Cloning and Spatiotemporal Expression Analysis of IGF1R Gene cDNA in Alopex lagopus (Arctic Fox)
Wei Xu, Hualin Fu, Xiangyu Meng, Yiwen Sun, Fangyong Ning, Zhiheng Du

TL;DR
This study cloned and analyzed the IGF1R gene in Arctic foxes, revealing its sequence and expression patterns across tissues and ages.
Contribution
The study provides the first detailed cloning and spatiotemporal expression analysis of the IGF1R gene in Arctic foxes.
Findings
The Arctic fox IGF1R gene encodes a 538-amino-acid protein with high homology to the canine IGF1R gene.
IGF1R expression is highest in testis and increases in liver with age, suggesting roles in metabolism and development.
Phosphorylation sites and structural features of the IGF1R protein were identified, indicating functional relevance.
Abstract
This study aimed to clarify the sequence characteristics and spatiotemporal expression patterns of the insulin-like growth factor 1 receptor (IGF1R) gene in Alopex lagopus (Arctic fox), thereby addressing the existing knowledge gap regarding IGF1R-mediated growth regulation in this species. The findings establish a crucial foundation for subsequent investigations into the correlation between this gene and Arctic fox growth traits. Specific primers were designed based on the cDNA sequence of the canine IGF1R gene (Accession No. XM_545828). The full-length coding sequence (CDS) of the Arctic fox IGF1R gene (1617 bp, encoding 538 amino acids) was successfully cloned via RT-PCR. Phylogenetic analysis using the Unweighted Pair Group Method with Arithmetic Mean (UPGMA) algorithm revealed a 99% sequence homology in the IGF1R gene between the Arctic fox and canine, confirmed their closest…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
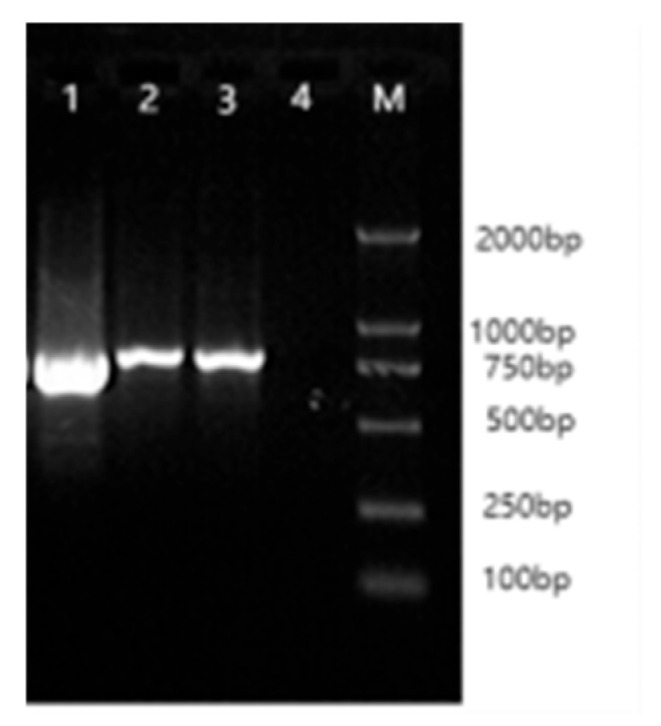 Figure 1
Figure 1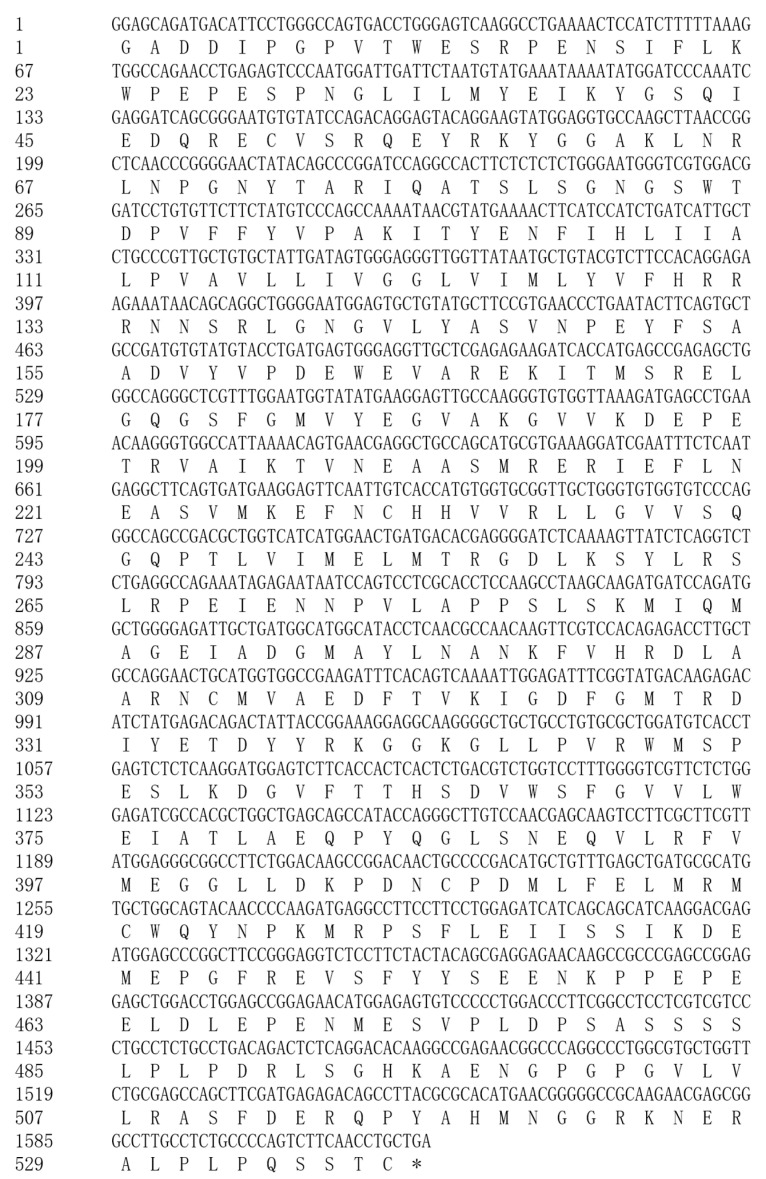 Figure 2
Figure 2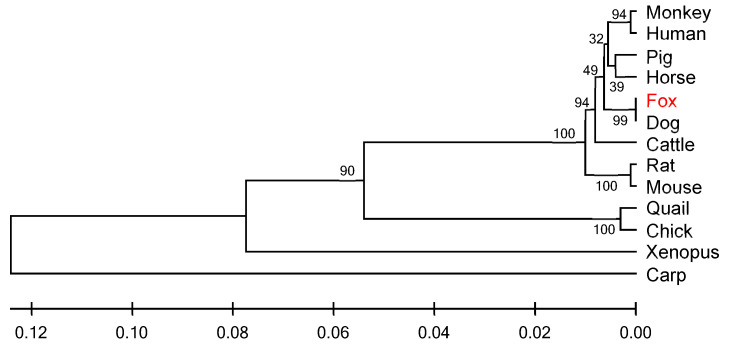 Figure 3
Figure 3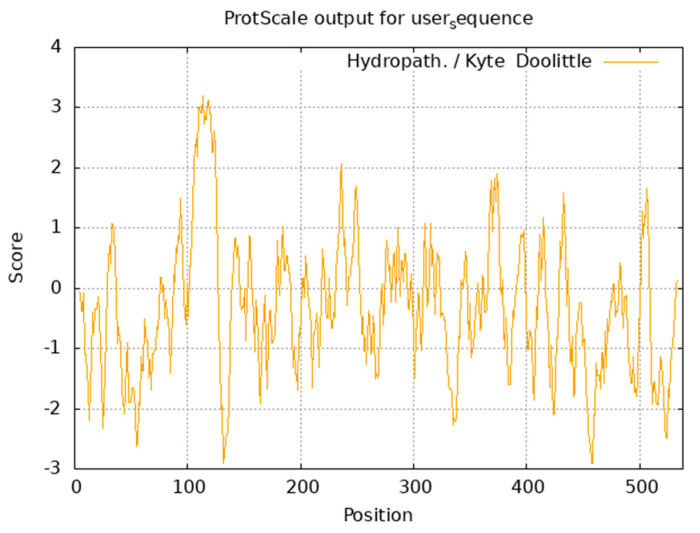 Figure 4
Figure 4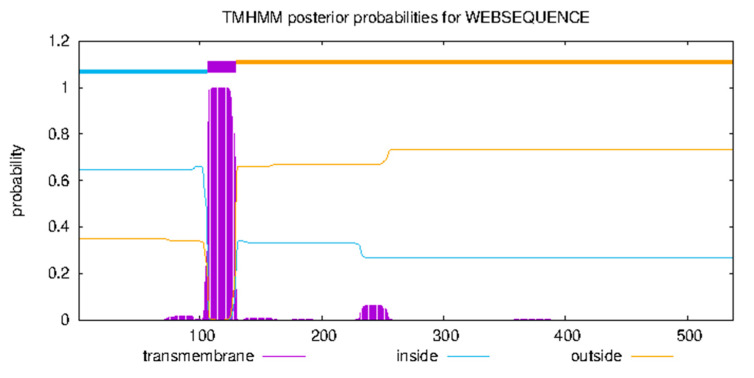 Figure 5
Figure 5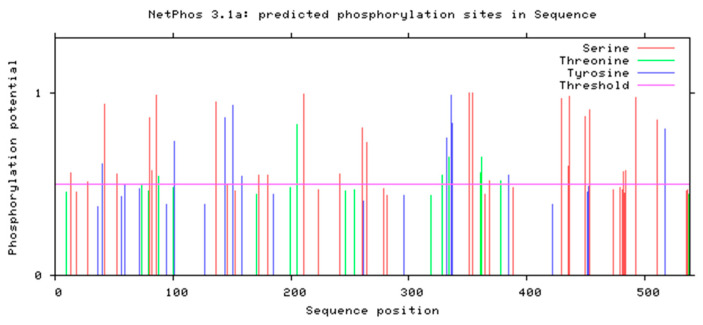 Figure 6
Figure 6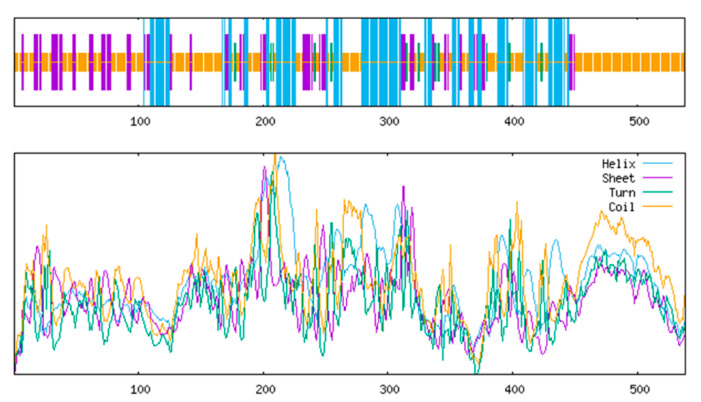 Figure 7
