δ9^C16:1 (30.5 ± 0.8%, wt/wt) and C16:0 (24.1 ± 1.3%, wt/wt), followed by ^Δ5,8,11,14,17^C20:5 (19.9 ± 0.1%, wt/wt) (Table 4). The lipid N fraction showed a similar pattern to that of TLs, but contained ^Δ5,8,11,14,17^C20:5 at a significantly lower percentage (10.4 ± 1.9%, wt/wt). The lipid fraction G showed a similar fatty acid composition to TLs, while the P fraction showed a small percentage of ^Δ9^C16:1 and a large percentage for C16:0 (36.3 ± 1.0%, wt/wt) and C18:0 (17.6 ± 0.9%, wt/wt) fatty acids, while ^Δ5,8,11,14,17^C20:5 was detected at the levels of the N fraction. The three fatty acids that dominated at 25 °C were also those with the highest percentages when M. gaditana was grown at 20 °C, and at similar levels to those determined in the culture incubated at T = 25 °C (Table 4). As shown in Table 4, there is a correspondence with the 25 °C growth case in terms of the fatty acid composition of the lipid fractions, with the only difference being the high percentage of ^Δ5,8,11,14,17^C20:5 (29.5 ± 3.3%, wt/wt) in the lipid fraction of P.
Lastly, the predominant fatty acids of N. pyriformis PAT2.7 were ^Δ9^C16:1 (40.1 ± 0.1%, wt/wt) and C14:0 (31.0 ± 0.8%, wt/wt) in TLs when this strain was grown at T = 25 °C, and the N and G lipid fractions showed a similar pattern to that of TLs (Table 5). However, the presence of C14:0 at lower amounts (15.5 ± 0.7%, wt/wt) was recorded for the P lipid fraction and a remarkable percentage for ^Δ9^C18:1 (19.4 ± 2.3%, wt/wt), compared to TLs and the two aforementioned lipid fractions. When N. pyriformis was grown at T = 20 °C, a similar pattern to the 25 °C growth was observed in terms of the composition and fatty acid content of total lipids and their lipid fractions (Table 5).
4. Discussion
Microalgae are used in a plethora of biotechnological applications and several species are being used to produce several tons of biomass and metabolites of high biotechnological value [4,5,21]. Concurrently, microalgae are an important biological material for basic research [39]. The main cellular components that determine the caloric value and quality of biomass of photosynthetic microorganisms intended for feeding terrestrial and aquatic organisms are lipids, proteins, and polysaccharides [3,4,21]. Some species are of additional interest because of their ability to synthesize lipids that contain PUFAs in high concentrations. PUFAs are quite popular in the food, feed, and pharmaceutical industries due to their proven beneficial effects against various threats to human and animal health [9,10,11]. On the other hand, species which contain more saturated fatty acids can be used in biodiesel production [7,21,40].
All isolates used in the present study were previously characterized biochemically and cultured under laboratory conditions, exhibiting satisfactory yields regarding growth, biomass, and high added-value metabolite production [13]. Herein, special emphasis was given to the ability of the selected microalgal strains to accumulate significant amounts of lipids rich in PUFAs (i.e., Picochlora and M. gaditana VON5.3), or more saturated lipids suitable for biodiesel (i.e., N. pyriformis PAT2.7) and, secondly, to polysaccharide, protein, and pigment production. The predominant reserve material in all microalgae was proteins, followed by lipids and polysaccharides. Another factor for the selection of these strains was the limited literature available on species of the genera Picochlorum and Nephroselmis; thus, the opportunity to offer useful data regarding exploring the exploitation potential of such microorganisms has arisen.
