Bax Inhibitor MoBi-1 Is Required for Conidiation, Pathogenicity, and Stress Responses in Magnaporthe oryzae
Shuai Meng, Yangyang Shen, Dixuan Zhang, Liutao Bao, Hao Cao, Gening Song, Chenshun Xie, Jane S. Jagernath, Guoqiang Shen, Jie Chen

TL;DR
This study explores the role of the Bax inhibitor MoBi-1 in the rice pathogen Magnaporthe oryzae, revealing its importance in fungal development, stress response, and disease.
Contribution
The study reveals that MoBi-1 lacks Bax inhibitory function but is crucial for conidiation, pathogenicity, and mitochondrial energy metabolism in M. oryzae.
Findings
MoBi-1 is essential for conidiation and pathogenicity in M. oryzae.
MoBi-1 plays a role in stress adaptation and mitochondrial energy metabolism.
MoBi-1 does not inhibit Bax-induced programmed cell death in this species.
Abstract
Magnaporthe oryzae serves as a model organism for studying the molecular biology of filamentous fungi and the pathogenic mechanisms of fungal pathogens. It also poses a significant threat to rice production in China. Bax inhibitor-1 (Bi-1), a protein with evolutionary conservation, functions as an inhibitor of programmed cell death induced by the proapoptotic protein Bax. Despite the widespread presence of Bi-1 proteins in hyphal fungi, their biological functions have not been extensively characterized. Here, we characterized the function of MoBI-1, a putative Bax-inhibitor protein in M. oryzae, which is located in the mitochondria and participates in conidiation, stress adaptation, and pathogenicity. Further investigations revealed that MoBi-1 is also essential for the regulation of mitochondrial energy metabolism. Remarkably, experimental evidence indicates that MoBi-1 does not seem…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
