Some Insights into the Inventiveness of Dinoflagellates: Coming Back to the Cell Biology of These Protists
Marie-Odile Soyer-Gobillard

TL;DR
This paper reviews the unique and diverse cell biology of dinoflagellates, highlighting their evolutionary innovations and significance in the protist kingdom.
Contribution
The paper provides a comprehensive overview of dinoflagellate cell biology, emphasizing their evolutionary novelty and diversity.
Findings
Dinoflagellates exhibit remarkable diversity in lifestyles and cellular structures.
They display innovative features in mitosis, meiosis, and other cellular processes.
The eyespot (ocelloid) is an evolutionary marker found in certain dinoflagellates.
Abstract
In this review dedicated to the great protistologist Edouard Chatton (1883–1947), I wanted to highlight the originality and remarkable diversity of some dinoflagellate protists through the lens of cell biology. Their fossilized traces date back to more than 538 million years (Phanerozoic eon). However, they may be much older because acritarchs from the (Meso) Proterozoic era (1500 million years ago) could be their most primitive ancestors. Here, I described several representative examples of the various lifestyles of free-living (the autotrophic thecate Prorocentrum micans Ehrenberg and the heterotrophic athecate Noctiluca scintillans McCartney and other “pseudo-noctilucidae”, as well as the thecate Crypthecodinium cohnii Biecheler) and of parasitic dinoflagellates (the mixotroph Syndinium Chatton). Then, I compared the different dinoflagellate mitotic systems and reported observations…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
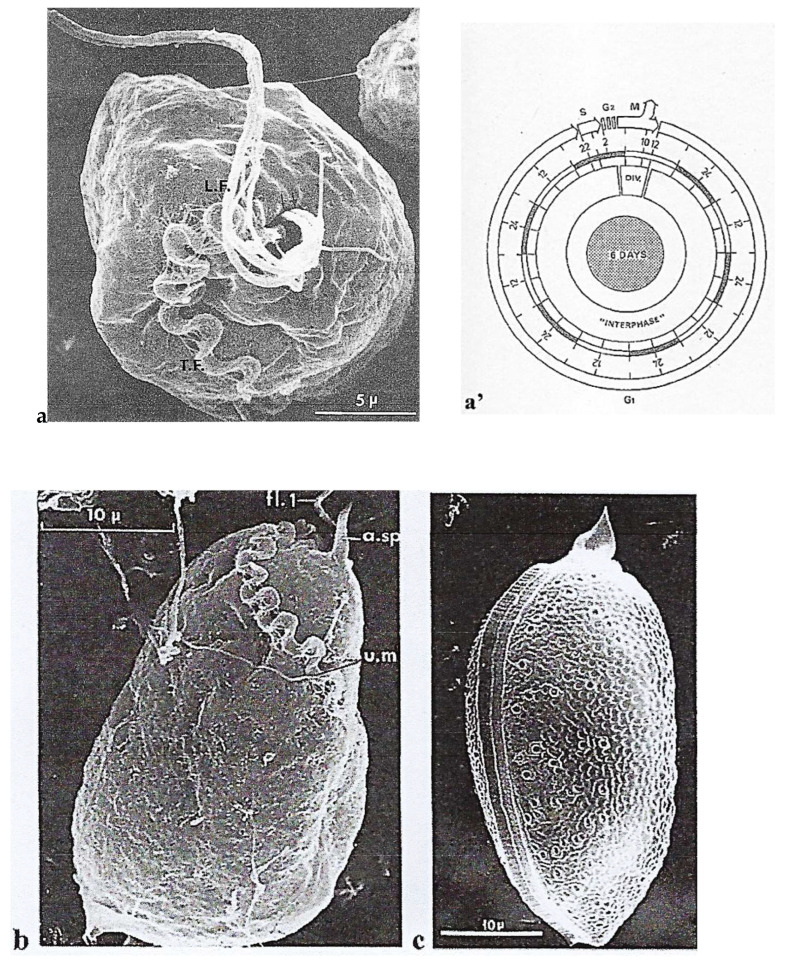 Figure 1