Figure 7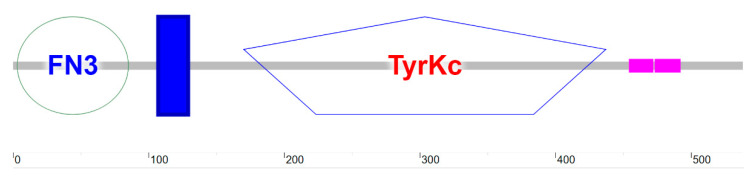 Figure 8
Figure 8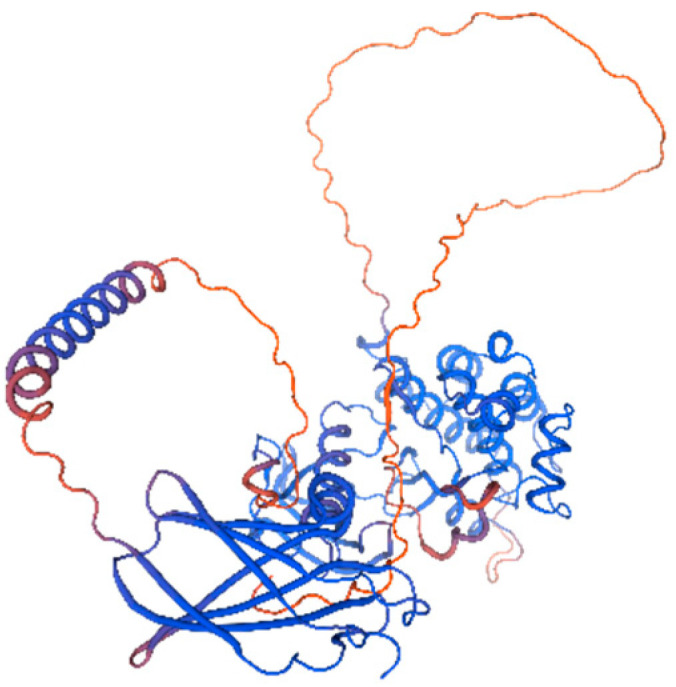 Figure 9
Figure 9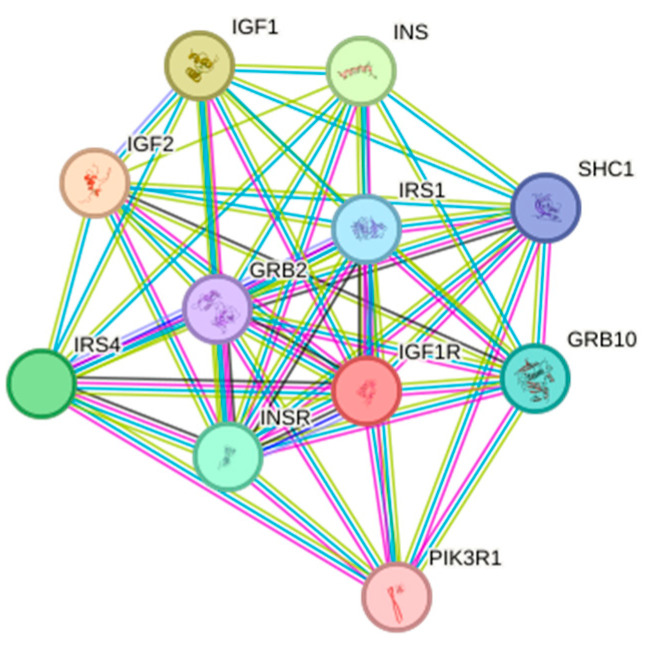 Figure 10
Figure 10- —National Key R&D Program of China
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsGrowth Hormone and Insulin-like Growth Factors · Muscle Physiology and Disorders · Adipose Tissue and Metabolism
1. Introduction
Alopex lagopus is a significant species in the global fur trade, prized for its exceptionally soft texture and pristine fur coloration, which holds irreplaceable value in luxury markets. Body size directly determines pelt surface area—the key grading criterion for commercial valuation—making the breeding of large-sized Arctic foxes a top priority. However, current research on growth regulation remains limited to phenotypic factors, like husbandry conditions (e.g., temperature/humidity) and nutritional management, while molecular genetic mechanisms remain underexplored, hindering the development of precision breeding technologies.