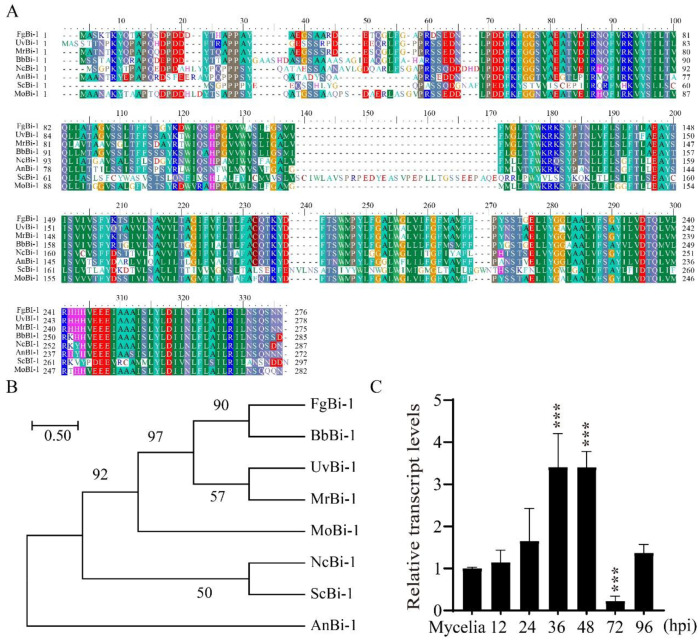 Figure 1
Figure 1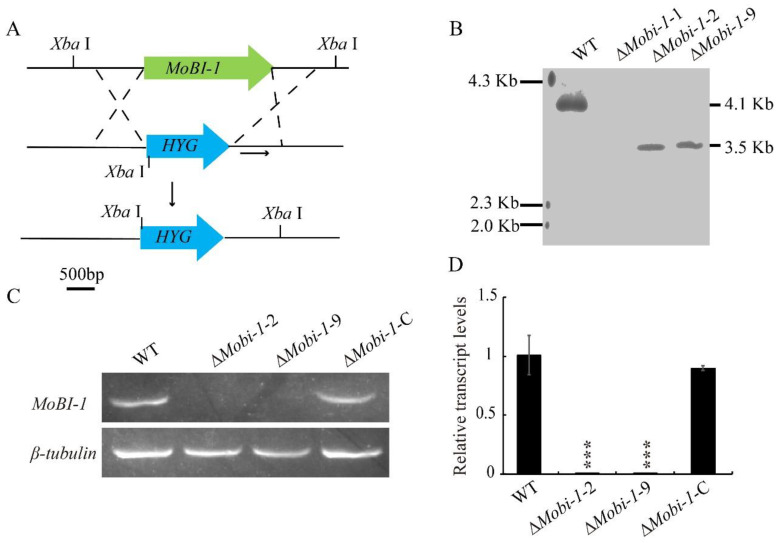 Figure 2
Figure 2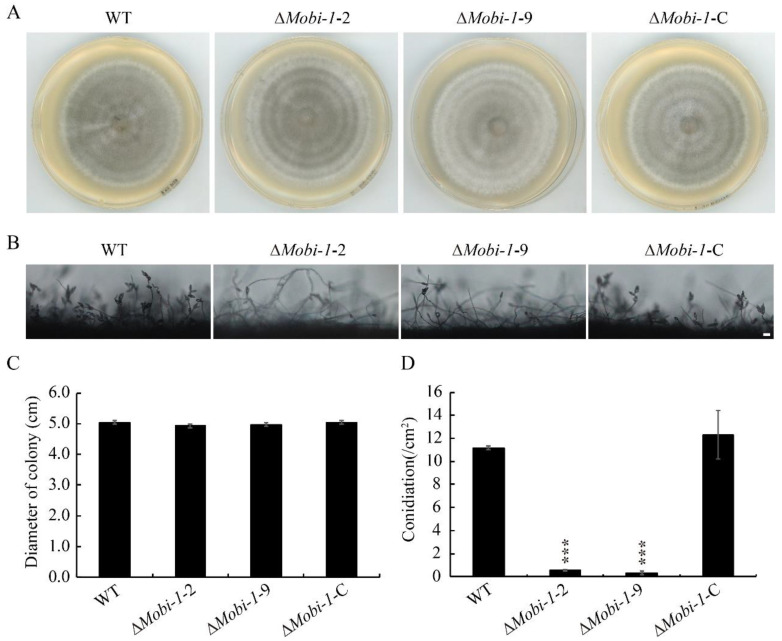 Figure 3
Figure 3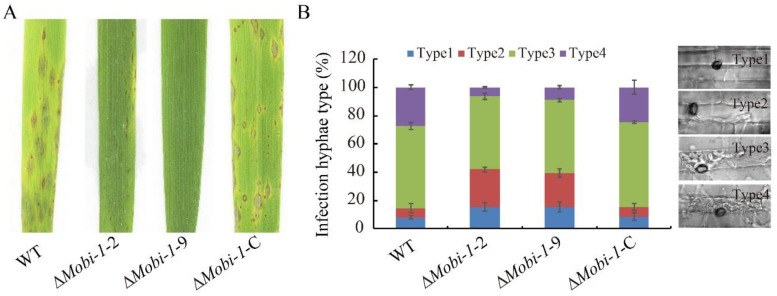 Figure 4
Figure 4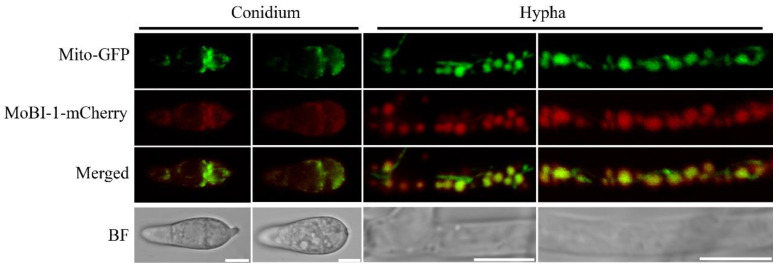 Figure 5
Figure 5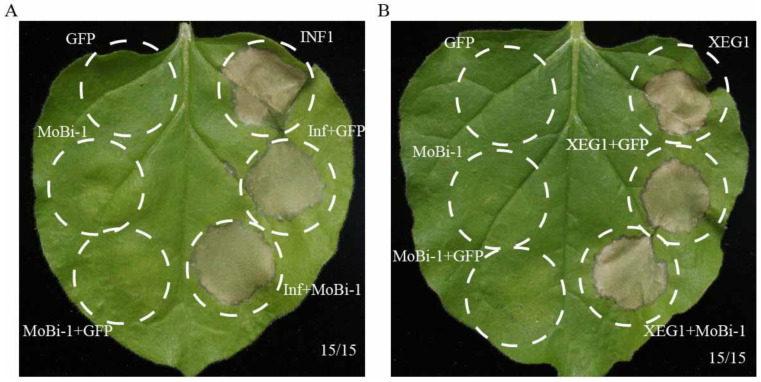 Figure 6
Figure 6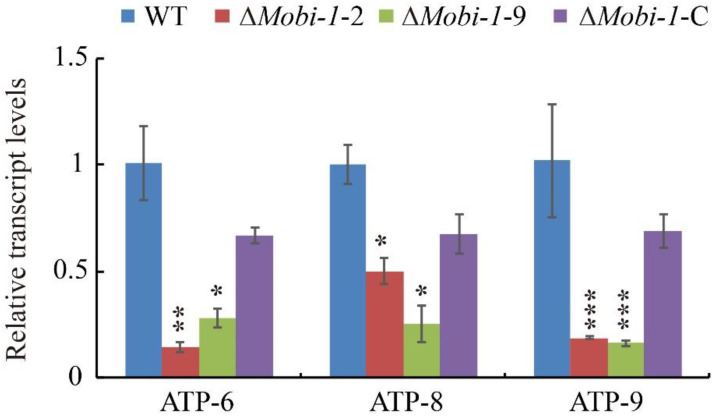 Figure 7
Figure 7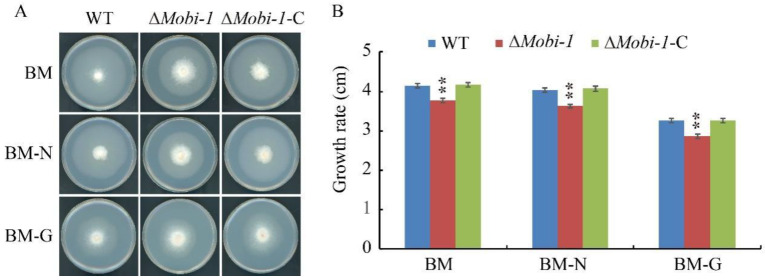 Figure 8
Figure 8- —National Key Research and Development Program of China
- —Zhejiang Provincial Natural Science Foundation of China
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsFungal and yeast genetics research · Fungal Biology and Applications · Plant-Microbe Interactions and Immunity
1. Introduction
Rice blast, caused by the filamentous ascomycete Magnaporthe oryzae (the rice blast fungus), is a highly destructive disease of rice that poses a significant threat to global rice production [1,2]. Mitochondria, as essential organelles in eukaryotic cells, not only provide energy but also participate in vital cellular activities such as cell signaling, differentiation, senescence, and death [3,4]. Recent research has revealed that mitochondria play a crucial role in fungal development and virulence, and mitophagy is a critical process in the pathogenicity of M. oryzae [5,6,7,8,9,10].
