Ultrastructural Characterization of Developmental Stages and Head Sensilla in Portici okadai, Vector of Thelazia callipaeda
Da Sun, Yang Luo, Yikang Wang, Hongle Cui, Yanting Gou, Juan Zhou, Bo Luo, Hui Liu, Rong Yan, Lingjun Wang

TL;DR
This study uses scanning electron microscopy to describe the ultrastructure of Phortica okadai across its life stages and head sensilla, aiding in species identification and olfactory research.
Contribution
The first comprehensive ultrastructural characterization of Phortica okadai developmental stages and head sensilla is provided.
Findings
Eggs of P. okadai are dark brown and cylindro-oval, lacking dorsal appendages that distinguish them from D. melanogaster.
Sexual dimorphism is observed in the length of certain sensilla types on the adult head of P. okadai.
The ultrastructural features of developmental stages can be used for species identification.
Abstract
Phortica okadai, a vector of Thelazia callipaeda, relies heavily on its chemosensory system for reproduction and population expansion. However, ultrastructural data across its developmental stages remain uncharacterized. This study employed scanning electron microscopy to observe and describe the ultrastructure of P. okadai across four developmental stages: egg, larva, pupa, and adult. It also focused on characterizing the ultrastructural features, morphometric parameters, and distribution patterns of five sensilla types on the adult head. Sexual dimorphism was observed in the length of Type II chaetica sensilla (ChII), Thin basiconic sensilla (TB), and Large basiconic sensilla (LB). The ultrastructural characteristics of P. okadai during developmental stages, such as eggs and larvae, can be utilized for species identification. Additionally, the focused investigation on the head…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