The STR was chosen for the cultures performed since it offers better control on culture conditions, e.g., regarding temperature, aeration, agitation, etc. Another crucial factor was the ability of considerable high illumination supply (i.e., 1071 μmol/m^2^∙s). Moreover, Sorokin and Krauss (1958) [41], in early studies, after examination of several strains of the genus Chlorella (related to Picochlorum as both genera belong to the order Chlorellales), under light intensities of about 300 μmol/m^2^∙s and higher, concluded that in green microalgae the maximum growth rate is achieved under continuous illumination at intensities up to saturation of the photosynthetic capacity of the microalgae. The aforementioned conclusions are also confirmed by the later study of Sukenin and Carmeli (1989) [42] who, apart from biomass production, linked the synthesis of lipids and polysaccharides to the intensity of light provided to a Nannochloropsis sp. strain grown in F/2 nutrient medium on turbidostat at three different light intensities (35 μmol/m^2^∙s, 290 μmol/m^2^∙s, 550 μmol/m^2^∙s). De la Vega et al. (2011) [43], at supplied light intensities of 100 and 1,200 μE/m^2^∙s, recorded a small increase in the specific growth rate (from 0.031 1/h to 0.034 1/h) of Picochlorum sp. HM1, concluding that its growth is nearly saturated at relatively low light intensities, while no photoinhibition was observed for light intensities of 1,200 μE/m^2^∙s. In any case, it has to be noted that the findings of this research may suggest the existence of two limiting factors for growth, which were probably the dissolved CO_2_ concentration, as atmospheric air, containing 0.03% CO_2_, was provided to the cultures, and lighting, which is often a limiting factor in high volume cultures.
Another parameter to be considered regarding the light energy supply, which can potentially influence biomass growth, is ‘mutual shading’ [44,45,46]. Specifically, an increase in cell concentration can be associated with an increase in the ‘mutual shading’ caused by preventing light from penetrating deeper into the culture. This phenomenon is also observed in the natural environment of microalgae, which can cause shading of either cells of the same species or cells of other species, affecting the balance of the ecosystem. In this respect, the lateral light supply in the agitated cultures carried out in the STR contributed positively to a better uptake of light energy by the cells. In this context, a safe strategy to increase the synthesis of pigments, especially carotenoids, is considered to be the provision of high light intensity or a gradual increase in temperature in the culture environment, as both are parameters that stress the cells and activate the photoprotection mechanisms of microalgae [47]. However, in the present study, regardless of culture conditions, a generally similar, but not significantly high, production of chlorophylls was observed when comparing to other studies [29,48,49,50,51,52]. Of note, the pigment content of M. gaditana VON5.3 was higher than other strains, e.g., Nannochloropsis sp., used in their study by Magpusao et al. (2024) [53]. In any case, the synthesis of chlorophyll-a was higher than chlorophyll-b (i.e., results are given as total chlorophyll values) in all measurements, which is in agreement with the findings other studies [48,49]. However, since all the examined strains are known producers of carotenoids, rich in lutein and zeaxanthin beta-carotene, siphonaxanthin, neoxanthin, etc. [20,43,53,54,55,56], which are of high biotechnological value due to their proven antioxidant and anti-inflammatory activity, culture conditions should be modified when the desired product is these molecules.
Herein, the conclusion drawn from the cultures in the STR of the three Picochlora strains was that they all exhibited relatively high specific growth rates and gained satisfying amounts of biomass. The results were comparable or better to the cultures previously performed in Erlenmeyer flasks [13]. Specifically, P. costavermella VAS2.5 produced biomass that exceeded 670 mg/L at T = 25 °C, i.e., almost twice the size compared to the experiments in Erlenmeyer flasks [13], while even at T = 20 °C, biomass production reached 418 mg/L. On the other hand, both P. oklahomense strains, which were able to grow only at T = 25 °C, reached similar biomass values between them and when compared to their respective cultures in Erlenmeyer flasks [13]. The preference of Picochlora for relatively high temperatures has been recorded elsewhere as well [47,57]; thus, the response of the P. oklahomense strains came as no big surprise. In any case, taking into consideration that the cultures of this investigation lasted half the time of cultures in Erlenmeyer flasks, it is highlighted that, for these strains, light supply played a key role in increased productivities. Notably, the potential of Picochlora to gain high biomass yields is confirmed by other studies as well. For example, Zhu and Dunford (2013) [58] who cultured a P. oklahomense strain in a 2 L bioreactor at medium F/2 and even applied a 12:12 photoperiod, recorded biomass production at a significantly higher level (x = 2.1 g/L). Intriguingly, strains such as Picochlorum sp. HM1 [43] and Picochlorum celeri [59], which present short generation times and similar growth rates with the strains used herein, are directly competitive to commercial strains belonging to genera such as Dunaliella and Nannochloropsis. However, despite the aforementioned studies, it must be taken into account that, to date, Picochlora have not been used extensively in commercial/industrial scale applications. Specifically, to the best of our knowledge, the only study involving the culture of a Picochlorum oculatum strain at pilot (V_w_ = 150 L) and commercial scale (V_w_ = 2000 L) in an open horizontal bioreactor outdoors belongs to Dogaris et al. (2019) [60]. The researchers concluded that this strain can be a good candidate for use in the biodiesel production industry. From the above study, it can be concluded that strains of the genus Picochlorum could pose as attractive candidates for large-scale applications.