The insulin-like growth factor 1 receptor (IGF1R) serves as a central regulator in the vertebrate growth axis, driving cellular proliferation, differentiation, and organ development by mediation in the IGF-1/IGF-2 signaling pathway [1,2]. Functional studies demonstrate that IGF1R deficiency results in embryonic developmental retardation [3,4], significantly reduced body size [5], while its overexpression correlates with malignant cellular proliferation [6], collectively establishing its pivotal role in growth-regulatory networks. In mammals, IGF1R expression displays distinct tissue specificity and temporal dynamics: during embryogenesis, it shows predominant expression in growth-related organs (liver and skeletal muscle), synchronizes with sexual maturation processes in juvenile reproductive/endocrine organs (testes and pituitary), and transitions to maintaining metabolic homeostasis in adulthood [7,8,9].
Compared to molecular biology research on domestic animals such as pigs and cattle [10], molecular biology research on the Arctic fox remains in its infancy. Current studies are predominantly focused on the regulation of hair follicle development by Wnt signaling pathway genes and the seasonal influence of the MC1R gene on coat color [11,12], while research on key growth-axis genes is largely absent. As a canid species, the Arctic fox shares 99% homology in the IGF1R gene with canids and exhibits similar growth-regulatory pathways [13]. However, its expression patterns in Arctic fox-specific tissues (e.g., testes and muscle) and developmental stages (e.g., rapid growth phase) remain uncharacterized. Given the IGF1R gene’s direct correlation with body size and growth rate in other species, combined with the Arctic fox industry’s urgent demand for larger-sized breeding lines, this gene represents an ideal candidate for elucidating growth regulation mechanisms in this species.
IGF1R is a well-established critical gene in mammals; however, whether its expression and functions are conserved in Arctic foxes remains unverified. This study aims to investigate the structural and functional characteristics of the IGF1R gene in Arctic foxes, thereby elucidating its role in their growth and development, and to establish a foundation for more in-depth molecular biological studies on this species.
2. Materials and Methods
2.1. Experimental Materials
2.1.1. Experimental Animals
This experiment was conducted with approval from the Animal Ethics Committee of Northeast Agricultural University. Experimental animals were individually housed in separate cages, fed three times daily with ad libitum access to water. A total of 20 Arctic foxes were used, with 3 individuals sampled at each developmental stage (2-, 4-, and 6-month-old individuals). All animals were provided by the Fur-Bearing Animal Breeding Base of Tuqiang Forestry Bureau in the Greater Khingan Mountains (122°48′ E, 52°48′ N; average elevation 454 m). Euthanasia was performed by electrical shock.
2.1.2. Main Reagents
Molecular cloning system: pMD18-T vector, rTaq DNA polymerase, and restriction endonucleases (Takara Bio, Dalian, China).
RNA extraction system: TRIzol reagent (Invitrogen, Carlsbad, CA, USA), DEPC (Sigma, STL, MO, USA).
Nucleic acid purification system: DH5α competent cells (Tiangen Biotech, Beijing, China), Gel Recovery Kit, and Plasmid Extraction Kit (Tiangen Biotech).
RT-PCR system: PrimeScript RTase and SYBR Premix Ex Taq (Tiangen Biotech).
2.2. Experimental Methods
2.2.1. Tissue Sample Collection
Samples were collected at 2, 4, and 6 months of fox growth. At each stage, twelve organ tissues—liver, spleen, kidney, lung, stomach, pancreas, brain, pituitary gland, duodenum, testis, heart, and muscle—were collected for total RNA extraction. All organ tissue samples were immediately flash-frozen in liquid nitrogen and stored at −80 °C until use.