Figure 1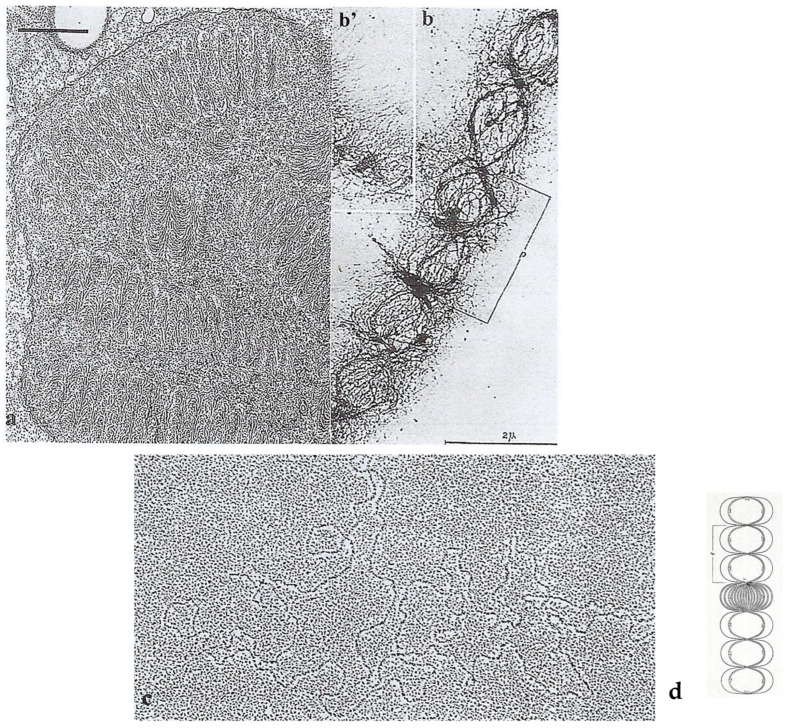 Figure 2
Figure 2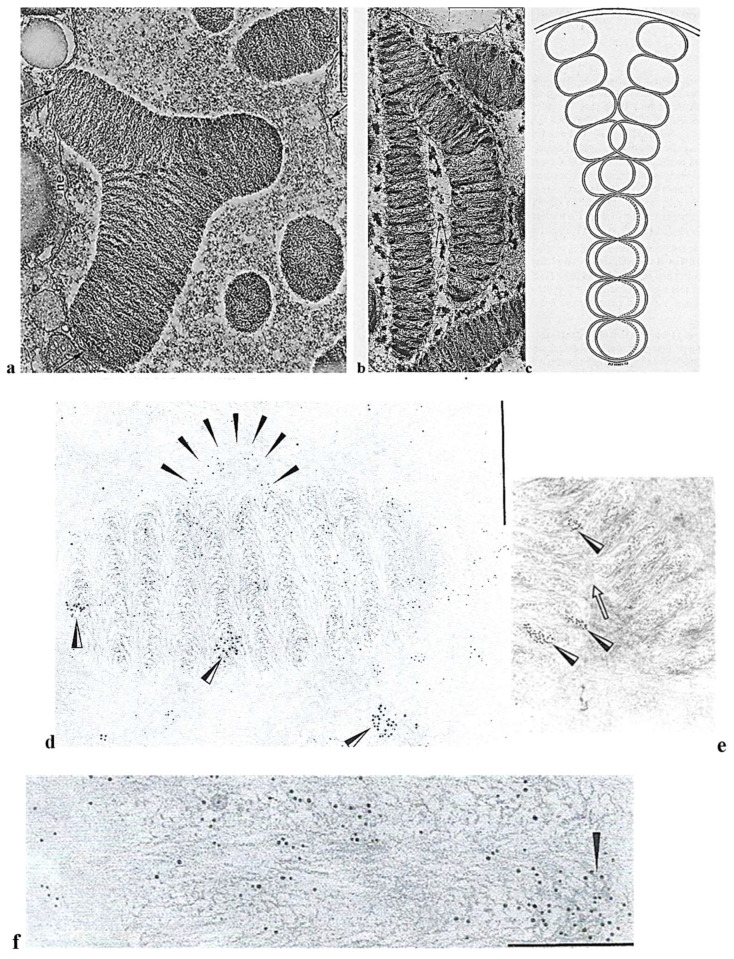 Figure 3
Figure 3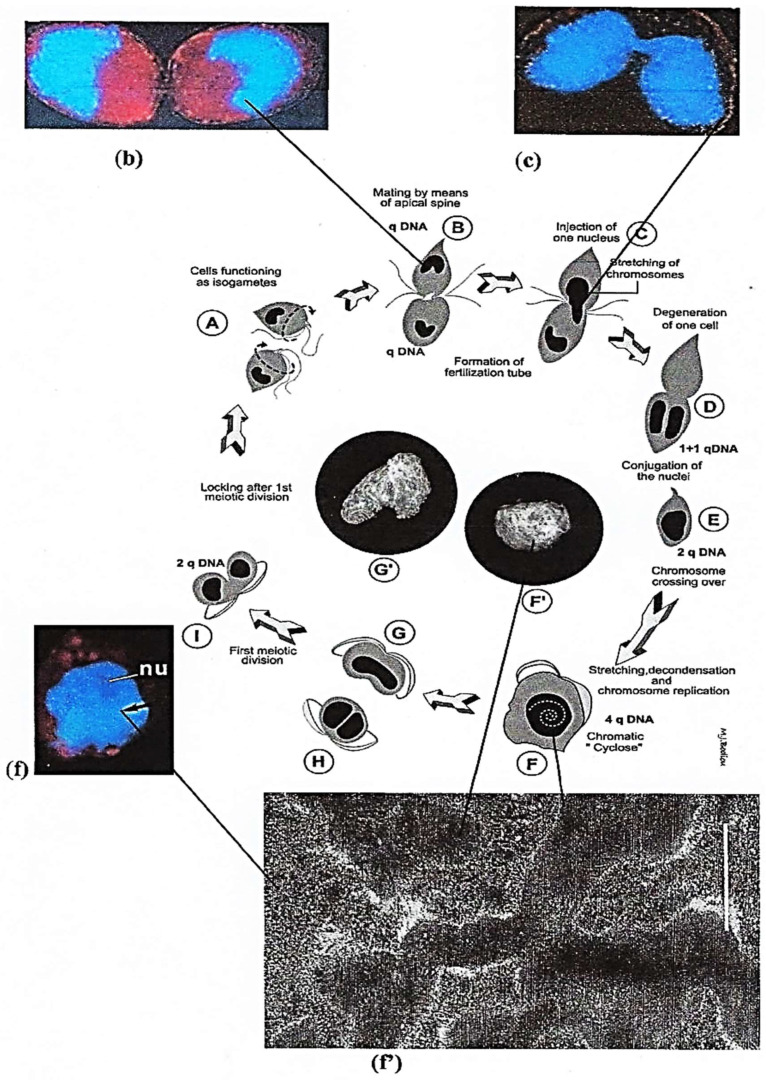 Figure 4
Figure 4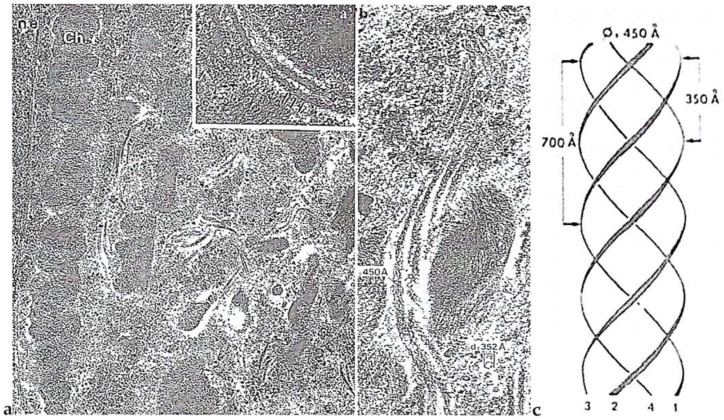 Figure 5
Figure 5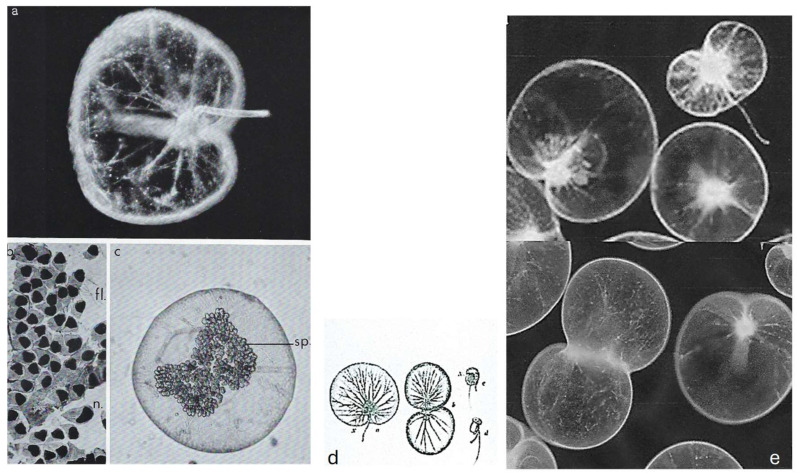 Figure 6
Figure 6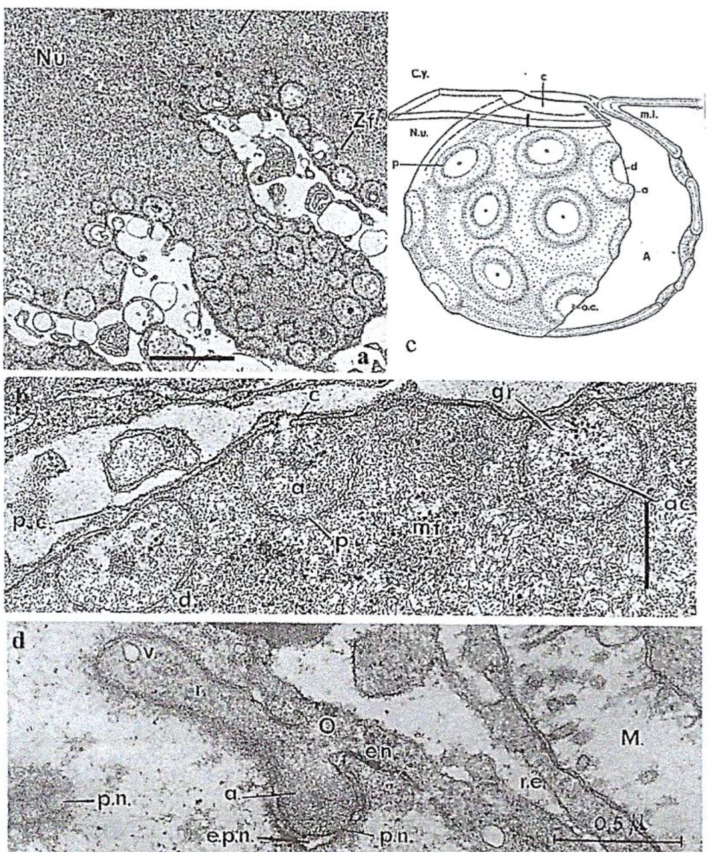 Figure 7
Figure 7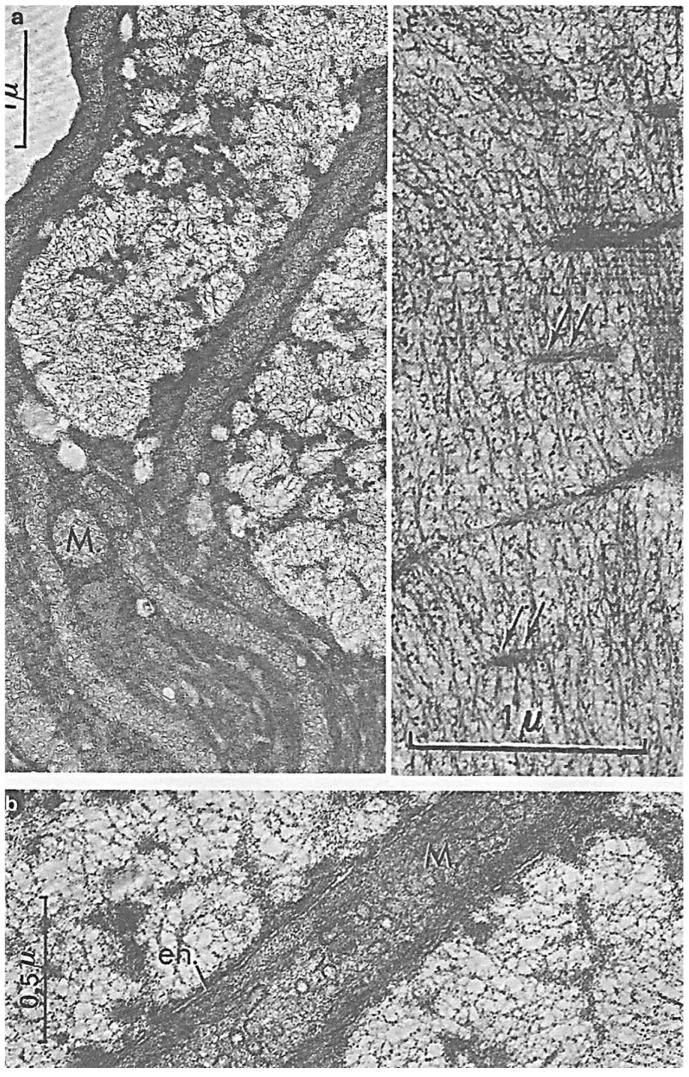 Figure 8
Figure 8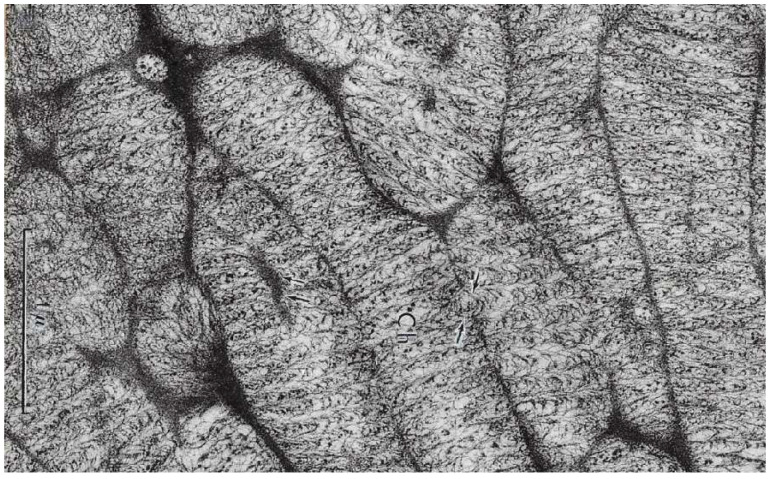 Figure 9
Figure 9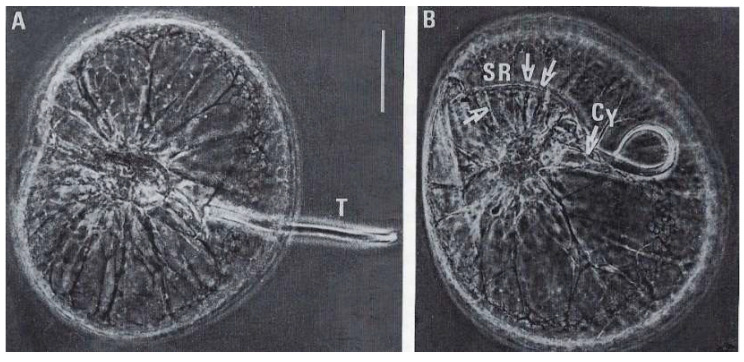 Figure 10
Figure 10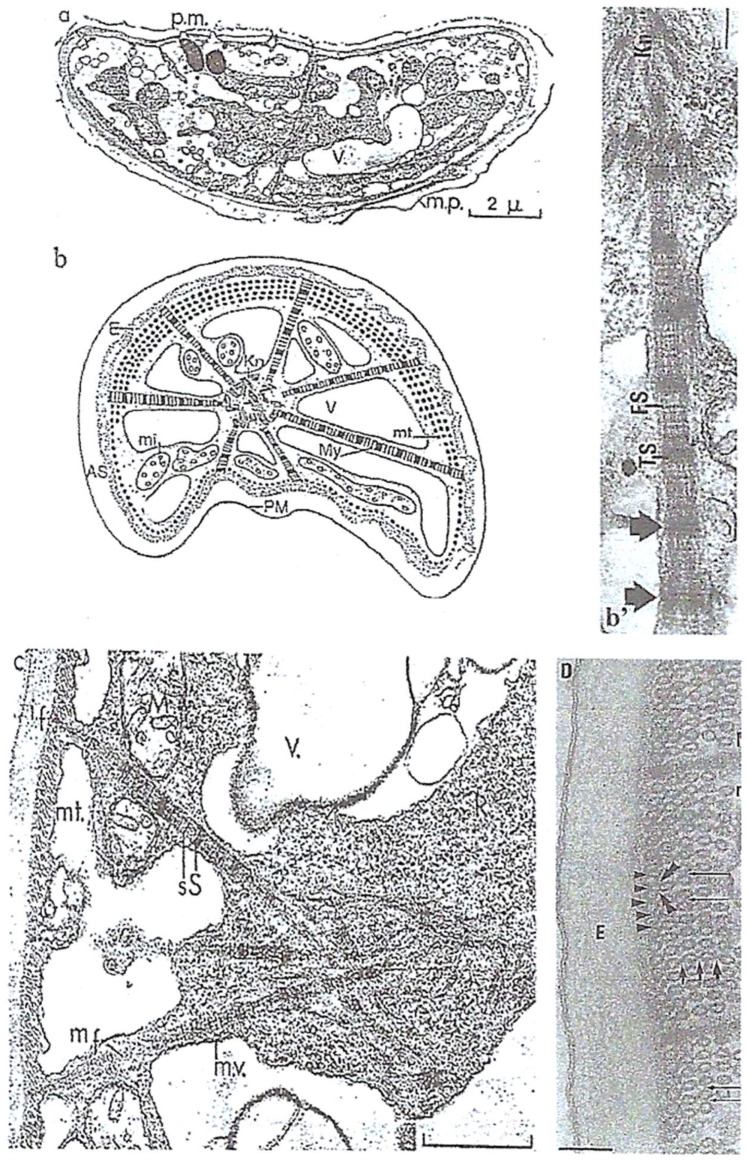 Figure 11
Figure 11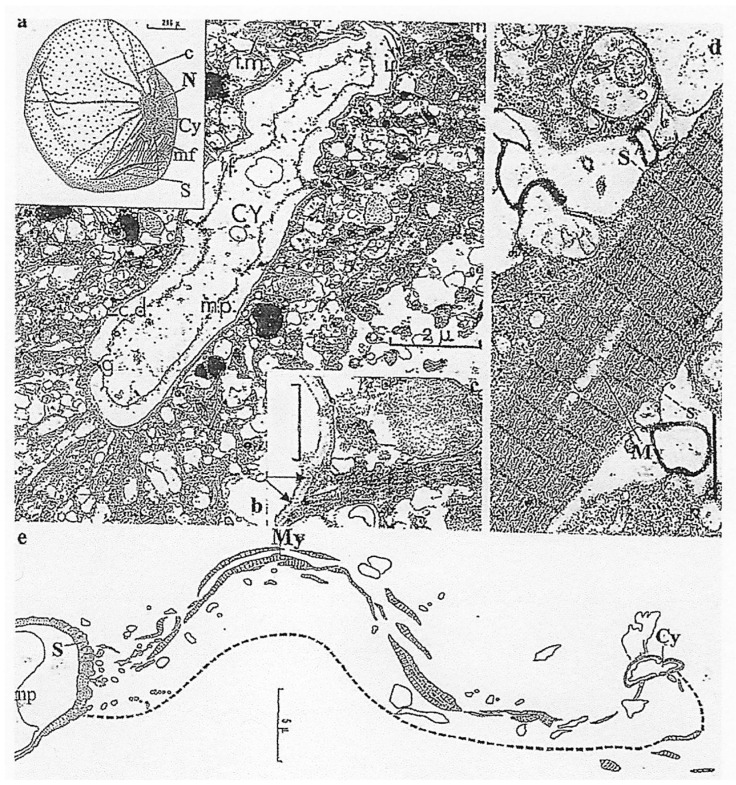 Figure 12
Figure 12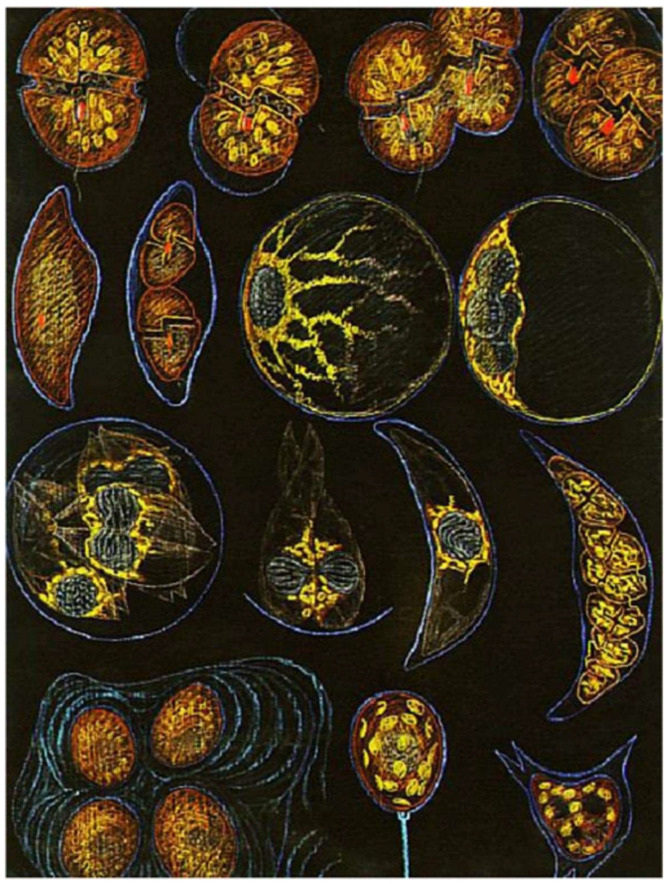 Figure 13
Figure 13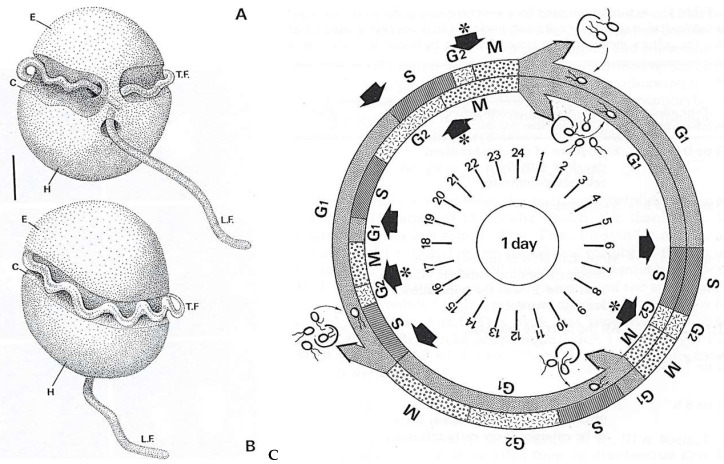 Figure 14
Figure 14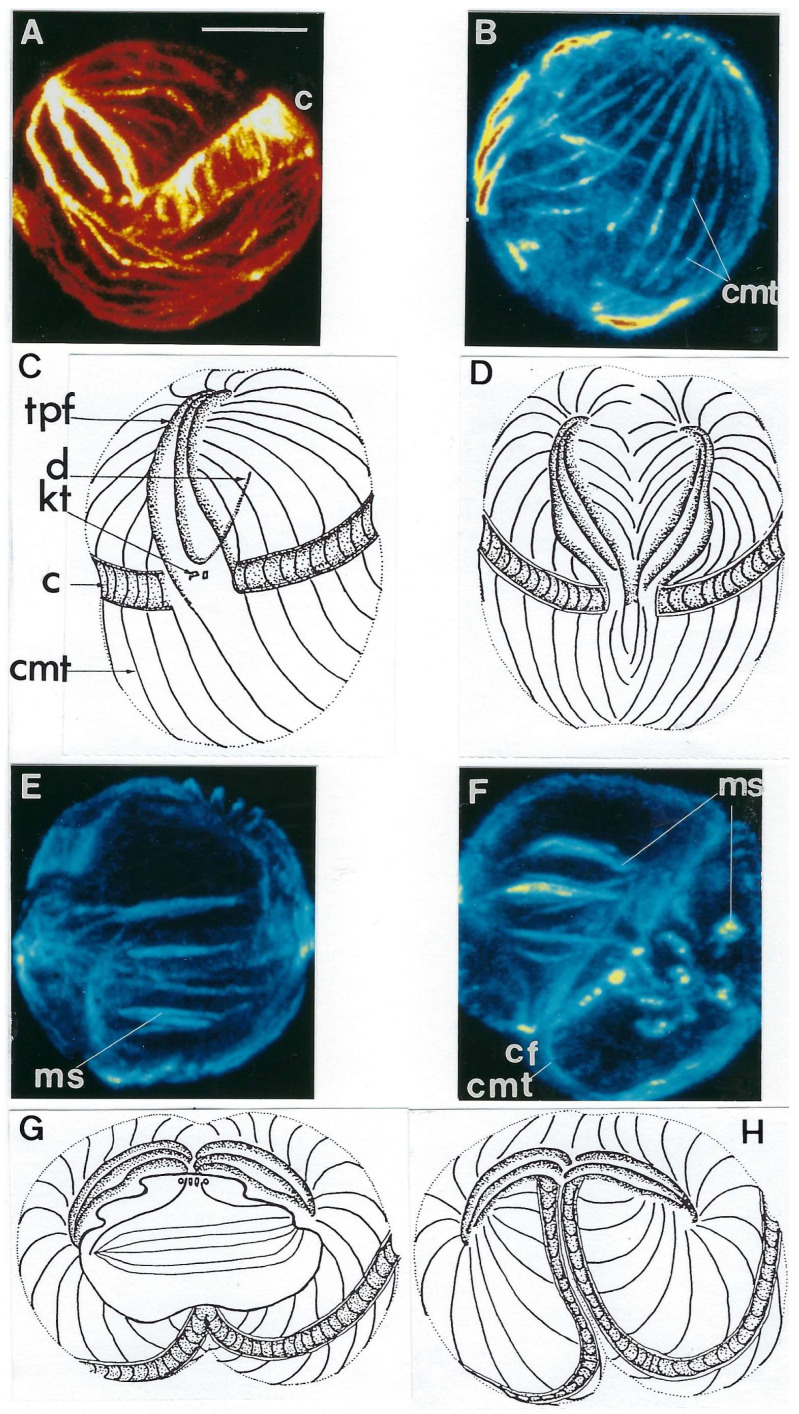 Figure 15
Figure 15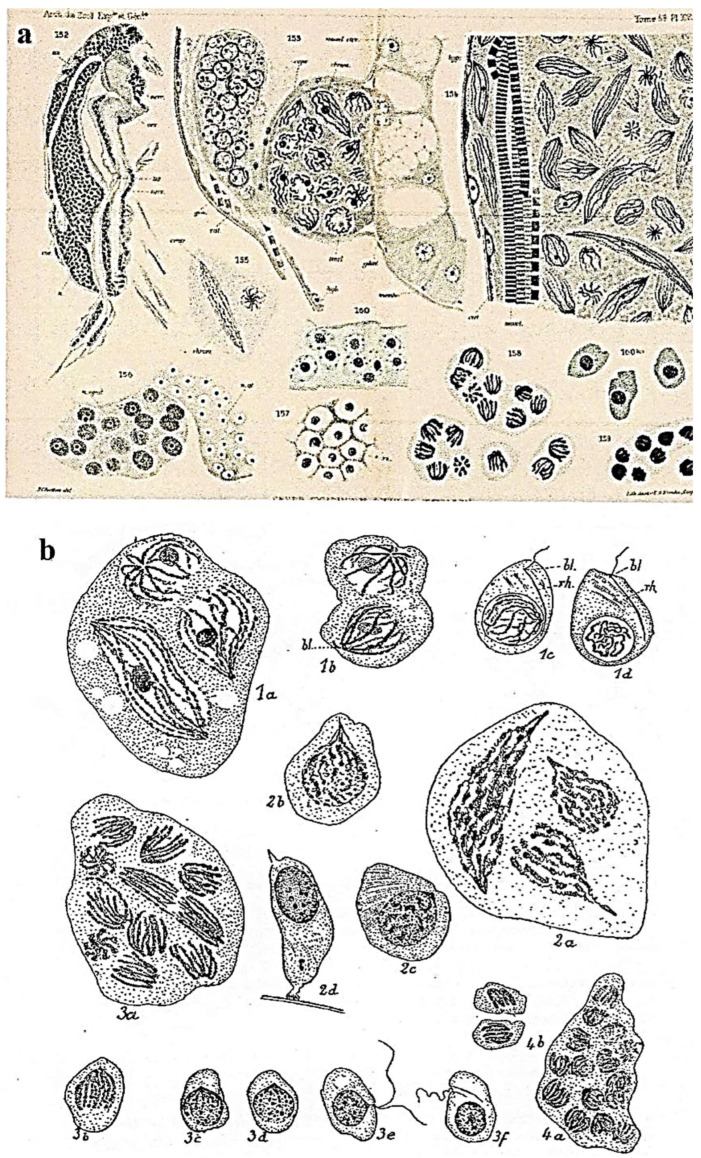 Figure 16