M. gaditana VON5.3 grown in an STR showed a higher specific growth rate and higher biomass production, similarly to Picochlora strains, compared to its culture in Erlenmeyer flasks [13]. However, it is worth mentioning that in this study a similar amount of biomass was also produced when this strain was cultured at T = 20 °C, demonstrating its very good adaptation even in lower temperature environments. In agreement with the observations on the positive effect of incubation under high light intensity, another study on N. salina also shows a positive correlation of the increase in biomass production with the intensity of the light energy provided [61]. On the other hand, N. salina grown in mASW in what resembles an open-pond system reactor at 120 μE/m^2^∙s, produced biomass of equivalent amount to that of M. gaditana VON5.3 [33]. Simionato et al. (2011) [62] supported that within a certain light energy threshold little change in biomass production occurs, while only under extreme light intensities can significant changes in biomass production be observed. However, when N. oculata was grown in 2 L containers on growth medium F/2, with a light output of 56 ± 4 μmol/m^2^∙s and photoperiod of 12:12, it produced biomass that reached 1.2 g/L, though in a culture that lasted almost as long [58].
N. pyriformis PAT2.7 showed high specific rate values. Even though N. pyriformis PAT2.7 gained more biomass when grown in Erlenmeyer flasks [13], significant biomass production was recorded in STR at both examined temperatures, as well, especially taking into consideration that these cultures lasted half the time compared to the ones in Erlenmeyer flasks. Noteworthily, in this case, growth appeared to be more efficient when N. pyriformis was cultured at T = 20 °C. The results of the research by Hotos and Avramidou (2021) [63] seem to verify the above observation, since they recorded satisfactory growth and remarkable biomass production from another Nephroselmis strain when cultured at T = 20–21.5 °C. Nevertheless, it should be mentioned that other studies recorded the production of equivalent or even more biomass when representatives of the genus were grown at higher temperatures (e.g., 25–27 °C) [29,40,64]. From the above observations, it can be concluded that species of this genus exhibit plasticity responsive to different temperature values, with the optimal growth temperature varying between species or even strains. This observation suggests that the intensity of the light energy provided, although clearly high, had neither a positive nor a negative effect on the growth of the microalgae. However, it should be mentioned that the intershading from the very day of the cultures played an important role since the cultures showed a dark green color, indicating the insufficient availability of the supplied light in the inner part of the culture volume.
With regard to lipid biosynthesis, the lowest yields for P. costavermella VAS2.5 were observed when grown at 20 °C, which can be attributed to the environmental stress conditions and the need to maintain cell homeostasis. All other values regarding Picochlora strains used in this study were comparable to their respective cultures in Erlenmeyer flasks [13]. Lipid accumulation of the P. oklahomense strain used by Zhu and Dunford (2013) [58] was similar to the one presented by P. costavermella VAS2.5 at T = 25 °C. However, Tran et al. (2014) [65] characterized a strain of the genus that gained up to L/x% = 48.6% (wt/wt) under stress conditions and low temperature (T = 15 °C), while its growth rate was not severely affected. The aforementioned observations confirm the plasticity of the representatives of this genus. Higher lipid content was observed for M. gaditana VON5.3 compared to when it was grown in Erlenmeyer flasks (i.e., L/x% = 9.3 ± 3.0%, wt/wt) [13] at both culture temperatures, while no significant effect of temperature was recorded for the amount of lipids synthesized between the two cultures. The increased yields compared to the cultures in Erlenmeyer flasks can be accounted for by the high light energy supplied to the cultures grown in this type of bioreactor, in accordance with the findings of Mohammady (2014) [61]. In any case, lipid content of this strain was comparable to that of the commercial strain Nannochloropsis sp. of Reed Mariculture (USA), which though it exceeded 17% (wt/wt), was significantly lower than N. oceanica F&M-M24, which approached 30% (wt/wt) [53,66]. With N. pyriformis PAT2.7 cultures, lipid synthesis was low at T = 25 °C, while it was considered adequate under culture at T = 20 °C. The available literature on the ability of Nephroselmis to accumulate lipids is quite limited, yet the results of the present study are comparable to those of another Nephroselmis strain (L/x% = 10.5%, wt/wt), which was grown for about one week in F nutrient medium at 25 °C, with a salinity of 28‰ and a light intensity of 80–100 μmol/m^2^∙s [67]. However, the Nephroselmis sp. that Hotos et al. (2023) [68] cultured in Walne medium accumulated lipids at 15%, wt/wt. Even higher lipid accumulation was exhibited by Nephroselmis sp. KGE1, also grown on BBM, with L/x% = 33.0 ± 0.06% (wt/wt) [64]. BBM, taking into account its composition, is a growth medium more suitable for freshwater microalgae, highlighting the adaptability of Nephroselmis in various aquatic systems, which has been confirmed elsewhere (e.g., Hotos et al. (2020) [69]). Yet, it could also be noted that BBM contains KNO_3_ = 0.25 g/L as a nitrogen source, compared to 1 g/L contained in mASW, implying that in Nephroselmis lipid production is be significantly influenced by the availability of nitrogen rather than the temperature in the growth environment.
Regarding the content of total lipids in their lipid fractions, it must be taken into consideration that a common feature for a plethora of microalgae is the dominance of the G + S fraction over total lipids, as has been reported for strains of various genera, like Chlorella, Nannochloropsis, and Tetraselmis [33,34,49,70]. The above is answered by the fact that G + S are the main components (in percentages up to 80–90%) of lipids of chloroplast and thylakoid membranes [71,72,73,74]. Glycolipids of microalgae are considered an important source of omega-3 fatty acids, while they can also play the role of storage material [75,76]. Sphingolipids are a diverse group of membrane lipids that were highly conserved during evolution and play an important role in cell communication with its environment and other cells as well as in several important cell functions, such as the control of cell division [77,78]. The P fraction also belongs to the predominant structural lipids in microalgae, as phospholipids are a key component of all cell membranes. Biosynthesis of the polar lipids is strongly affected by several factors, such as the composition of the nutrient medium, the provided light intensity and photoperiod, and the growth phase of the culture [79]. However, one of the most influential factors is environmental temperature. In low temperature, the presence of saturated lipids in cell membranes reduces their fluidity and negatively affects cellular metabolism. Consequently, an increased synthesis of unsaturated fatty acids is expected, which could increase survival under such adverse conditions [26,27].
The main lipid fraction, for P. costavermella VAS2.5 at T = 25 °C, the sole case that was possible to perform such analyses among the cultures performed using the Picochlora strains, was that of G + S, followed by N lipid fraction and then P. In the same manner, from the cultures of M. gaditana VON5.3, the fraction G + S was also expected to be the dominant one, as documented elsewhere [33,34]. However, when this strain was cultured at T = 25 °C, the N fraction dominated; albeit, this could be attributed to the high light intensity [80]. N. gaditana 1049 was also shown to synthesize mainly N lipids, though after the 16th day of cultivation [81]. The authors interpreted this phenomenon as a consequence of a shift in the biochemical pathways of lipid biosynthesis of an aged culture from chloroplasts and other cell membranes mainly towards N lipid production. Another study denoted growth of Nannochloropsis sp. SCSIO-45217 under phosphorus-limiting conditions as favorable for synthesis of N lipids [82]. In contrast, when was M. gaditana VON5.3 cultured at T = 20 °C, it presented high G + S content, aligning with the aforementioned observations regarding the need of cells to remodel their cell wall and photosynthetic system in response to low temperature. Overall, since G + S and P are more abundant in PUFAs, the M. gaditana strain VON5.3, especially under relatively low temperatures, appears to be a particularly attractive candidate for production of PUFAs in large-scale applications. In contrast to the other microalgae, N. pyriformis PAT2.7 proved to be a major producer of N lipids and, secondarily, of G + S. The very high content in N lipid reserves is another indication of the ability of this microalga to survive at T = 20 °C.