2.2.2. Total RNA Extraction and cDNA Synthesis
Total RNA was extracted using an RNA extraction kit and stored at −80 °C. RNA concentration and purity were measured with a UV spectrophotometer. High-integrity and high-purity RNA was reverse-transcribed into cDNA using a reverse transcription kit and stored at −20 °C.
2.2.3. Cloning, Sequencing, and Bioinformatics Analysis of IGF1R Gene CDS Region
Primers were designed based on homologous sequences from various species (see Table 1). The CDS of the Arctic fox IGF1R gene was cloned via PCR, using cDNA derived from liver tissue of 6-month-old individuals. The PCR reaction system (25 µL total volume) included sterile deionized water (16.3 µL), tissue cDNA (2.0 µL), 10× PCR Buffer (2.5 µL), dNTP (2.5 mmol/L, 2.0 µL), forward and reverse primers (10 pmol/µL, 1.0 µL each), and Taq polymerase (0.5 U/µL, 0.2 µL). The amplification protocol was as follows: initial denaturation at 94 °C for 5 min; 35 cycles of denaturation at 94 °C for 40 s, annealing at temperatures as listed in Table 1 (80 s, 50 s, and 40 s, respectively), and extension at 72 °C for 60 s; and a final extension at 72 °C for 10 min. PCR products were electrophoresed on 1% agarose gels, purified using a gel recovery kit, ligated into the pMD18-T molecular vector, and transformed. Then, positive clones were selected for sequencing.
Sequencing fragments were assembled using DNAMAN 8.0, and the IGF1R gene structure was annotated using the canine reference genome. Cross-species homology alignment was performed via NCBI Nucleotide BLAST. The open reading frame (ORF) was identified using ORF Finder, and amino acid sequences were translated via DNAMAN. Protein conserved domains were analyzed using Protein BLAST 2.2.26; secretory signal peptides were predicted with SignalP3.0. Multiple sequence alignment was conducted using ClustalX1.83, and a molecular phylogenetic tree was constructed via the neighbor-joining method in MEGA 4.0.
Bioinformatics analysis of Arctic fox IGF1R included physicochemical properties, hydrophobicity, transmembrane domains, functional domains, phosphorylation sites, secondary/tertiary structures, and protein interactions. Software tools and prediction functions are listed in Table 2.
2.2.4. Spatiotemporal Expression Analysis of IGF1R Gene in the Arctic Fox
qRT-PCR was used to quantify IGF1R expression in 12 organ tissues (liver, spleen, kidney, lung, stomach, pancreas, brain, pituitary gland, duodenum, testis, heart, and skeletal muscle) from 2-, 4-, and 6-month-old individuals (n = 3). The reaction system (25 µL) included sterile deionized water (16.3 µL), cDNA template (1 µg/µL, 2.0 µL), PCR Buffer (2.5 µL), dNTP (2.5 mM, 2.0 µL), forward/reverse primers (1.0 µL each), and rTaq polymerase (0.2 µL). Amplification conditions were 94 °C for 5 min; 35 cycles of 94 °C for 30 s, 60 °C for 30 s; 72 °C for 40 s; and a final extension at 72 °C for 10 min. Relative expression levels were calculated using the 2^−ΔCt^ method, with GAPDH as the reference gene. The analytical method involved calculating IGF1R gene expression levels across samples using the 2^−ΔCt^ formula [ΔCt = Ct (IGF1R) − Ct (GAPDH reference gene)]. One-way ANOVA was performed using GraphPad Prism 10.1 to compare mRNA expression levels among samples.
3. Results and Analysis
3.1. IGF1R Gene CDS Amplification Results
Total RNA extracted from the organ tissues of Arctic foxes was assessed using Nanodrop spectrophotometry, with A260/A280 ratios ranging between 1.8 and 2.0, confirming that RNA integrity met the requirements for molecular cloning. PCR amplification of cDNA templates using primers RIGF1R1-3 generated specific bands of 850 bp, 896 bp, and 727 bp, as verified by 1.5% agarose gel electrophoresis (Figure 1).
3.2. Sequencing Results and Sequence Analysis
Sequencing data assembly yielded a 2411 bp cDNA fragment of the Arctic fox IGF1R gene, consistent with the predicted structure of canine IGF1R. NCBI BLAST analysis demonstrated cross-species homology: 98% with Canis, 93% with Equus caballus, 91% with Homo sapiens, Macaca mulatta, Sus scrofa domesticus, and Ovis aries, 90% with Ailuropoda melanoleuca and Bos taurus, and 88% with Mus musculus. Phylogenetic analysis confirmed its conservation among mammalian IGF1R genes. Partial nucleotide and amino acid sequence alignments are detailed in Figure 2.