Bax inhibitor-1 (Bi-1) is a highly conserved multifunctional protein during evolution, primarily found in plants, fungi, bacteria, and viruses. The BI-1 gene can attenuate or suppress stress-induced physiological cell death and plays a critical regulatory role in the interaction between programmed cell death and external stress [11]. Bi-1 was initially identified in yeast as a homolog of a human protein that inhibits Bax-mediated cell death processes [12]. The Bi-1 protein contains multiple transmembrane domains, is predominantly localized to the endoplasmic reticulum (ER), and acts as a resident sensor of the ER. In the mitochondrial pathway, Bi-1 inhibits the translocation of Bax from the cytosol to the mitochondria. Since Bi-1 interacts with the BH4 domain, it cannot directly bind to Bax or Bak. Instead, it modulates the ratio between Bi-1/Bcl-2/Bcl-XL and Bax by interacting with Bcl-2 and Bcl-XL, thereby influencing apoptosis [13,14]. Bi-1 possesses ion channel activity. In the endoplasmic reticulum (ER) pathway, cytosolic Ca^2^⁺ can leak extracellularly through this channel, triggering stress responses that induce apoptosis. Thus, Bi-1 suppresses mammalian cell apoptosis by regulating Ca^2^⁺ homeostasis [15]. When stress activates apoptosis-related signals on the ER, it also activates Bax, thereby linking these two pathways. The Bax inhibitor-1 (Bi-1) family consists of compact transmembrane proteins with significant anti-apoptotic activity. These evolutionarily conserved proteins are widely distributed across species and play key roles in regulating programmed cell death, cellular homeostasis, and stress responses [16,17].