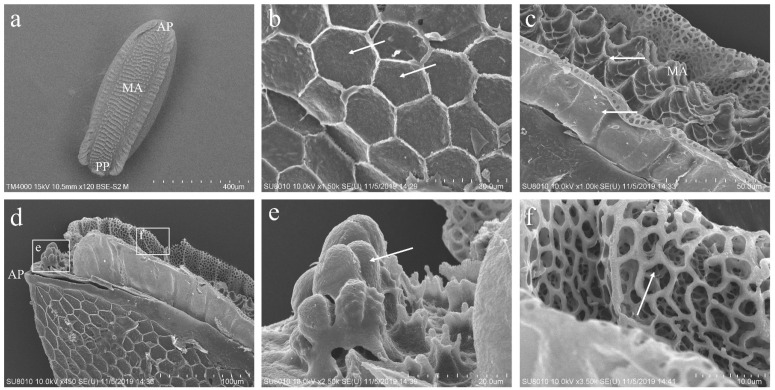 Figure 1
Figure 1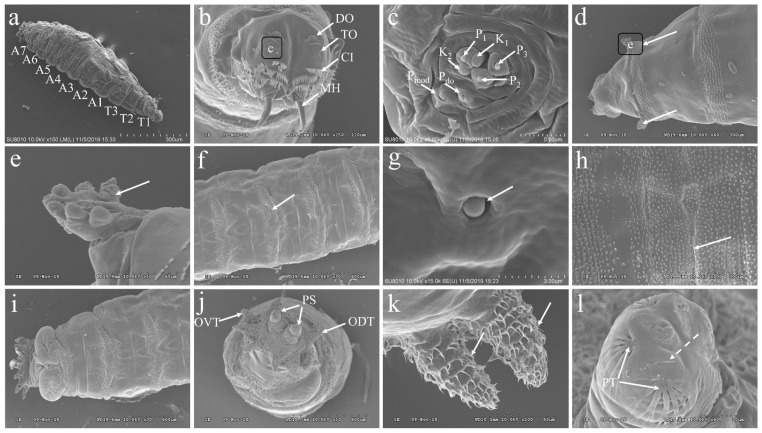 Figure 2
Figure 2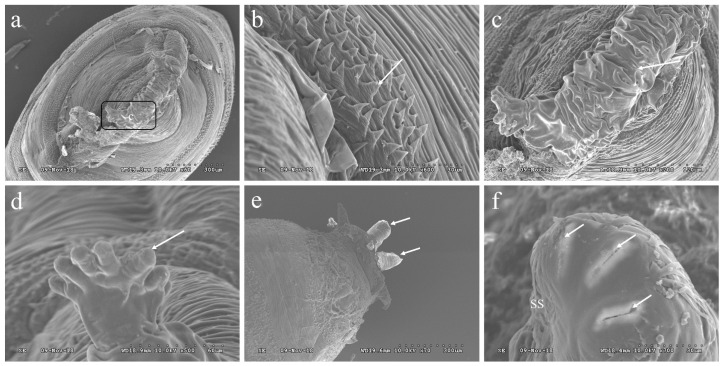 Figure 3
Figure 3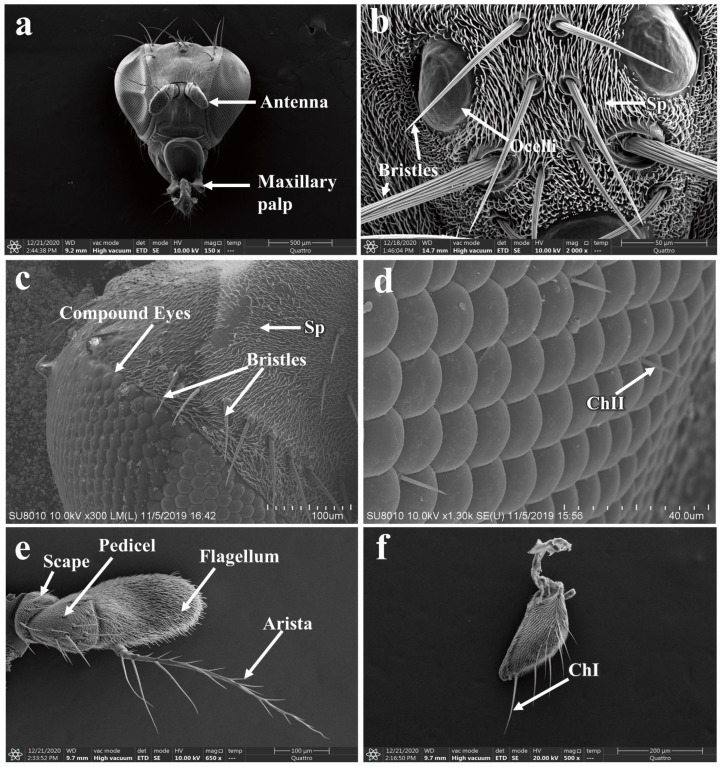 Figure 4
Figure 4 Figure 5
Figure 5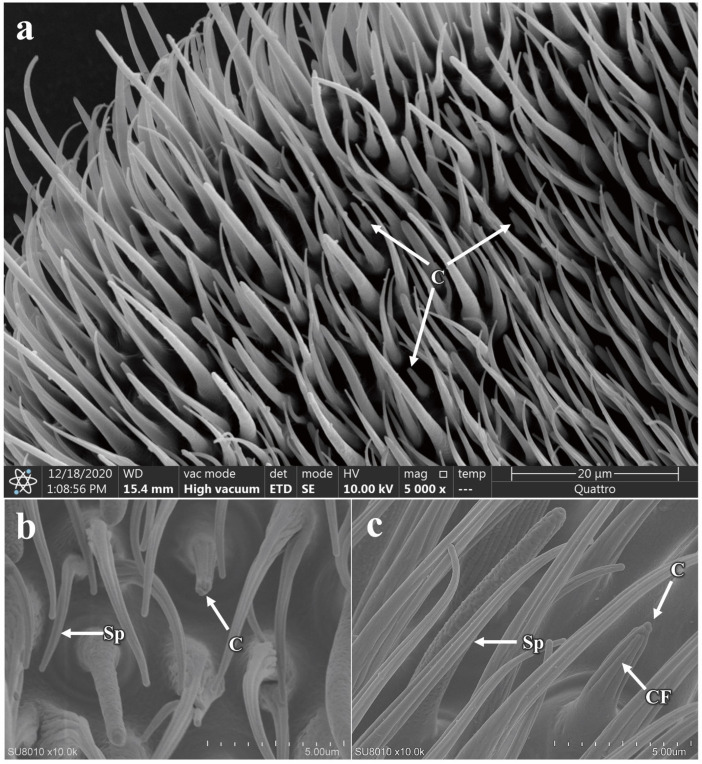 Figure 6
Figure 6 Figure 7
Figure 7 Figure 8
Figure 8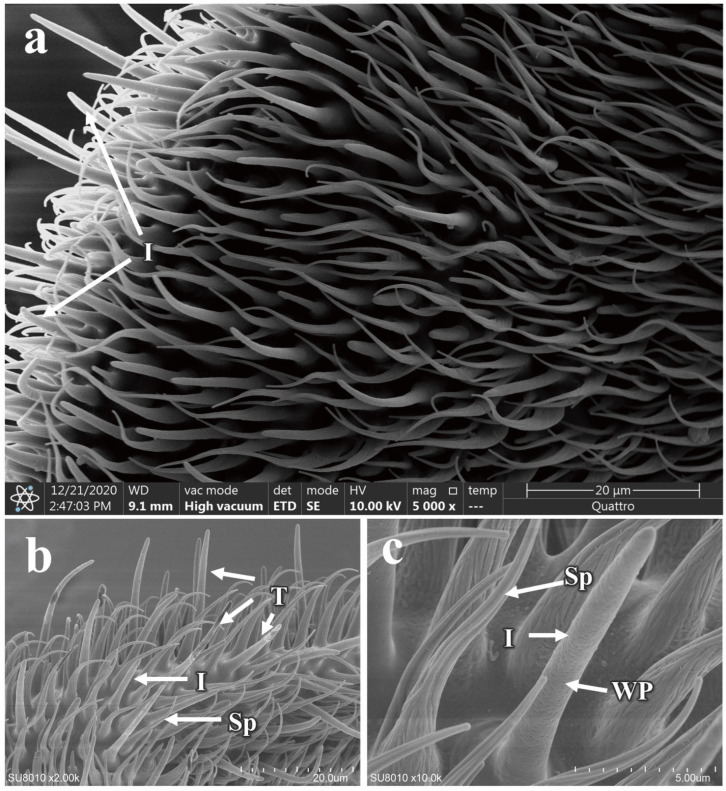 Figure 9
Figure 9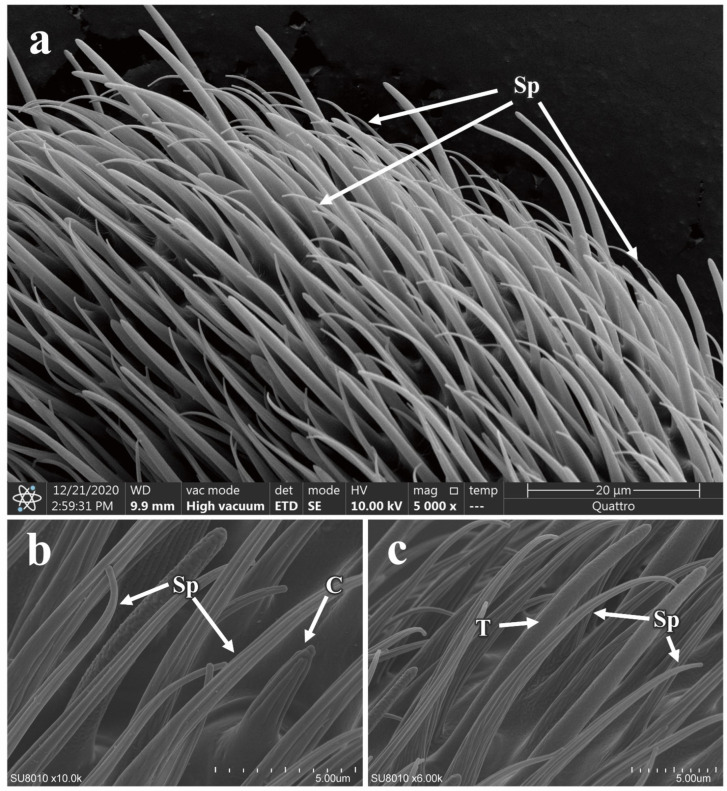 Figure 10
Figure 10 Figure 11
Figure 11- —National Natural Science Foundation Project of China
- —Science and Technology Foundation of Guizhou Province
- —Guizhou Provincial Basic Research Program (Natural Science)
- —Open Project Fund of Key Laboratory of Parasite and Vector Biology
- —Youth Science and Technology Talent Growth Project of Education Department of Guizhou Province
- —Science and Technology Foundation of Zunyi City
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsNeurobiology and Insect Physiology Research · Insect and Arachnid Ecology and Behavior · Insect Utilization and Effects
1. Introduction
Phortica okadai (Máca, 1977) (Diptera: Drosophilidae: Steganinae) is known as the only confirmed vector of Thelazia callipaeda (Railliet et Henry, 1910) (Spirurida: Thelaziidae) in China [1,2]. Meanwhile, it is also recognized as a highly polyphagous pest [3]. Not only does it inflict damage on various commercially significant fruits including pears, apples, bananas, and citrus [3], but it can also transmit thelaziasis to a variety of species such as rabbits, dogs, cats, the iconic giant pandas, and humans through lachryphagous behavior by feeding on their ocular secretions [4,5]. Thelazia callipaeda induces thelaziasis, a parasitic infection clinically manifested by lachrymation, foreign body sensation, pruritus, conjunctival follicular hyperplasia, and corneal