3.3. Similarity Analysis and Phylogenetic Tree Construction of IGF1R Gene
Comparative analysis of IGF1R amino acid sequences from Arctic foxes and 12 vertebrates (including Canis, Felis catus, and Bos taurus) revealed cross-class conservation: nucleotide/amino acid similarities ranged from 71% to 99%, with the highest homology to canids (99%) and the lowest to Cyprinus carpio (71%) (Table 3). A UPGMA phylogenetic tree constructed using MEGA 4.0 (Figure 3) showed Arctic foxes forming a monophyletic clade with canids, while primates exhibited topological anomalies, suggesting potential lineage-specific evolution of IGF1R in primates. Genetic distance analysis via the Kimura 2-parameter method indicated zero net distance between Arctic foxes and canids, and a maximum distance of 0.271 with Cyprinus carpio (Table 4), aligning with vertebrate phylogenetic hierarchy.
3.4. Structural and Functional Analysis of Arctic Fox IGF1R Protein
3.4.1. Physicochemical Properties and Hydrophobicity
ExPASy ProtParam analysis revealed the Arctic fox IGF1R protein (molecular formula, C2702H4203N725O805S28) has a molecular weight of 60.62 kDa, comprising 538 amino acids. Dominant residues included glutamate (Glu, 9.1%) and leucine (Leu, 9.1%), while histidine (His, 1.5%) and tryptophan (Trp, 1.5%) were least abundant. Acidic residues (Asp + Glu = 73) exceeded basic residues (Arg + Lys = 56) (Table 5), with a theoretical pI of 5.15, confirming acidic properties. The protein exhibited hydrophilicity (average GRAVY = −0.337), thermolability (instability index = 48.59), and moderate lipophilicity (aliphatic index = 81.51). Kyte–Doolittle hydrophobicity analysis identified a maximum hydrophobic peak (+3.189) at Ala114 and a minimum (−2.911) at Asp457 (Figure 4).
3.4.2. Transmembrane Domain and Subcellular Localization
Prediction indicated a single α-helix transmembrane domain (107–129 aa), with the N-terminus (1–106 aa) being cytoplasmic and the C-terminus (130–538 aa) being extracellular (Figure 5), consistent with type-II transmembrane receptors. PSORT II predicted nuclear (34.8%), cytosolic (21.7%), mitochondrial (21.7%), and secretory vesicle (8.7%) localization, suggesting dynamic involvement in transmembrane signaling and vesicle trafficking, aligning with IGF1R-mediated PI3K/AKT and MAPK pathway activation.
3.4.3. Phosphorylation Site Prediction
NetPhos 3.1 (threshold > 0.5) predicted 27 potential phosphorylation sites: 18 serine (66.7%), 6 threonine (22.2%), and 3 tyrosine (11.1%) residues (Figure 6).
3.4.4. Secondary and Tertiary Structure Prediction
SOPMA analysis indicated dominance of random coils (52.79%) and α-helices (26.39%), with fewer extended strands (16.91%) and β-turns (3.90%) (Figure 7). SMART identified three domains: an N-terminal fibronectin type-III domain (3–85 aa) for ligand binding, a transmembrane helix (107–129 aa), and a C-terminal tyrosine kinase catalytic domain (170–437 aa) (Figure 8). SWISS-MODEL tertiary structure prediction (Figure 9) corroborated these features.
3.4.5. Protein Interaction Analysis
STRING database analysis revealed an 11-node interaction network with high connectivity (average node degree = 9.09) and clustering (local clustering coefficient = 0.911). Core interactors included insulin signaling components (INS/IGF1/IGF2), adaptors (SHC1/GRB2/GRB10), and PI3K-AKT regulators (PIK3R1/IRS1/IRS4), enriched in cell proliferation and organ development processes (Figure 10).
3.5. Tissue-Specific and Spatiotemporal Expression Patterns of IGF1R Gene in the Arctic Fox
qRT-PCR showed IGF1R expression in 11 of 12 organs (excluding duodenum) in 6-month-old Arctic foxes, with the highest levels in the testis and the lowest in the stomach (Table 6). Temporal analysis indicated age-dependent increases in liver expression, peak testis expression at 6 months, and highest muscle expression at 4 months (Table 7).