In Arabidopsis, overexpression of AtBI-1 suppresses Bax, H_2_O_2_, and salicylic acid-induced plant cell death and delays methyl jasmonate-induced leaf senescence by inhibiting MAP6 activation [18]. In Nicotiana benthamiana, silencing of NbBI-1 reduces autophagy activity induced by N gene-mediated resistance to tobacco mosaic virus (TMV) and methyl viologen (MV), whereas overexpression of NbBI-1 enhances autophagic activity, triggering autophagy-dependent cell death [19]. The wheat TaBI-1 gene plays a critical role in plant resistance to salt and heat stress [20]. Ectopic expression of cotton GhBI-1A and GhBI-1B genes in Arabidopsis significantly improves tolerance to salt and tunicamycin [21]. In Metarhizium robertsii, the MrBI-1 knockout mutant exhibits increased sensitivity to heat shock and is essential for virulence [11]. Silencing of Ss-BI1 in Sclerotinia sclerotiorum reduces its pathogenicity, thermotolerance, and ER stress tolerance. In Ustilaginoidea virens, UvBI-1 regulates vegetative growth, conidia production, stress tolerance, rice coleoptile elongation, and pathogenicity [22]. However, the biological functions and mechanisms of Bi-1 in M. oryzae remain largely unclear.
In this study, we demonstrated that MoBi-1 encodes a Bax inhibitor-1 (Bi-1) protein that is localized in the mitochondria and plays roles in conidiation, stress adaptation, and pathogenicity in M. oryzae. We also found that MoBi-1 plays an important role in mitochondrial energy metabolism. Additionally, our study indicates that MoBi-1 may not take part in suppressing the Bax-triggered cell death, and therefore have no function as a Bax inhibitor.
2. Materials and Methods
2.1. Sequence Analysis
All gene and protein sequences used in this study were downloaded from the National Center for Biotechnology Information (NCBI, https://www.ncbi.nlm.nih.gov/ 11 June 2021). Protein sequence alignments were performed using BioEdit, and the phylogenetic analyses were conducted using the MEGA 7.0 software [23].
2.2. Fungal Strains and Culture Media
The M. oryzae strain B157 was used as the wild type in this study. All the strains were cultured on complete medium (CM), basal medium (BM), basal medium without NH_4_NO_3_ (BM-N), and basal medium without glucose (BM-G) agar plates under 16 h of light and 8 h of dark at 25 °C [24]. Colony diameters were measured on CM plates at 7 days old, and the conidia were collected from 7-day-old colonies cultured on CM plates for testing. Three separate biological experiments were performed with three replicates at a time. The data were subjected to Duncan’s multiple range test.
2.3. Construction of the ΔMoBI-1 Strains and Complementation Analyses
To construct the MoBI-1 deletion vector, the 985 bp upstream and 1125 bp downstream flanking fragments of MoBI-1 were amplified from the genomic DNA of the B157 strain using the primer pairs MoBI-5F/R and MoBI-3F/R (Table S1). Then, these flanking fragments were ligated into Hind III/Hind III and XhoI/BamHI sites of pFGL821 (Addgene: 58224, www.addgene.org 29 June 2021) [25]. For the complementation, a genomic sequence driven by the MoBI-1 native promoter and 3′ UTR region was amplified and inserted into EcoRI/KpnI-digested pFGL822 (Addgene: 58225, www.addgene.org 2 August 2021) with a glufosinate ammonium resistance gene. The complemented construct was transformed into the ΔMobi-1 strain. Agrobacterium-mediated transformation (ATMT) was applied for genetic transformation in M. oryzae. The correct transformants of ΔMoBI-1 and the complementation assay were ascertained using Southern blot and qRT PCR analyses.
2.4. Southern Blotting, RNA Isolation, and qRT-PCR Analyses
The genomic DNA from the WT and mutant strains was extracted and digested with XbaI. Then, the digested products of genomic DNA were separated on a 0.8% agarose gel and mounted onto a positively charged nylon membrane (GE Healthcare, London, UK). The MoBI-1 gene probe was amplified from B157 genomic DNA using the primers MoBI-probeF/R (Table S1). The purified probe MoBI was subsequently DIG-labeled with Labeling Reagents (GE Healthcare, London, UK) to hybridize with the digested DNA products from the WT and ΔMoBI-1 mutant strains. The hybridization procedure was carried out according to the manufacturer’s instructions for the Amersham^TM^ AlkPhos Direct Labeling Reagents (GE Healthcare, London, UK). The signals of Southern blotting were detected by a ChemiDoc XRS+ system (Bio-Rad, Hercules, CA, USA).