ulceration [4,6]. Thelazia callipaeda, a zoonotic nematode, poses a significant public health concern due to its broad host range, encompassing a variety of species such as rabbits, dogs, cats, the iconic giant pandas, and, of course, humans [1,6,7]. Thelaziasis has emerged as a widespread parasitic infection, with its presence confirmed in an array of countries across the continents of Europe and Asia [7,8]. China has been recognized as the epidemiological focus of thelaziasis, accounting for the highest proportion of globally reported human cases [9], which underscores the dual significance of P. okadai in both safeguarding public health and advancing agricultural pest management strategies [10].
Conducting an in-depth study of the morphological characteristics of P. okadai at different developmental stages can facilitate species identification based on morphological features. For instance, morphological differences observed in the eggs of Anopheles culicifacies can distinguish these species from their close relatives [11]. Similarly, the larvae of D. melanogaster and Musca domestica can be differentiated based on larval structures such as the pseudocephalon and mouthparts [12,13]. Additionally, the morphological developmental mechanisms during the pupal stage of D. melanogaster can be used to distinguish it from other insect species [14]. However, there is currently a lack of detailed morphological studies on the various developmental stages of P. okadai. Notably, the adult stage of insects often exhibits heightened harmfulness due to environmental stimuli eliciting various behavioral responses mediated by diverse types of antennal sensilla, which are crucial for vital processes such as host localization, habitat selection, mate recognition, foraging, and mating [15]. For example, in Bactrocera dorsalis (Diptera: Tephritidae), sensilla trichodea and sensilla basiconica can collect olfactory information through pores in their cuticular walls, thus participating in semiochemical perception [16]. In Simulium umphangense (Diptera: Simuliidae), the sensillum coeloconicum is sensitive to temperature and humidity [17]. Meanwhile, the diversity of sensilla correlates with their ecotopes and geographic distribution [18,19]. For example, in Rhodnius brethesi (Hemiptera: Reduviidae), laboratory-reared lines have fewer thick-walled trichoidea than wild lines [20].
Furthermore, insect sensilla also exhibit sexual dimorphism, exemplified by the Bőhm bristles in Megabruchidius dorsalis, which are longer in males than in females [21]. In Sitodiplosis mosellana Géhin (Diptera: Cecidomyiidae), the male’s sensilla circumfila is highly elongated compared to the female’s, possibly to enhance pheromone detection [22]. In wild-type D. melanogaster, the distribution of sensilla trichodea on the third antennal segment is higher in males than in females [23]. Compared to females, the male P. okadai were the primary vectors of T. callipaeda [5], which is mainly because male P. okadai prefer to feed on mammalian tear secretions, also known as lachryphagy [24]. The differences in lachryphagy between males and females P. okadai are probably linked to the sexual dimorphism in their antennal sensilla. However, the sexual dimorphism in the antennal sensilla of P. okadai remains unclear. To thoroughly investigate the morphological structures of P. okadai across its developmental stages, with a focus on the detailed morphology of sensilla, scanning electron microscopy was employed in this study, which are essential for advancing research on the olfactory systems of P. okadai and improving strategies in public health management and agricultural pest control.