Figure 16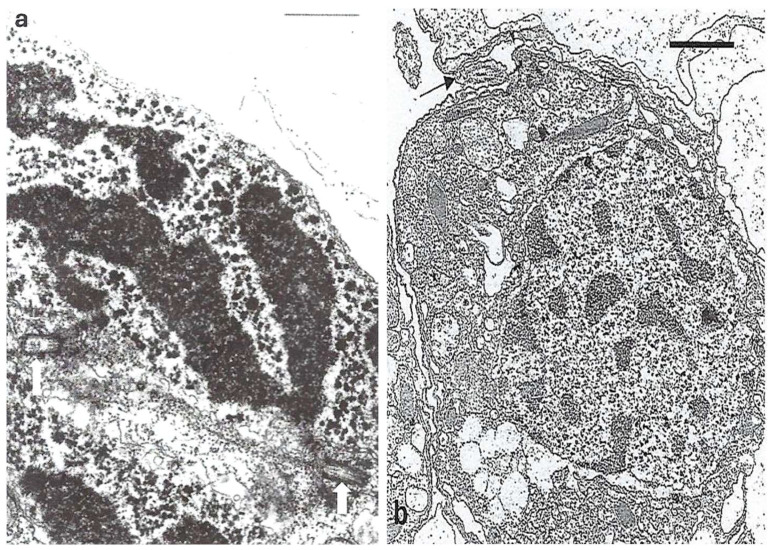 Figure 17
Figure 17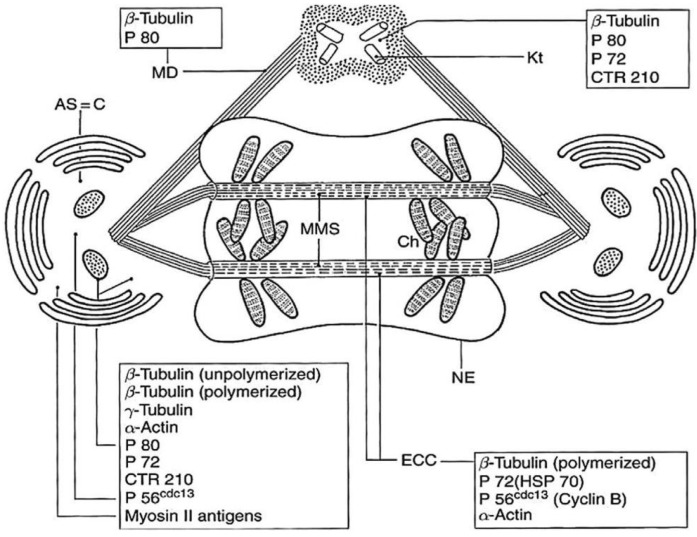 Figure 18
Figure 18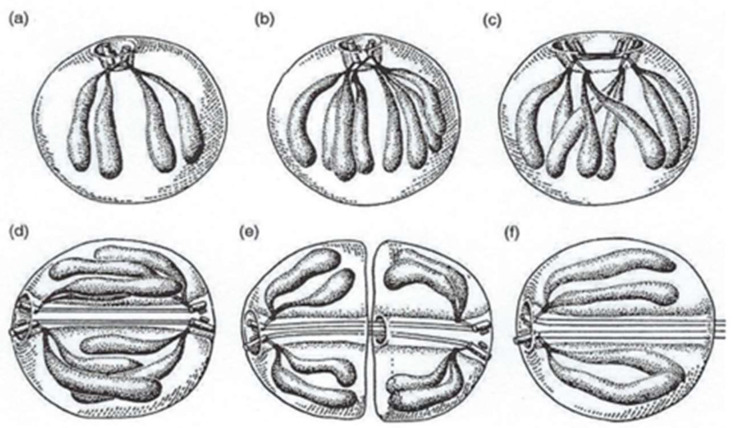 Figure 19
Figure 19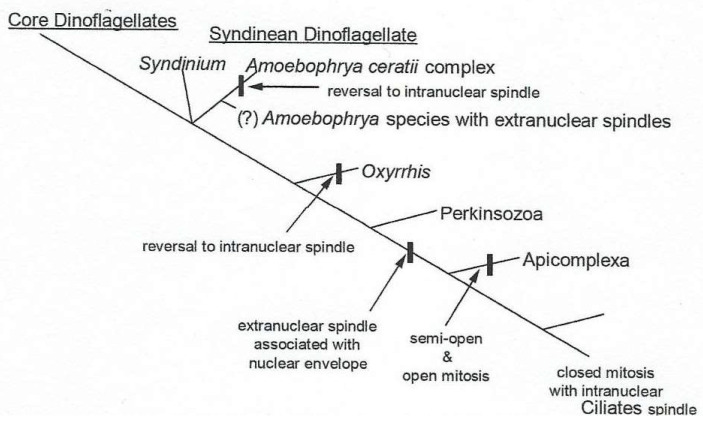 Figure 20
Figure 20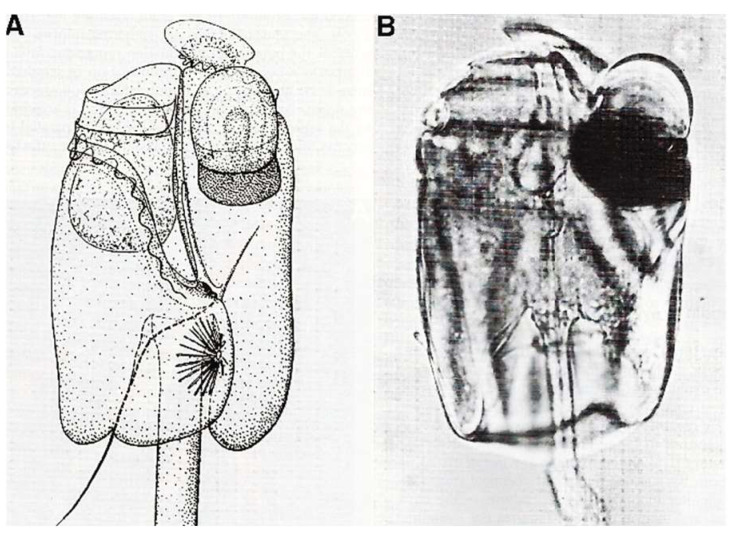 Figure 21
Figure 21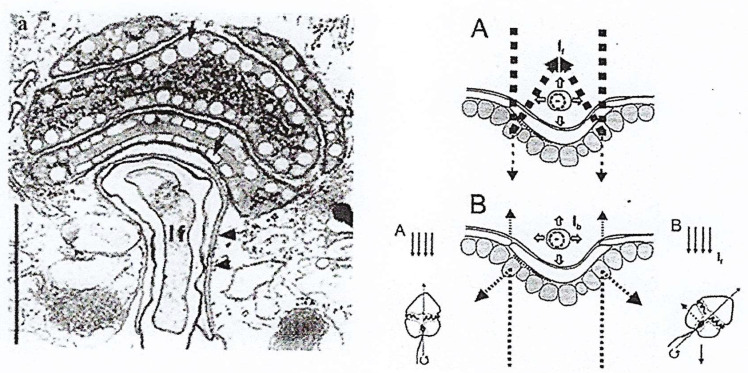 Figure 22
Figure 22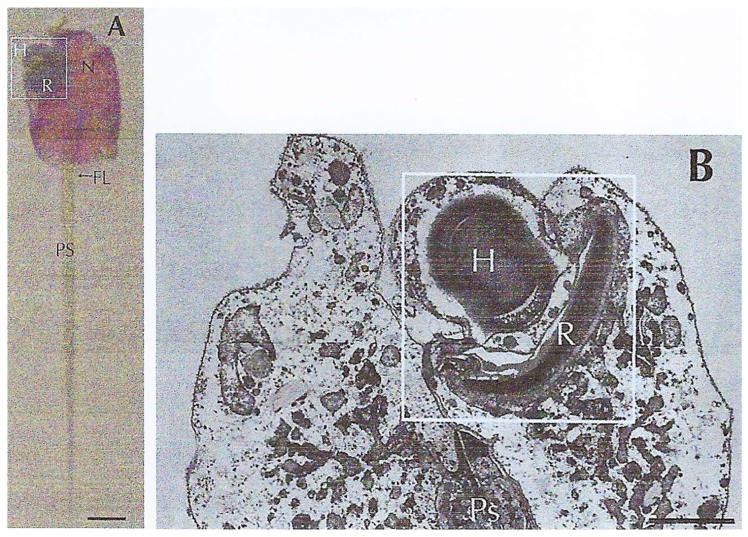 Figure 23
Figure 23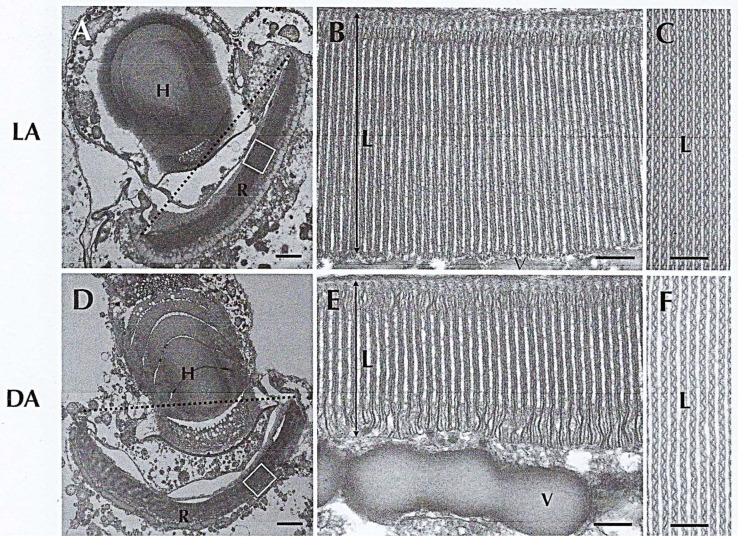 Figure 24
Figure 24Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsProtist diversity and phylogeny · Microbial Community Ecology and Physiology · Marine and coastal ecosystems
1. Preamble
This review is dedicated to the great protistologist Edouard Chatton (1883–1947), a pioneer in the study of the cytology of many (free-living and mixotrophic) dinoflagellates, who revealed the complexity and originality of these protists at the beginning of the 20th century. E. Chatton (1883–1947) was the first scientist to take a detailed interest in the description of the external morphology of many dinoflagellates and, especially, in their life cycle, cytology and mitosis (dinomitosis and syndinian mitosis). He masterfully described his findings in his PhD thesis, published more than one hundred years ago [1]. Chatton reported all his observations in his monumental Titres et Travaux Scientifiques (in English, Titles and Scientific Works) [2], a publication enriched by splendid course boards (160/110 cm), genuine artistic works he drew for his students, starting in 1920. A selection of the finest drawings by Chatton have been recently published in a book [3]. Chatton was also the first to distinguish between prokaryotic and eukaryotic microorganisms [4].