Total RNA was extracted using the Fungal RNA Kit 200 (OMEGA, Norcross, GA, USA) according to the manufacturer’s instructions from the mycelia of WT and ΔMobi-1 cultured in liquid CM. The cDNA of the first strand was synthesized by a reverse transcription kit (TaKaRa, Ōsaka shi, Japan) and analyzed by qRT-PCR using the TB Green^TM^ Premix Ex Taq^TM^ (Tli RnaseH Plus) (TaKaRa, Japan, Ōsaka shi).The β-tubulin gene was used as the endogenous reference gene, and the expression levels of MoBI-1, ATP-6, ATP-8, and ATP-9 were calculated using the 2^−∆∆Ct^ method [26]. The primers used are listed in Table S1.
2.5. Pathogenicity and Rice Infection Assay
For plant infection assays, the rice cultivar Oryza sativa cv. CO39, grown for three weeks, was used for the virulence test. Conidia harvested from B157, ΔMobi-1, and ΔMobi-1-C were suspended in a 0.1% (w/v) gelatin solution (1 × 10^5^ conidia/mL), then two milliliters of each solution was sprayed onto the plants. The disease lesions were photographed after 5 days post-inoculation under full humidity conditions at 25 °C. The infection process assay was conducted on a rice sheath cell. A rice sheath 5 cm in length was cut and dropped with about 50 μL of conidial suspension (1 × 10^5^ conidia/mL). After incubation for 12–48 h in a sealed chamber, a single layer of sheath cell was shaved to observe the invasive hyphae growing in the rice cell.
2.6. Subcellular Localization
For the fluorescent microscopic observation, the MoBI-1 gene with a native promoter and an open reading frame (ORF) was cloned with primers and ligated into the pFGL822-GFP [27] digested by EcoRI/KpnI sites. The subsequent vector MoBi-1-mCherry was transformed into the Mito-GFP-expressing strain to produce coexpression transformants. Rescued transformants with GFP and mCherry signals were observed and documented by confocal fluorescent LSM700 microscope (Carl Zeiss Inc., Oberkochen, Germany). GFP and mCherry were imaged with 488 and 543 nm laser excitation, respectively. Images were conducted with ImageJ v 1.8.0 and Photoshop CS6 software.
2.7. Agrobacterium-Mediated Transient Expression of Proteins in N. benthamiana
The MoBI-1 coding region was cloned and ligated into a Cla I/Sma I-digested pGR107 vector [28]. The positive transformants were confirmed by sequencing and introduced into A. tumefaciens strain GV3101 by electroporation. The overnight-cultured Agrobacterium was collected and resuspended in 10 mM MgCl_2_ buffer. The final cell density was adjusted to an OD_600_ of 0.4. The A. tumefaciens containing the corresponding plasmid was infiltrated into 4–5-week-old N. benthamiana leaves using syringes without a needle. Photos were taken after 2–3 days post-inoculation.
3. Results
3.1. Identification and Knockout of MoBI-1 Gene in M. oryzae
To identify the homolog of Bi-1 in M. oryzae, the protein sequence of S. cerevisiae was used for a BLASTP search of M. oryzae in the NCBI database. The search results for S. cerevisiae Bi-1 identified the most closely matching protein, which is named MoBi-1 (XP_003720584.1) (Figure 1A). Sequence analysis using Motif Scan (http://smart.embl-heidelberg.de/, accessed on 20 October 2021) revealed that Bi-1 contained a conserved Bax inhibitor-1-like domain in fungi. A phylogenetic analysis of Bi-1 amino acid sequences from Fusarium graminearum, Beauveria bassiana, U. virens, Metarhizium robertsii, Magnaporthe oryzae, Neurospora crassa, S. cerevisiae, and Aspergillus niger showed that MoBi-1 is not closely related to Bi-1 homologs from other fungi (Figure 1B). Next, to investigate the potential role of MoBi-1, its transcript abundance was assessed via qRT-PCR across various infection stages of M. oryzae. Furthermore, an increase in MoBi-1 expression was observed during the infection process initiated by M. oryzae (Figure 1C). The results show that the transcriptional up-regulation of MoBI-1 implicated a possible role of MoBi-1 during pathogenesis in M. oryzae.