4. Discussion
Insulin-like growth factor I receptor (IGF1R), as a core effector molecule in the growth hormone axis cascade, coordinates organismal development by maintaining cellular homeostasis and driving proliferation–differentiation programs. As a key mediator of the GH-IGFs signaling axis, it exerts pleiotropic effects in metabolic regulation [1,14,15]. Loss-of-function studies demonstrate that IGF1R knockout leads to embryonic muscle-developmental defects in mice [16], hepatic regeneration disorders, and pulmonary developmental abnormalities [17], while muscle-specific overexpression induces abnormal muscle hypertrophy [5]. Evolutionary analyses reveal that structural variations in and transcriptional–translational efficiency attenuation of the IGF1R gene constitute critical mechanisms underlying dwarfism phenotypes, validated in both murine models and human genetic cases. Compared to extensive research on pigs [8], cattle [18], sheep [19], poultry [20], and aquatic species [21], functional characterization of IGF1R in Arctic foxes remains largely unexplored. This study systematically elucidates its molecular features and expression regulatory patterns for the first time, providing novel perspectives on growth axis research in polar mammals.
Based on the domestic dog IGF1R gene (Accession No. XM_545828), primers were designed to successfully clone a partial cDNA sequence (2411 bp) of Arctic fox IGF1R using RT-PCR. This sequence showed high homology with mammalian IGF1R, with the complete CDS region (1617 bp) encoding 538 amino acids. Phylogenetic analysis revealed 99% nucleotide sequence homology between Arctic fox IGF1R and domestic dogs, significantly higher than homology with pigs (98%), mice (96%), and poultry (84%), consistent with mammalian taxonomic relationships. Notably, IGF1R evolutionary trees exhibited deviations from traditional classifications in primates, suggesting potential functional divergence of the IGF family in primates. Phosphorylation prediction identified 27 potential phosphorylation sites, with structural domain remodeling induced by site-specific phosphorylation potentially regulating multi-pathway signal transduction networks [22]. Six Ser sites localized in the tyrosine kinase catalytic domain may mediate intracellular signaling through autophosphorylation [23]. Functional domain analysis demonstrated that Arctic fox IGF1R contains an 83-amino-acid fibronectin type-III (FN3) domain mediating DNA/heparin binding [24], while its transmembrane region (107–129 aa) confirms its identity as a transmembrane receptor. The tyrosine kinase (TyrKc) domain harbors multiple binding sites, enabling diverse catalytic reactions [25]. Protein interaction network analysis revealed strong interactions between IGF1R and 11 proteins, including IGF1/IGF2/INS. IGF1, as a key member of the insulin-like growth factor family, plays vital roles in growth regulation, skeletal muscle development, cellular proliferation/differentiation, tissue repair, and metabolic modulation. Importantly, IGF1R not only mediates IGF ligand signaling but also critically regulates embryonic development, cell growth/differentiation, mitosis, embryonic survival, and metabolic adaptation [26]. IGF1 drives evolutionary processes by modulating growth rates, body size differentiation, and physiological adaptability. IGF1R-IGF2 binding triggers tyrosine kinase domain autophosphorylation, activating PI3K/AKT and MAPK/ERK signaling cascades to promote metabolism, proliferation, and differentiation [26]. Genomic analyses identify numerous SNPs in porcine and ovine IGF1R genes, with specific variants significantly influencing growth traits through receptor conformation and expression levels, contributing to breed-specific size variations [27,28]. Notably, bovine IGF1R polymorphisms correlate strongly with milk-production traits (milk fat/protein content), highlighting its potential as a molecular marker in dairy cattle breeding [29]. These findings collectively underscore the central role of IGF signaling in animal growth regulation.
Using qRT-PCR (with GAPDH as reference), tissue-specific expression profiling of IGF1R in 6-month-old Arctic foxes revealed differential expression across the 12 organ tissues, except duodenum. Testicular IGF1R mRNA levels significantly exceeded other organ tissues, followed by kidney, muscle, liver, pituitary, brain, spleen, lung, heart, pancreas, and stomach. Notably, the absence of IGF1R expression in duodenal tissue may indicate its limited physiological role in this organ [30]; suggest predominant reliance on alternative signaling pathways for growth regulation [31]; or reflect technical limitations, such as undetectable expression levels (below qPCR detection threshold) or RNA degradation during preservation, affecting assay sensitivity.
Developmental expression analysis in liver, testis, and muscle tissues at 2-, 4-, and 6-month-old stages demonstrated age-dependent expression patterns. Hepatic expression showed progressive upregulation (2-month-old levels significantly lower than subsequent stages), aligning with developmental patterns observed in Nanjiang yellow goats [27]. Testicular expression peaked at 6 months (6-month-old > 2-month-old > 4-month-old), while muscular expression reached maximum at 4 months (4-month-old > 6-month-old > 2-month-old). The inclusion of testicular tissue in developmental studies stemmed from its highest expression among 12 organ tissues at 6 months, suggesting IGF1R involvement in Arctic fox reproductive regulation—a hypothesis reinforced by sustained high expression across 2- and 4-month-old testicular samples.