2. Introduction
Opening the chapter on the Phylum Dinoflagellates, written by FJR Taylor in the 1990s [5], is enough to appreciate the immense diversity of these protists. The current classification recognizes ~550 genera and a total of 6000 described species that play a prominent role in water ecology. Molecular clock and biogeochemical indices indicate that the dinoflagellate lineage diverged ~650 Ma, and fossil traces of thecate cells and of cysts/zygotes appeared during the Triassic [6]. Evidences of dinoflagellate existence come from the biogeochemical analysis of early Cambrian sediments (~520 million years ago), in which dinoflagellate-specific sterols (i.e., dinosterols) were detected in rock extracts and petroleum. However, some scholars think that they may have an older, earlier Precambrian origin, forming part of the Acritarchs [6,7]. According to B. Dale (2023) [8], the first dinoflagellates would have been naked and, therefore, without fossil traces except for biogeochemical residues. Only later they would acquire a protective theca to escape various predators. According to the phylogeny analyses by Bachvaroff et al. [9] and by Delwiche [10], they belong to the alveolates and can be autotrophic (about half are photosynthetic), heterotrophic, parasitic, and/or mixotrophic, symbiotic, plasmodial, or organized as “bi-nucleated” or “pseudo-multinucleated” cells. They all live in aquatic environments (seawater, brackish, or fresh water), and they have hemolymph, gut, or coelomic cavity for mixotrophic dinoflagellates [11]. The composition of their chromatin and the organization of their nucleus (dinokaryon) are also unique, as well as their “dinomitosis” [6,12,13]. In the nucleus of many dinoflagellate species, chromosomes are maintained in a quasi-permanent, compacted state. Molecular studies to estimate their diversity and to attempt their classification were carried out, especially to locate dinoflagellates among other protists and to classify dinoflagellate taxa [14]. Phylogenetic trees were built using protein sequences (e.g., dimeric iron-containing superoxide dismutases) [15] or the sequence of specific genes (e.g., 18S rRNA, 5.8S rRNA, 24S rRNA, or SS rRNA). All genomic data have been regrouped in the recent and very complete review by Serjie Lin [16].
In this review, I chose to present several examples of different categories of these original protists, from a non-exhaustive list of representatives of their different lifestyles, in order to highlight their remarkable diversity and adaptability, but without classifying them from the simplest to the highest evolutive complexity. I then focused on some innovative features, several of which were previously but partially described [6] using cell biology tools. I particularly highlight the acquisition of an eyespot (ocelloid) in several species belonging to the Warnowiidae family.
3. Materials and Methods
A review of the literature was carried out by searching the PubMed and Google Scholar databases from 2000 to 2024 following the preferred reporting items for systematic reviews and meta-analyses (PRISMA) guidelines, except for pioneering references (from 1920 to 2000). Studies were identified using the following keywords: dinoflagellates, eyespot, mitotic apparatus, Prorocentrum, Noctiluca, Crypthecodinium, Syndinium. Many illustrations in this review were taken from the selected publications, after having obtained permission from the publishers. For unpublished microphotographs taken by the author, the preparation/analysis methods were described in [17,18].
4. Results
This review summarizes the amazing and innovative diversity of dinoflagellates (autotrophic, heterotrophic, or mixotrophic) and their adaptations.
4.1. An Autotrophic Free-Living Dinoflagellate: Prorocentrum micans Ehrenberg
4.1.1. General Features
The cell cycle of P. micans Ehrenberg vegetative cells (Figure 1a’) was deciphered by Bhaud and Soyer-Gobillard [19], and it lasts 6 days, which is a relatively long cell cycle. This protist is characterized by an original flagellar system: an undulating membrane attached to the polysaccharidic epitheca that ends in a short transverse flagellum and a longitudinal flagellum (Figure 1a–c).
4.1.2. Original Features
Nucleus
The nucleus is filled with a hundred large chromosomes (10 µm long and 1 µm wide) (Figure 2a) and surrounded by large plastids, not visible in this nucleus picture. As these chromosomes are quite large, Haapala and Soyer-(Gobillard) could observe them using transmission electron microscopy (TEM) for the first time after spreading them on water (Figure 2b). This allowed for describing their organization, particularly the superhelical twisted structure of their nucleofilaments [22]. The twisted and regular unwinding of these chromosomes led to the hypothesis that they are organized in multiple circular chromatids (Figure 3c) [22,23]. This hypothesis was later confirmed and thoroughly described by Oakley and Dodge [24]. The isolation of chromatids from purified DNA molecules, spread and shadowed with platinum carbon, was particularly difficult to achieve [25] (Figure 2c).
P. micans has two types of reproduction: a vegetative reproduction, carried out by binary fission, and a sexual one. In a vegetative cell, dividing chromosomes must be attached to the internal part of the nuclear envelope (Figure 3a,b) and are not directly in contact with the microtubular spindle located in cytoplasmic channels that pass through the mitotic nucleus. This observation is supported by the absence of centromeric heterochromatin in P. micans [26]. In these rod-shaped chromosomes, centromeres could be localized at the chromosome tip and can be considered to be (virtual) telomeres. However, due to their lack of centromeric heterochromatin, P. micans could be considered as a kind of “immortal cell”, unable to undergo apoptosis.
P. micans chromosomes are devoid of histones and nucleosomes but are rich in 5-hydroxymethyluracil [27]. However, nucleosomal structures could be easily reconstructed in vitro using P. micans DNA and purified corn histones, showing that the presence of this abnormal base is not an impediment to their reconstruction [28]. P. micans also allowed us to carry out some of the first phylogenetic molecular analyses using ribonucleoprotein subunit (SSU, LSU) sequencing data [29].
Interestingly, using immunogold electron microscopy after sample preparation by vitrification at the liquid helium temperature (−269 °C) [17], a technique developed by Professor Jacques Dubochet (Nobel Prize in Chemistry) that preserves the antigenic sites particularly well, we could detect two DNA types in P. micans chromosomes: B-DNA (right-handed DNA) and the so-called “mirror” Z-DNA (left-handed DNA). B-DNA represents the usual DNA conformation (right-handed double helix). It is the most represented in the living world (Figure 3f), unlike Z-DNA (which turns left). Thanks to work in collaboration with Dr Etienne Delain’s group at the Gustave Roussy Cancer Research Institute in Villejuif (France), who produced an anti-Z-DNA antibody from urine samples of people with cancer [18], we could detect the presence of Z-DNA as clusters of gold particles at the chromosome periphery (Figure 3f) and in the chromosomal fission zones (Figure 3e). Conversely, B-DNA was localized in the chromosome body (Figure 3d,f). We hypothesized that in these always compacted chromosomes, transcription takes place at the chromosome periphery and DNA opening occurs through a loop opening process dependent on the mirror Z-DNA [18].
Sexual Reproduction
P. micans sexual reproduction was observed in our laboratory by chance, after a glass bottle with some cultured cells was placed in a refrigerator at 4° in the dark for 12 h. The next morning, many vegetative cells (n = 1qDNA) had paired up, clinging to one another through their apical spine and emitting a connecting tube (i.e., fertilization tube) (Figure 4A–C) through which one cell (male?) injects its genetic material surrounded with nuclear envelope into the other (the female?) (Figure 4D,E) [29]. Shortly after the fusion of the two nuclear envelopes and nuclei, their chromatin undergoes a very impressive circular and rapid movement on the right named chromatic cyclosis as observed with phase contrast optical microscope for several minutes, during which the male and female genetic materials mingle and chromosomes completely lose their regular helical structure (Figure 4F,f,f’). By quantifying DNA in single cells, we showed that in P. micans, early planozygotes (Figure 4f) (n = 2qDNA content) double their DNA to n = 4qDNA before the first of the two zygotic divisions (meiosis), which leads to n = 1qDNA in vegetative cells. We also observed that nuclear chromatic cyclosis occurred just before the first meiotic division and that the chromosome nucleofilament organization was altered [21]. Nuclear chromatic cyclosis was discovered in two Ceratium species by Pouchet in 1885 [30] and thoroughly described in six Peridinium species by Biecheler in 1935 [31]. Many other dinoflagellates present both vegetative (binary reproduction) and sexual reproduction, the last occurring often when the physicochemical conditions are unfavorable.