To investigate the biological roles of MoBi-1 in M. oryzae, knockout vectors carrying the hygromycin resistant cassette gene HYG were constructed and transferred into the wild-type strain (Figure 2A). Quantitative real-time PCR (qRT-PCR) and Southern blot assays were conducted to confirm the targeted gene knockout and exclude ectopic integrations (Figure 2B). The correct transformants were obtained with similar phenotypes, and ΔMobi-1-2 and -9 were chosen for further experiments (Figure 2B). Complemented strains were generated by the full-length gene copy of MoBI-1 with its native promoter into the knockout mutant ΔMobi-1-2 strain. The resultant MoBI-1-C strain was confirmed by PCR and qRT-PCR analyses, which showed that the abundance of MoBI-1 transcript was comparable to that of the WT strain (Figure 2C,D).
3.2. The Role of MoBI-1 in Morphological Development
To investigate the roles of MoBI-1 in fungal growth and development, the ΔMobi-1 mutants were cultured on complete medium (CM) agar plates after 6 days of incubation. In comparison with the WT and complementation strains, the colony diameter of ΔMobi-1 mutant showed no significant differences (Figure 3A,C), indicating that MoBi-1 may not be involved in vegetative growth. However, the ΔMobi-1 mutant produced almost no spores relative to the WT and ΔMobi-1-C complementary strains (Figure 3B,D), suggesting that MoBi-1 plays an important role in the conidia of M. oryzae.
3.3. MoBI-1 Contributes to Full Virelence
To determine whether MoBI-1 is required for pathogenicity virulence, conidial suspension (1 × 10^5^ conidia/mL) of WT, ΔMobi-1-2, ΔMobi-1-9, and ΔMobi-1-C complementary strains were sprayed onto three-week-old rice leaves. The null mutant ΔMobi-1-2 and ΔMobi-1-9 showed few lesions on the rice leaves, while numerous typical spreading lesions were caused on the WT and complementation strain-infected rice leaves (Figure 4A). To elucidate the difference in the process of infection, we conducted an infection assay on the rice sheath to be certain of the early infection hyphae growth. Conidial suspension was dropped onto the rice sheath for 12–48 h in a sealed chamber [29]. At 48 hpi, invasive hyphae were observed and analyzed, with almost 60% of cells showing a type 4 lesion, nearly 30% displaying a type 3 lesion, and less than 10% showing type1 or type 2 lesions. However, only 10% of cells showed type 4 lesions, 60% of cells exhibited type 3 lesions, and 30% showed type 1 and type 2 lesions in the ΔMobi-1 mutants (Figure 4B). These results indicated that MoBi-1 plays an important role in conidiation and pathogenicity in M. oryzae.
3.4. MoBi-1 Is Mainly Localized in the Mitochondria
Numerous studies showed that Bax inhibitor-1 (Bi-1) is a highly conserved multifunctional protein in evolution, mainly in plants, fungi, bacteria, and viruses [11,15,30,31,32]. Thus, we found that MoBi-1 was mainly located in mitochondria and distinguished from other species in localization. A red fluorescent protein sequence was fused to the C-terminus of MoBi-1 using a native promoter, and the vector was introduced into the mitochondria marker Mito-GFP strain. The results indicated that MoBi-1-mCherry could co-localize with Mito-GFP in spores and mycelia (Figure 5).
3.5. MoBi-1 Cannot Inhibit BAX-Induced Cell Death in N. benthamiana
Previous studies have reported that the Bi-1 domain has the function of inhibiting cell death induced by Bax [30,31,32,33,34,35]. Considering that MoBi-1 contains a Bi-1 domain, we ascertained whether MoBI-1 can also inhibit Bax-induced cell death. Agrobacterium strains carrying MoBI-1 and Bax were co-infiltrated into N. benthaminan leaves. MoBI-1, INF/XEG, and GFP were also expressed independently. The results showed MoBi-1 cannot inhibit the INF- or XEG-triggered cell death symptom in the infiltrated leaves (Figure 6). Furthermore, no cell death was detected when GFP and MoBi-1 separately were infiltrated into N. benthamiana leaves (Figure 6). These data indicated that MoBi-1 may not take part in suppressing the Bax triggered cell death, and therefore have no function as a Bax inhibitor.