In any case, the examined microalgal strains presented a notable fatty acid composition. P. costavermella VAS2.5 synthesized ^Δ5,8,11,14,17^C20:5 at high concentrations, exceeding the P fraction, where the PUFAs are biosynthesized, 30% (wt/wt) at 25 °C, or even 35% (wt/wt) at 20 °C in total lipids. The ^Δ5,8,11,14,17^C20:5 content of this strain can explain the very low percentages or even the occasional absence of ^Δ9,12,15^C18:3 and ^Δ6,9,12,15^C18:4. According to the mechanisms described by Bellou et al. (2014) [4] and Jesionowska et al. (2023) [83], very long chain PUFAs (VLC-PUFAs) are synthesized by the catalytic action of desaturases and elongases on ^Δ9^C18:1 via the ω-3 and/or ω-6 pathways. In brief, C16:0 is initially synthesized within chloroplasts through the enzymatic action of the fatty acid synthase (FAS) complex and is then converted to C18:0 by the addition of two carbon atoms derived from acetyl-CoA. The latter one is then desaturated to ^Δ9^C18:1 which in its activated form is transferred to the endoplasmic reticulum where it is further desaturated by Δ12 and Δ15 desaturases and produces ^Δ9,12^C18:2 and ^Δ9,12,15^C18:3. The last ones serve as precursors for the ω-6 and ω-3 pathways, respectively, for the biosynthesis of VLC-PUFA. In the ω-3 biosynthetic pathway, ^Δ9,12,15^C18:3 is converted to ^Δ6,9,12,15^C18:4, ^Δ8,11,14,17^C20:4, and then to ^Δ5,8,11,14,17^C20:5 by desaturase Δ6, elongase Δ6, and desaturase Δ5, respectively. ^Δ5,8,11,14,17^C20:5 is a PUFA of high biotechnological significance, since it is a biosynthetic precursor of prostaglandins-3 which inhibit platelet aggregation, thromboxane-3, and eicosanoids [83]. The presence of this fatty acid appears to have been recorded in only one other representative of the genus, though at much lower levels [65].
The main fatty acids produced by M. gaditana VON5.3 were the same as when grown in Erlenmeyer flasks [13], namely, ^Δ9^C16:1 and ^Δ5,8,11,14,17^C20:5. Total lipids and lipid fractions of M. gaditana VON5.3 were generally similar in composition, and the ^Δ5,8,11,14,17^C20:5 was detected in high proportions, in polar lipids. At the same time, ^Δ9^C16:1 was also high in polar lipids, especially at T = 20 °C. A similar pattern in total lipids and their lipid fractions was also recorded for other Microchloropsis and Nannochloropsis strains elsewhere [33,34,53]. More importantly, M. gaditana VON5.3 showed a significantly higher ^Δ5,8,11,14,17^C20:5 content compared to the nine Nannochloropsis and Microchloropsis strains used by Ma et al. (2014) [84]. In their study, the most productive strain in terms of ^Δ5,8,11,14,17^C20:5 did not exceed 13% (wt/wt) on total lipids. Also, Nannochloropsis sp. SCSIO-45217 presented significantly lower ^Δ5,8,11,14,17^C20:5 content (i.e., 2.1–7.3%, wt/wt) than M. gaditana VON5.3 when grown under phosphorus-limiting conditions (at concentrations of 4–40 mg/L), albeit it synthesized ^Δ9^C16:1 in significant amounts (35.2–37.1%, wt/wt) [82]. Even lower was the lipid content of the aforementioned fatty acids in the case of N. oceanica F&M-M24 [66]. The above observations suggest the ability and suitability of M. gaditana VON5.3 as a source of lipid production of high nutritional and medicinal value, especially cultured at T = 20 °C, which presented further enhanced yields in PUFA synthesis.