Intriguing Intranuclear Supercoiled Microcables During Meiosis [32]
During meiosis, TEM observations showed a total disorganization of the chromosomal nucleofilaments. Specifically, the chromosomes are very uncoiled but still well individualized (Figure 4f’) [29] to allow for intergenomic exchanges. During this period, chromatic cyclosis occurs, and microcables are visible in the nucleoplasm from TEM observations (Figure 5a) [32]. They are composed of twisted filaments organized in a tetrahelix in which the unitary microfilaments have a diameter of 60 Å. They are organized in a right tetrahelix with a diameter that varies from 35 to 45 nm (Figure 5a’,b,c). The unit microfilament diameter corresponds to that of actin that we detected and described for the first time in a dinoflagellate [33]. The presence of actin was confirmed by Berdieva et al. (2018) [34] who demonstrated the presence of F-actin in the cytoplasm and G-actin in the nucleus at the nucleolar level in the dinoflagellate Prorocentrum minimum (Pavillard) Schiller. This evidence allowed us to hypothesize that microcables could be the driving element of chromatid mixing during chromatic cyclosis as well as of chromosome transport during dinomitosis.
4.2. Two Heterotrophic Free-Living Dinoflagellates: Noctiluca Scintillans Mc Cartney and Crypthecodinium cohnii Biecheler
4.2.1. Noctiluca Scintillans McCartney
General Features
Commonly called “Fire of the Sea”, N. scintillans McCartney is a non-parasitic athecate free-living spherical protist (Figure 6a–c) without plastids that lives in marine environments. It exhibits bioluminescence when agitated, through a luciferin-luciferase system in its cytoplasm [35,36], like some other dinoflagellate species. These planktonic dinoflagellates, which have not been recorded to produce toxins, can bloom into non-toxic “red tides” that spread as a several-centimeter-thick layer on the sea surface. N. scintillans McCartney does not have cellulose thecal plates, but it is not completely “naked”. Indeed, its cell covering or “amphiesma” presents an outer membrane that surrounds the cell, amphiesmal vesicles, and a more or less thin pellicular layer, the epiplasm, as described by Melkonian and Höhfeld [37]. N. scintillans McCartney can divide by binary fission after loss of the tentacle or by sporulation after sexual reproduction [38] to form sporocytes (Figure 6a–c) and then uniflagellate spores, which will ensure the dissemination and resistance to growth-unfavorable conditions (Figure 6b) [39,40,41]. During sporogenesis, their chromatin progressively condenses as the divisions progress [42]. Chromosomes are well individualized only in mature spores where there are at least 120 chromosomes per nucleus.
Original Features
In the trophozoite and at the start of divisions, the inner part of the nuclear envelope is lined in a peripheral fibrous zone (Figure 7a,b) with nuclear ampullae that bear nuclear pores and actively participate in sporogenesis preparation (Figure 7a–d). After the calculation, the surface area of the ampullae is twice the surface area of the total nuclear envelope [40]. Trophozoites can undergo a binary division or form spores. In a very short time, nuclear ampullae constitute nuclear membrane reserves immediately available during sporulation, accelerating the divisions during a bloom (red tide). Indeed, during the beginning of sporulation, nuclear ampullae fuse to leave a smooth nuclear envelope devoid of ampullae in the spores (Figure 8b) [40,41]. Sporulation is triggered by physicochemical reasons not yet specified in the surrounding environment, and then, multiplication is very rapid by splitting of the chromatic mass, toward stage 16 nuclei during which chromatin condenses and individualizes chromosomes (Figure 9).
Cytochemical investigations did not detect histone-like proteins in the trophozoite nucleus. At the molecular level, the unusual 5-hydroxymethyluracil base has been found in the nuclear DNA of N. scintillans [27]. In the investigated dinoflagellate species, this unusual base replaces 12–68% of all thymine residues, a relatively important replacement amount.
Cytoskeletal elements that participate in N. scintillans trophozoite motility are also involved in nutritional functions. They are located in the tentacle (Figure 10A,B) and at the level of the oral apparatus (cytostome) where cytoplasmic myofibrils are organized in striated and contractile fibers named “myonemes” (Figure 11a,b,b’). We were the first to observe these striated fibrillar structures in N. scintillans, which are similar to the striated myonemes in its contractile tentacle [39,43]. Myonemes are inserted on the epiplasmic membrane (E), equipped with a large number of microtubule rows (Figure 11D, arrowheads) that are crosslinked with each other (Figure 11D, arrows) [39]. Striated myonemes are distributed along the tentacle and linked to one another by an axial knot (Figure 11b,c). Tentacle contraction involves epiplasmic membrane deformation, myoneme contractility, and microtubule modifications [43].
The oral apparatus (cytostome) opening and closing (Figure 12a–c) during prey capture is ensured by a complex system of curtains of striated (contractile) myofibers (Figure 12d) that run between the cytostome and are anchored at a reinforced furrow or “sulcus” (schematized in Figure 12a,e) [39]. Preys include pollen grains, other protists, and sometimes even congeners. It is amusing to note that this is one of the first recorded cases of cannibalism in the Kingdom Protista.
Phylogenetic analyses of this complex cell, using the gene sequences of beta-tubulin, HSP90 [44] or nuclear 28S rDNA [45], support its placement among dinoflagellates, not far from Oxyrrhis marina Dujardin. These species do not have a theca and probably appeared very early in the Precambrian era, according to Dale [8].
4.2.2. Other Free-Living Bioluminescent Dinoflagellates: An Homage to Edouard Chatton’s Scientific and Artistic Talents [3].
E. Chatton (1930) represented these species on a magnificent course board (160/110 cm) drawn with colored pastels on black paper (Figure 13), for his students [3].
4.2.3. Crypthecodinium cohnii Biecheler
General Features
The heterotrophic dinoflagellate C. cohnii Biecheler is a particularly fascinating protist despite its classical biflagellate morphology (Figure 12a,b). Kubai and Ris described its mitosis for the first time [46].
Innovative Features
C. cohnii presents a complex cell cycle [47]. One vegetative cell can perform two successive complete cell cycles (16 h), as shown in Figure 14C, and release four daughter cells. One of these new swimming cells releases two daughter cells 10 h later (external circle of the diagram). During this time, other swimming cells could produce two or four daughter cells. Different diagrams could be possible with different cycle lengths and numbers of daughter cells. Immunocytochemistry and confocal microscopy analyses of the microtubular cortex and mitotic apparatus [48,49,50] showed a well-developed microtubular cytoskeleton (Figure 15). Interestingly, we have shown that a HSP70-related protein is associated with the centrosome and is conserved from C. cohnii to human cells [51]. C. cohnii is rich in lipids [52], and among the heterotrophic marine dinoflagellates, is a prolific producer of docosahexaenoic acid (DHA), an important fatty acid [53]. Amusingly, after the publication of the study on the complex C. cohnii cell cycle by our laboratory [47], we were contacted on several occasions to give advice concerning the specific culture of this species by industrial groups in the USA working on the elaboration of artificial “maternal” milk or of DHA production.
4.3. A Mixotrophic Dinoflagellate: Syndinium spp. Chatton
4.3.1. General Features
According to the first descriptions by Chatton (1910), the main hosts of this parasitic plasmodial dinoflagellate are the pelagic copepods of the Mediterranean Sea close to Banyuls-sur-Mer, France [1,2]. The “plasmodium”, which is composed of a cytoplasmic mass with numerous nuclei containing five V-shaped chromosomes and without cellular partitioning, is constantly in mitosis, growing in the general cavity (coelomic cavity) of copepods and other crustaceans and rapidly destroying all their vital organs. The mature biflagellate dinospores, totally devoid of plastids, are released into the seawater and very rapidly swim to quickly parasitize another prey.
4.3.2. Innovative Features
Like Hematodinium perezi (Syndiniales) that was discovered by Chatton and Poisson [54] on the French coasts and parasitizes the blood of crabs, Syndinium sp. can also parasitize the general cavity of many crustaceans, especially copepods. Currently, these Syndiniales are widespread throughout the world, and many scientists have been working to refine their molecular characterization [55,56] in connection with important economic issues because they often infest edible crustaceans. Their destructive action due to their rapid multiplication can cause the death of many crustacean species in a short time, and their power of contamination by dinospores (totally devoid of plastids) is infinite.