3.6. Relative Expression of ATP Synthesis Genes Decreased
In all eukaryotic cells, the mitochondrion is a highly dynamic specialized organelle that serves as a master regulator of metabolism [36], and it is the main place of energy synthesis in the process of development [24]. Since the MoBi-1 protein localizes to the mitochondria, transcriptomes of the MoBI-1 mutants, as well as the WT and complemented strains, were analyzed for differential ATP synthesis genes, including mtATP6 (Genbank: MGG_21007), mtATP8 (Genbank: MGG_21008), and mtATP9 (Genbank: MGG_00892) [37,38]. The data showed that the expression levels of mtATP6, mtATP8, and mtATP9 were downregulated in ΔMobi-1 mutant strains in comparison with the WT and ΔMobi-1-C complementation strains (Figure 7). These results indicated that MoBi-1 plays an important role in the synthesis of mitochondrial energy metabolism.
3.7. MoBi-1 Is Involved in Responses to Nitrogen and Glucose
Carbon and nitrogen sources play an important role in fungal growth and development. To study the function of MoBi-1 in nitrogen and glucose on mycelia growth, the ΔMoBI-1 mutants, WT, and complementation strains were grown on BM medium, BM-N medium (BM medium without nitrogen source), and BM-G (BM medium without carbon source) for 7 d at 28 °C. The results showed that ΔMoi-1 mutants were not able to efficiently utilize the carbon and nitrogen sources and grew slower than the WT and ΔMobi-1-C complementation strains (Figure 8), indicating that MoBi-1 is involved in glucose and nitrogen utilization.
4. Discussion
The rice blast fungus M. oryzae is a well-known model for fungal–plant interactions. Until now, a large number of mitochondria-related genes have been identified for functional analysis. They are primarily involved in the process of fungus development, physiology, and pathogenicity [6,8,38,39]. The rice blast fungus Magnaporthe oryzae is a highly specialized pathogen renowned for its exceptional morphogenetic and biochemical adaptations for a parasitic lifestyle, posing a significant threat to rice cultivation worldwide. Substantial progress has been made in understanding the molecular mechanisms underlying this destructive disease, and numerous genes crucial for the early stages of infection have been identified [40].
The mitochondrion, a highly specialized and dynamic organelle, resides within all eukaryotic cells and functions as a central regulator of cellular metabolism [36]. These organelles ensure an uninterrupted supply of adenosine triphosphate (ATP), the primary energy source for cellular activities [41]. Phytopathogenic fungi, organisms that cause diseases in plants, acquire the energy necessary for their metabolic processes from substrates obtained from living or recently deceased plant matter [42]. The endosymbiotic acquisition of mitochondria, a pivotal event in eukaryotic evolution, significantly enhanced the capacity for energy production and sophisticated metabolic regulation, laying the foundation for the emergence of complex life. Mitochondria, which are not only the primary energy generators in modern eukaryotic cells but are also integral players in a wide array of cellular functions, orchestrate these processes through oxidative phosphorylation [43]. The mitochondrial respiratory chain, a major source of reactive oxygen species (ROS), serves as a convergence point for numerous metabolic pathways [44].
Programmed cell death, an essential mechanism for eliminating unwanted cells, is a fundamental process inherent in all living organisms [45]. Apoptosis, the most prevalent form of this cellular demise, is a meticulously regulated process that selectively removes cells during various biological events [40]. The Bax inhibitor-1 (Bi-1) protein family, a group of small transmembrane proteins, exhibits potent anti-apoptotic properties and is remarkably conserved across diverse organisms [30]. Recent studies have unveiled intricate interactions between Bi-1 and Bcl-2 protein families, demonstrating that BI-1 proteins can either amplify the anti-apoptotic activity of Bcl-2 or counteract cell death triggered by the pro-apoptotic protein Bax. Moreover, Bi-1 plays a crucial role in mediating cellular responses to both biotic and abiotic stresses in a wide range of organisms, including animals, plants, fungi, and yeast [30,46,47].