The spatiotemporal expression of IGF1R across tissues carries significant biological implications: In liver tissue, IGF1R expression increased with age, reflecting its core role in energy metabolism and growth substrate synthesis. As the primary site of IGF-1 synthesis, upregulated receptor expression in liver may enhance responsiveness to circulating IGF-1, promoting glycogen synthesis and protein accumulation to support rapid growth in juvenile foxes [32]. Testicular tissue expression peaked at 6 months old, suggesting IGF1R’s potential involvement in Arctic fox sexual maturation by regulating spermatogonial proliferation and testosterone synthesis, thereby influencing reproductive organ development and breeding performance [33]. The muscular expression peak at 4 months old coincided with the rapid growth phase of skeletal muscle, demonstrating that IGF1R directly regulates muscle growth efficiency through satellite cell differentiation and myofiber hypertrophy—consistent with its established role in mammalian myogenic differentiation [17].
As a transmembrane tyrosine kinase receptor, IGF1R ubiquitously localizes on cell membranes, mediating proliferation, differentiation, and anti-apoptotic functions through IGF1/IGF2 binding [34]. Current research predominantly focuses on its roles in tumorigenesis and therapeutic resistance, whereas tissue-specific functions in Arctic foxes require further exploration via proteomic interactome mapping and conditional knockout models. This study provides foundational insights into Arctic fox IGF1R expression profiles and structural characteristics, facilitating future investigations into molecular mechanisms underlying growth trait regulation.
5. Conclusions
This study successfully cloned the full CDS (1617 bp) of IGF1R in the Arctic fox, with phylogenetic analysis confirming its high conservation (99% homology) within canines. The receptor exhibited characteristic tyrosine kinase features, including a fibronectin type-III ligand-binding domain, transmembrane region, and catalytic domain, with acidic hydrophilic properties (pI = 5.15). Spatiotemporal expression analysis revealed tissue-specific and developmental stage-specific regulation: IGF1R expression was highest in testicular tissue, while dynamic patterns were observed across ages—liver expression increased progressively with postnatal development, aligning with enhanced metabolic demands; testicular expression peaked in 6-month-old individuals, suggesting roles in sexual maturation; and muscular expression maxima at 4 months of age corresponded to rapid skeletal muscle growth.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Alex P. Kanakkaparambil R. Gopalakrishnan R. Ramasamy C. Thazhathuveettil A. The effect of insulin-like growth factor 1 receptor gene single nucleotide polymorphism on growth and milk production traits in two native Indian tropical goat breeds Anim. Biotechnol.2023344828483610.1080/10495398.2023.219746837029727 · doi ↗ · pubmed ↗
- 2Baker J. Liu J.P. Robertson E.J. Efstratiadis A. Role of insulin-like growth factors in embryonic and postnatal growth Cell 199375738210.1016/S 0092-8674(05)80085-68402902 · doi ↗ · pubmed ↗
- 3Constância M. Hemberger M. Hughes J. Dean W. Ferguson-Smith A. Fundele R. Stewart F. Kelsey G. Fowden A. Sibley C. Placental-specific IGF-II is a major modulator of placental and fetal growth Nature 200241794594810.1038/nature 0081912087403 · doi ↗ · pubmed ↗
- 4Woods K.A. Camacho-Hübner C. Savage M.O. Clark A.J. Intrauterine growth retardation and postnatal growth failure associated with deletion of the insulin-like growth factor I gene N. Engl. J. Med.19963351363136710.1056/NEJM 1996103133518058857020 · doi ↗ · pubmed ↗
- 5Liu J.P. Baker J. Perkins A.S. Robertson E.J. Efstratiadis A. Mice carrying null mutations of the genes encoding insulin-like growth factor I (Igf-1) and type 1 IGF receptor (IGF 1R)Cell 199375597210.1016/S 0092-8674(05)80084-48402901 · doi ↗ · pubmed ↗
- 6Galifi C.A. Wood T.L. Insulin-like growth factor-1 receptor crosstalk with integrins, cadherins, and the tumor microenvironment: Sticking points in understanding IGF 1R function in cancer Endocr.-Relat. Cancer 202330 e 23003110.1530/ERC-23-003137490874 · doi ↗ · pubmed ↗
- 7James H.A. O’Neill B.T. Nair K.S. Insulin Regulation of Proteostasis and Clinical Implications Cell Metab.20172631032310.1016/j.cmet.2017.06.01028712655 PMC 8020859 · doi ↗ · pubmed ↗
- 8Pierzchała M. Pareek C.S. Urbański P. Goluch D. Kamyczek M. Różycki M. Smoczynski R. Horbańczuk J.O. KuryłJ. Study of the differential transcription in liver of growth hormone receptor (GHR), insulin-like growth factors (IGF 1, IGF 2) and insulin-like growth factor receptor (IGF 1R) genes at different postnatal developmental ages in pig breeds Mol. Biol. Rep.2012393055306610.1007/s 11033-011-1068-821695430 · doi ↗ · pubmed ↗