Chatton considered syndinian mitosis as a particular mitosis in dinoflagellates [57] and described it in detail (Figure 16a,b) [1,2] in several different species that parasitize various copepod crustaceans or radiolarians. Subsequently, TEM observations by Ris and Kubai (1974) showed the originality of this mitotic system [58]. The compacted chromosomes are attached to the extranuclear microtubular mitotic spindle through the nuclear membrane by means of kinetochores via a large cytoplasmic channel [58,59]. They are connected to the centrosome region that contains two centrioles (Figure 17a) and to microtubules of the mitotic spindle. It is important to note that kinetochores appeared for the first time within dinoflagellates in the kingdom Protista. Later, when mature dinospores totally devoid of plastids are released into seawater (Figure 17b), their chromatin is totally fragmented.
In his different descriptions of Syndinium sp., Chatton observed three different spore types, with different sizes (Figure 15B) [2], that parasitize two different copepod species (Clausocalanus arcuicornis and Paracalanus parvus) and wondered about their roles. Later, Skovgaard et al. [55], using full-length SSU rDNA sequences of Syndinium turbo from these two copepod hosts, concluded that the three spore morphotypes are 100% identical and belong to a single species, Syndinium turbo Chatton. Moreover, phylogenetic analyses place Syndinium as a sister taxon of Hematodinium sp., a blue crab parasite according to Chatton and Poisson. The main innovations in Syndinium sp. essentially concern motility, nutrition, and reproduction (sexual or not), and also the appearance of histone-like basic nuclear proteins linked to their DNA [58,59], as well as the presence of kinetochores during mitosis [58].
5. Dinoflagellate Mitotic Apparatus as an Evolutionary Marker
Despite the great dinoflagellate diversity in terms of innovations, physiology, lifestyle, and cell cycle, they have a remarkably homogeneous mitosis mechanism, except for Syndinium spp., Oxyrrhis marina, and some Amoebophrya species. The system of cytoplasmic channels passing through the intact nucleus indicates that microtubules are never in direct contact with the chromosomes but are always separated from them by the persistent nuclear envelope, as shown in the “models” C. cohnii Biecheler and P. micans Ehrenberg (Figure 18) [49].
It should be noted that the kinetochores (contact between chromosomes and mitotic spindle) and the polarized centrosomes, which contain two centrioles, were first observed in Syndinidae. This kind of mitosis has been particularly well studied by Ris and Kubai (Figure 19a–f) [58].
From all the precedent data and their own findings in Amoebophrya spp., Moon et al. (2015) observed that not all species classified as dinoflagellates have an extranuclear spindle [60]. For instance, in some Amoebophrya spp. species, an extranuclear microtubule cylinder, located in a depression of the nuclear surface during the interphase, moves into the nucleoplasm via sequential membrane fusion events and develops into an entirely intranuclear spindle [60]. Their results suggest that the intranuclear spindle of Amoebophrya spp. may have evolved from an ancestral extranuclear spindle, as shown on the phylogenetic tree of different mitotic apparatuses in Figure 20.
6. A Rather Perfected System: The Eyespot (Ocelloid) of Dinoflagellates
Surprisingly, the eyespot (ocelloid) is one of the most evolved photosensitive organelles in protists [5] and is considered an important phylogenetic marker in dinoflagellates. The ocelloid is present in several heterotrophic athecate dinoflagellates from the Warnowiaceae family, such as Nematodinium, Warnowia, Erythropsis, in several Woloszynskioids, and in the autotrophic Glenodinium (Peridinium) foliaceum Stein, a binucleate dinoflagellate, with both a dinokaryon and another nucleus probably of endosymbiotic (diatom) origin. Greuet (1965) published the first description of an eyespot in the dinoflagellate Erythropsis pavillardi Hertwig [61], as reported by Gehring (2001) [62] and then Francis (1967) in Nematodinium spp. [63]. Using TEM, Greuet showed that this most sophisticated structure is ~25 µm long and 15 µm wide [64,65,66]. Its main characteristic is the presence of a transparent and domed hyalosome, which plays the role of the lens, and of a pigment layer, which plays the role of the retina (Figure 21A,B). This complex photosystem was observed in many species of the family of heterotrophic Warnowiidae [67,68] and also in the binucleated dinoflagellate Glenodinium foliaceum Stein, now Kryptoperidinium triquetrum (Ehrenberg) U. Tillmann, M. Gottschling, M. Elbrächter, W.-H. Kusber, and M. Hoppenrath, 2019. Its light transmission mechanism was elucidated by Kreimer [69].
In 1999, Kreimer [69] described the eyespot of the binucleated G. foliaceum in which two DNA types from two nuclei (a dinokaryon and a nucleus of endosymbiotic origin of diatom type) and two chloroplast types of the same origin have been detected [70]. In this dinoflagellate, the eyespot is located in the posterior part of the cell close to the sulcus (Figure 22a). It is composed of a pigment cup, retinoid, and lens and functions as a photoreceptor through which light can pass and be reflected outside (Figure 22A), or it can pass through the protist body before being reflected back out (Figure 22B), which determines the orientation of the cell swimming.
The eyespot of Warnowiidae is located in the anterior part of the cell. In Erythropsidinium spp., a heterotrophic dinoflagellate with a posterior appendage called a piston that plays a role in its locomotion, the eyespot occupies a significant volume of the cell (Figure 23A). Walter Gehring^†^ (1939–2014), to whom I would like to pay special tribute here, worked on the genetic control of eye development and the evolution of eyes and photoreceptors in the animal kingdom [62,71]. From the work of Greuet [61,64,65,66], he thought that the dinoflagellate ocelloid represents an evolutionary enigma because it looks like a multicellular camera-type eye, but is found in a unicellular protist. Then, in 2015, Hayakawa et al., with Walter Gehring [72] used TEM to determine whether the dinoflagellate ocelloid is functionally photoreceptive. They found that this sophisticated structure is composed of a retina and lens-like structures called the retinal body and of a transparent hyalosome (the lens) (Figure 23A,B). Moreover, they observed that the retinal body changes its morphology depending on the illumination conditions and that the hyalosome displays a refractile nature (Figure 24). Lastly, they identified a rhodopsin gene fragment by in situ hybridization in Erythropsidinium expressed sequence tags (ESTs) that is expressed in the retinal body [72] and is most closely related to bacterial rhodopsin. Therefore, with Gavelis et al. [73], they could strongly affirm that “Eye-like ocelloids of dinoflagellates are built from different endosymbiotically acquired components”.
7. Conclusions
As reported by Hayakawa et al. (2014) [72], Darwin wrote in his work “On the Origin of Species” [74] that the eyes are an example of organs of extreme perfection and complication. Darwin was convinced that they had only appeared thanks to natural selection. In the case of the Erythropsidinium ocelloid, a highly elaborate camera-type eye that resembles part of the metazoan eye [72,73] has evolved in a single cell, a protist dinoflagellate, and is probably the vestige of endosymbiosis. Despite the great diversity of these protists in terms of taxomony and innovations and the fact that some specific proteins associated with the centrosome are conserved up to human cells (e.g., HSP70-related protein) [51], and the presence of right (B-) and left (Z-) handed DNA in their chromosomes [18], we observe the result of their evolution as it appears today, after more than 1500 million years since the beginning, probably dating back to the Proterozoic era according to Brian Dale [8]. Much research using the most advanced techniques in cell biology is still necessary to try to solve these enigmas.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Chatton E. Les Péridiniens Parasites: Morphologie, Reproduction, Ethologie Librairie, H. Le Soudier Paris, France 19201475
- 2Chatton E. Titres et Travaux Scientifiques (1906–1937)Imp-Editor Sottano Sète, France 19381405
- 3Soyer-Gobillard M.-O. Schrével J. The Discoveries and Artistic Talents of Edouard Chatton and André Lwoff, Famous Biologists 1st ed.Cambridge Scholars Publishing Cambridge, UK 20211228
- 4Chatton E. Pansporella perplexa Ann. Sci. Nat. Zool.19258185
- 5Taylor F.J.R. Phylum Dinoflagellata Handbook of Protoctista 1st ed. Margulis L. Corliss J. Melkonian M. Chapman D.J. Mc Khan H.I. Jones and Bartlett Publishers Boston, MA, USA 1990419437
- 6Riding J.B. Fensome R.A. Soyer-Gobillard M.-O. Medlin L.K. A Review of the Dinoflagellates and Their Evolution from Fossils to Modern J. Mar. Sci. Eng.202311110.3390/jmse 11010001 · doi ↗
- 7Moldowan J.M. Talyzina N.M. Biogeochemical evidence for dinoflagellate ancestors in the early Cambrian Science 19982811168117010.1126/science.281.5380.11689712575 · doi ↗ · pubmed ↗
- 8Dale B. Paleontological Evidence for Dinoflagellates and Ciliates as Early Eukaryotes J. Mar. Sci. Eng.20231153310.3390/jmse 11030533 · doi ↗