While BI-1 proteins are ubiquitous within filamentous fungi, functional characterization remains limited for most. In U. virens, UvBi-1 exerts a suppressive influence on mycelial growth and conidiation, and is indispensable for stress tolerance, cell wall integrity, the production of secondary metabolites, and the ability to cause disease [22]. In M. robertsii, a fungal pathogen, the absence of MrBi-1 resulted in impaired fungal development, attenuated virulence, and compromised heat tolerance. Inactivation of MrBi-1 diminished fungal resistance to farnesol but had no discernible effect on hydrogen peroxide resistance, suggesting that MrBi-1 contributes to apoptosis-like cell death through the endoplasmic reticulum stress signaling pathway rather than the more conventional mitochondria-dependent pathway [11]. The potential of apoptotic proteins as targets for antifungal drug development has been recognized. However, the successful implementation of this approach hinges upon a more comprehensive understanding of fungal apoptotic networks and the identification of key regulatory proteins involved in apoptosis-like cell death within these organisms.
In conclusion, this study provides evidence that the Bax inhibitor-1 family MoBi-1 encodes the putative Bax-inhibitor protein, which plays an important role in sporulation and the full virulence of M. oryzae. Our findings further reveal that MoBi-1 serves a critical regulatory role in mitochondrial energy metabolism biosynthesis. Given the localization of MoBi-1 within mitochondria and its role in ATP production, the potential involvement of MoBi-1 in the mitochondrial autophagy pathway warrants additional investigation. Notably, the experimental evidence suggests that MoBi-1 does not appear to participate in the suppression of Bax-mediated apoptotic pathways, thereby lacking intrinsic activity as a Bax inhibitor. These results extend the current understanding of the function of Bax inhibitor-1 and provide novel and important insights into the pathogenic mechanisms of M. oryzae.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Dean R.A. Talbot N.J. Ebbole D.J. Farman M.L. Mitchell T.K. Galagan J.E. Birren B.W. The genome sequence of the rice blast fungus Magnaporthe grisea Nature 200543498098610.1038/nature 0344915846337 · doi ↗ · pubmed ↗
- 2Saleh D. Milazzo J. Adreit H. Fournier E. Tharreau D. South-East Asia is the center of origin, diversity and dispersion of the rice blast fungus, Magnaporthe oryzae New Phytol.201411440145610.1111/nph.12627 PMC 426529324320224 · doi ↗ · pubmed ↗
- 3Zulkifli M. Okonkwo A.U. Gohil V.M. FDX 1 Is Required for the Biogenesis of Mitochondrial Cytochrome c Oxidase in Mammalian Cells J. Mol. Biol.20233516831710.1016/j.jmb.2023.168317 PMC 1145189737858707 · doi ↗ · pubmed ↗
- 4Wu S. Zhang Y. Xu L. Zhang H. Li Y. Yang L. Zhang Z. Zhang H. Mitochondrial outer membrane translocase Mo Tom 20 modulates mitochondrial morphology and is important for infectious growth of the rice blast fungus Magnaporthe oryzae Mol. Plant Microbe Interact.20243740741510.1094/MPMI-10-23-0168-R 38171376 · doi ↗ · pubmed ↗
- 5He Y.L. Deng Y.Z. Naqvi N.I. Atg 24-assisted mitophagy in the foot cells is necessary for proper asexual differentiation in Magnaporthe oryzae Autophagy 201391818182710.4161/auto.2605723958498 · doi ↗ · pubmed ↗
- 6Zhong K. Li X. Le X. Kong X. Zhang H. Zheng X. Wang P. Zhang Z. Mo Dnm 1 Dynamin Mediating Peroxisomal and Mitochondrial Fission in Complex with Mo Fis 1 and Mo Mdv 1 is important for development of functional appressorium in Magnaporthe oryzae P Lo S Pathog.201612 e 100582310.1371/journal.ppat.100582327556292 PMC 4996533 · doi ↗ · pubmed ↗
- 7Kou Y. He Y. Qiu J. Shu Y. Yang F. Deng Y. Naqvi N.I. Mitochondrial dynamics and mitophagy are necessary for proper invasive growth in rice blast Mol. Plant Pathol.2019201147116210.1111/mpp.1282231218796 PMC 6640187 · doi ↗ · pubmed ↗
- 8Xiao Y. Liu L. Zhang T. Zhou R. Ren Y. Li X. Shu H. Ye W. Zheng X. Zhang Z. Transcription factor Mo Msn 2 targets the putative 3-methylglutaconyl-Co A hydratase-encoding gene Mo AUH 1 to govern infectious growth via mitochondrial fusion/fission balance in Magnaporthe oryzae Environ. Microbiol.20212377479010.1111/1462-2920.1508832431008 · doi ↗ · pubmed ↗