2. Materials and Methods
2.1. Insect Rearing
The species utilized in this study were captured from a pear orchard in Zunyi City, Guizhou Province, China, and were identified as P. okadai based on morphological characteristics such as the “shaped” black transverse bands on the dorsal sides of the third to fifth abdominal segments and three black rings on the tarsal segments. These individuals have been maintained and continuously bred under laboratory conditions for six years. The rearing conditions were optimized for these flies, utilizing naturally fermented fruits such as apples and pears, which were prepared in small sections (2 cm × 1 cm × 1 cm). The P. okadai were maintained under controlled environmental conditions: a temperature of 28 ± 2 °C, relative humidity of 75 ± 5%, and a 16:8-h light/dark photoperiod. Eggs, larvae, and pupae were collected from the rearing medium within their enclosures.
2.2. Scanning Electron Microscopy Observation
Eggs from the P. okadai rearing chambers were initially washed in 2% sodium hydroxide followed by a rinse in distilled water. Larvae and pupae were similarly rinsed with distilled water and subsequently euthanized by immersion in hot water for 5 min. The eggs, larvae, and pupae were then fixed in a mixture containing 2.5% glutaraldehyde and 0.1 M sodium cacodylate buffer (pH 7.2) at 4 °C for 24 h [25]. Following fixation, the specimens were washed twice with phosphate-buffered saline (PBS), every 10 min, and post-fixed in 1% osmium tetroxide at room temperature for 1 h. Post-fixation, they were washed again twice with PBS. Dehydration was achieved through a graded ethanol series (50%, 70%, 80%, 85%, and 90%), maintaining the specimens at each concentration for 12 h, concluding with a final placement in 100% ethanol for 24 h [26,27].
In this study, ten male and ten female adult P. okadai of similar size were anesthetized by chilling them in a −20 °C freezer for one minute. Post-anesthesia, the heads of these adults were carefully severed using fine forceps, and the antennae, along with maxillary palps, were meticulously dissected. The dissected samples were then fixed in 2.5% glutaraldehyde at 4 °C for 12 h. Following fixation, samples were rinsed in PBS (0.1 M, pH 7.2) for 10 min. To remove any adherent particles, the samples underwent a 30 s ultrasonic cleaning. The dehydration process involved a graded ethanol series starting from 70%, progressing to 80%, then 90%, and culminating in 100% ethanol.
The prepared samples were then subjected to critical point drying using a chemical dryer to ensure complete dehydration. Subsequently, they were mounted on aluminum stubs using double-sided adhesive tape. To enhance electron conductivity, the samples were coated with an 18 nm thick layer of gold–palladium alloy using a high vacuum coating system (EM ACE600, Leica Microsystems, Wetzlar, Germany). The coating process included 10 cycles, each lasting 3 min, to achieve a uniform layer. Finally, the samples were examined under a field emission scanning electron microscope (SU8010, Hitachi, Tokyo, Japan) set to operate at 3 kV. Descriptions of the eggs, larvae, and pupae were articulated using the specialized terminology of Courtney [28]. Furthermore, the classification and identification of sensilla on the antennae, eye areas, and maxillary palps adhered to the nomenclature established by Schneider [29] and Zacharuk [30].
2.3. Statistical Analysis
The sensilla located on the surface of the antennae and maxillary palps of P. okadai were meticulously identified, counted, and measured. In this study, the number of various types of sensilla per unit area was determined to assess the overall density of sensilla across the entire surfaces of the antennae and maxillary palps. For this purpose, three random regions on each antenna and maxillary palp were selected for detailed sensilla counting. The aforementioned counts were subsequently transformed into absolute densities and subjected to statistical analysis through the implementation of a t-test [21]. Some of these sensillas were evaluated using Welch’s t-test and a non-parametric Mann–Whitney U test in GraphPad Prism 9 (GraphPad Software, La Jolla, CA, USA).