N. pyriformis PAT2.7 synthesized mainly saturated fatty acids and an absence of PUFAs was recorded, with the exception of traces of ^Δ6,9,12^C18:3 detected in some cases. These features highlight N. pyriformis PAT2.7 as a remarkable candidate for the biodiesel production industry. The predominant fatty acid in all cases was ^Δ9^C16:1 at high percentages of total lipids. Moreover, ^Δ9^C16:1 occurred in similar proportions in the N and G + S lipid fractions, while its proportion was reduced to about half in the P fraction. The significant production of ^Δ9^C16:1 is considered to be of outstanding importance and possibly opens up new prospects for the exploitation of this microalga, since ^Δ9^C16:1 is attributed with anti-inflammatory properties and the ability to reprogram the intestinal microflora. Other strains of the genus synthesized much smaller amounts of ^Δ9^C16:1 (3.9–6.1%, wt/wt) [40,67] but also linolenic acids (^Δ6,9,12^C18:3 and ^Δ9,12,15^C18:3), which in total approached 15% (wt/wt) of total lipids. Even higher yields were recorded in other studies regarding ^Δ6,9,12^C18:3 content of total lipids (i.e., 37.7–56.4%) [51,64]. The above studies further support the intra-species variability and the need to select the proper species/strain in correspondence to the desired final product.
Regarding polysaccharide accumulation in Picochlora strains, their content was almost double in comparison with their respective cultures in Erlenmeyer flasks [13], yet not in considerable amounts. Similarly, M. gaditana VON5.3 synthesized polysaccharides at slightly higher levels compared to Dritsas et al. (2023) [13]’s respective culture and at comparable amounts to the N. salina used by Bellou and Aggelis (2012) [33]. However, strains that belong to Microchloropsis and Nannochloropsis can accumulate polysaccharides in higher levels (i.e., 20–25%, wt/wt) [34,53,66]. N. pyriformis PAT2.7 produced polysaccharides in the same range of values with other strains of the genus [13,51,67]. According to Bellou and Aggelis (2012) [33], the amount of intracellular reserve materials is linked with the growth phase of microalgae, while the authors also considered reduced photosynthesis and CO_2_ availability as crucial factors. Specifically, they concluded that the passage of light through the culture and CO_2_ captured at high cell densities can lead to not covering the metabolic needs of the cells in the culture. Consequently, the cells meet the need for energy and metabolic precursors at the expense of the reserve materials. In addition, it is worth noting that Mastropetros et al. (2023) [52] suggest high salinity and light intensity as environmental stressors for increased polysaccharide production in Nephroselmis.
Regarding the protein content of Picochlorum strains, high accumulation rates were recorded independent of culture temperature, especially for P. oklahomense PAT3.2B, in which protein content exceeding 50% (wt/wt). The protein synthesis was comparable to the cultures of these strains in Erlenmeyer flasks [13]. The above did not come as a surprise, as generally the ability of Picochlora to accumulate proteins at high levels has been documented elsewhere [48,49,58,85]. Interestingly, the existence of amino acid profiles of high commercial and biotechnological interest has been reported for other strains of the genus [56,65,86]. When M. gaditana VON5.3 was grown in the STR, the protein content of the biomass was about half compared to the experiments in Erlenmeyer flasks [13], which may possibly be explained by the increased metabolic activity of the cells and the high biomass production in a relatively short incubation period. In each case, the protein content under these conditions was comparable to that of other Microchloropsis or Nannochloropsis strains [33,34,58], while temperature practically did not affect protein synthesis. On the other hand, a particularly high content was observed in N. pyriformis PAT2.7 cultures in both temperatures, slightly higher when this microalga was cultured at T = 20 °C. Protein synthesis was comparable to the cultures of this strain previously performed in Erlenmeyer flasks [13]. The high protein content of N. pyriformis PAT2.7 grown at 20 °C demonstrates its good adaptation to these conditions and suggests the suitability of the strain for large-scale outdoor cultivation where temperatures are low during the winter months. Moreover, the very high protein content of Nephroselmis highlights it as capable of being utilized in the diet of fish (directly or indirectly) or even humans. It appears that under conditions of environmental stress the microalgae channel energy towards energy conservation and/or growth rather than towards protein synthesis.
Taking into consideration the above observations, it reasonably follows the deduction and the ambition that this investigation can contribute to the design of the proper culture conditions, taking into account parameters such as temperature, adapted to the species or even strains of the same species, with the aim to optimize yields of the desired products. This would offer the possibility and the prospect for the development of various applications of microalgae, both in the aquaculture sector and in the production of high added-value products through technologies compatible with green growth.
5. Conclusions
Cultures of the microalgal strains Picochlorum costavermella VAS2.5, Picochlorum oklahomense SAG4.4, Picochlorum oklahomense PAT3.2B, Microchloropsis gaditana VON5.3, and Nephroselmis pyriformis PAT2.7 at T = 25 °C led to significant biomass production in most cases, especially taking into account that they all lasted for almost 10 days. Notably, P. costavermella VAS2.5, M. gaditana VON5.3, and N. pyriformis PAT2.7 grew efficiently even at T = 20 °C. Except for the genetic plasticity of these strains, the above results suggest that, in addition to CO_2_ possibly being a limiting factor, the high and lateral, which confined ‘cross-shading’, light supply in the STR played a decisive role in the increase in cell number regardless of culture temperature. The most favorable condition for lipid accumulation proved to be incubation at 25 °C for P. costavermella VAS2.5 and M. gaditana VON5.3, though N. pyriformis PAT2.7 increased its lipid synthesis capacity in culture under T = 20 °C. The predominant lipid fraction was glycolipids and sphingolipids (G + S) for P. costavermella VAS2.5 and N. pyriformis PAT2.7 at 25 °C and M. gaditana VON5.3 at 20 °C, while neutral lipids (N) were higher in the other cultures. P. costavermella VAS2.5 and M. gaditana VON5.3 proved attractive ^Δ5,8,11,14,17^C20:5 producers. Furthermore, the content in ^Δ5,8,11,14,17^C20:5 regarding M. gaditana VON5.3 further increased when cultured under T = 20 °C. Also, N. pyriformis PAT2.7 stood out as a very promising candidate for ^Δ9^C16:1 production, which was present mostly in the N fraction. On the other hand, polysaccharide content seemed to not be affected by the culture temperature, while the high protein content at low incubation temperatures (e.g., T = 20 °C) is an indicator of the microalga’s good adaptation to these conditions and suggests its suitability for large-scale outdoor cultivation where temperatures are low during the winter months. Taking into account the findings of the present research, it is evident that all strains under the proper culture conditions can be exploited on a commercial scale, mostly due to their remarkable lipid profiles and protein content, especially in the aquaculture industry. At the same time, the lipid content of N. pyriformis makes it an alternative candidate for the biodiesel industry.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Chapman R.L. Algae: The world’s most important “plants”-an introduction Mitig. Adapt. Strateg. Glob. Change 20131851210.1007/s 11027-010-9255-9 · doi ↗
- 2Raven J.A. The possible roles of algae in restricting the increase in atmospheric CO 2 and global temperature Eur. J. Phycol.20175250652210.1080/09670262.2017.1362593 · doi ↗
- 3Jassey V.E.J. Walcker R. Kardol P. Geisen S. Heger T. Lamentowicz M. Hamard S. Lara E. Contribution of soil algae to the global carbon cycle New Phytol.2022234647610.1111/nph.1795035103312 · doi ↗ · pubmed ↗
- 4Bellou S. Baeshen M.N. Elazzazy A.M. Aggeli D. Sayegh F. Aggelis G. Microalgal lipids biochemistry and biotechnological perspectives Biotechnol. Adv.2014321476149310.1016/j.biotechadv.2014.10.00325449285 · doi ↗ · pubmed ↗
- 5Korozi E. Tsagou V. Kefalogianni I. Markou G. Antonopoulos D. Chakalis L. Kotzamanis Y. Chatzipavlidis I. Continuous culture of Auxenochlorella protothecoides on biodiesel derived glycerol under mixotrophic and heterotrophic conditions: Growth parameters and biochemical composition Microorganisms 20221054110.3390/microorganisms 1003054135336116 PMC 8953379 · doi ↗ · pubmed ↗
- 6Chew K.W. Yap J.Y. Show P.L. Suan N.H. Juan J.C. Ling T.C. Lee D.J. Chang J.S. Microalgae biorefinery: High value products perspectives Bioresour. Technol.2017229536210.1016/j.biortech.2017.01.00628107722 · doi ↗ · pubmed ↗
- 7Chisti Y. Biodiesel from microalgae Biotechnol. Adv.20072529430610.1016/j.biotechadv.2007.02.00117350212 · doi ↗ · pubmed ↗
- 8Damiani M.C. Popovich C.A. Constenla D. Leonardi P.I. Lipid analysis in Haematococcus pluvialis to assess its potential use as a biodiesel feedstock Bioresour. Technol.20101013801380710.1016/j.biortech.2009.12.13620117928 · doi ↗ · pubmed ↗