3. Results
3.1. A General Description of the Stages of P. okadai
The various developmental stages of P. okadai, namely eggs, larvae, pupae, and adults, were meticulously examined and described using scanning electron microscopy. Particular attention was paid to the head during the adult stage.
3.2. The Morphology of the Eggs of P. okadai
The eggs of P. okadai are sepia with a cylindrical oval shape. Structurally, the eggs are differentiated into the anterior pole (AP), median area (MA), and posterior pole (PP) (Figure 1a). The chorion is characterized by a reticulated pattern, which results from impressions made by polygonal (pentagonal or hexagonal) follicle cells. Each follicle cell’s borders are slightly elevated, accentuating this pattern (Figure 1b, arrows indicated). The head region at the AP is underdeveloped, presenting as a series of overlapping peaks (Figure 1d,e). Within the MA, prominently raised island-like structures densely populate the polygonal configurations, and a multitude of variably shaped small pores are interspersed along the erect folds (Figure 1c,f).
3.3. The Morphology of the Larvae of P. okadai
The larvae of P. okadai display a vermiform, featuring a slender anterior and a truncated posterior. The body is segmented into 11 distinct sections, beginning with the cephalic region, followed by three thoracic segments (T1-T3) and seven abdominal segments (A1-A7) (Figure 2a). The cephalic region encompasses a pair of terminal organs (TO), dorsal organs (DO), and mouth hooks (MH), with cuticle cirri (CI) enveloping the anterior region of the cephalic lobes (Figure 2b). The terminal organs are classified into two distinct morphological categories: the papilla group (P_1–3_, P_do_), which together cover about half of the cuticle surface. These papilla extend from the cuticle, forming a shaft with a bud-like structure emerging from a cylindrical sheath, and possess a terminal pore at the bud’s apex. The P_do_ features a cylindrical segment that encloses the bud and its terminal pore. Additionally, the knob-like sensilla (K_1–2_, P_mod_) also presents a cylindrical shaft surrounding a bud-like structure (Figure 2c). Two prothoracic spiracles, symmetrically positioned between the cephalic region and the first thoracic segment, consist of a circular plane formed by seven papillary protrusions (Figure 2d,e). Small, regularly arranged spines densely populate the intersegmental regions between the thoracic and abdominal segments (Figure 2f). Additionally, transverse slits are evident (Figure 2h), and undeveloped abdominal structures are visible during this larval stage (Figure 2g). The dorsal and ventral views of the A7 display anal pads within the last abdominal segment, featuring an anal opening and two posterior spiracles flanked by spines oriented both forward and backward (Figure 2i). A7 also hosts six pairs of tubercles: inner dorsal, middle dorsal, outer dorsal, outer ventral, middle ventral, and inner ventral, each covered with distinct, layered spines. The posterior spiracles are distinguished by spiracular slits and adjacent peristigmatic tufts, characterized by long, fine, multi-branched hairs (Figure 2j–l).
3.4. The Morphology of the Pupae of P. okadai
The pupae of P. okadai feature a distinctive tubular organ at the posterior end of the first thoracic segment, known as a respiratory tubercle. This tubercle displays a globular tubular structure with segmental wrinkles, which are characteristic of the pupation process (Figure 3a,c). Surrounding the respiratory tubercle, intersegmental spines are arranged in a stepped pattern, with each spine terminating in a flat, pointed tip (Figure 3b). The structure of the anterior spiracle in the pupal stage is slightly elongated compared to its larval form, offering a more defined appearance (Figure 3d). Meanwhile, the posterior spiracle is notably more pronounced, protruding with clear structural distinctions (Figure 3e). This posterior spiracle exhibits three spiracular slits arranged in an arc and features a conspicuous spiracular scar (Figure 3f).
3.5. Morphology of the Adult P. okadai Head
The head of P. okadai predominantly consists of compound eyes, ocelli, the vertex region, antennae, and maxillary palps (Figure 4a). The vertex region is characterized by the presence of three ocelli, each equipped with bristle mechanoreceptors (Figure 4b). Additionally, the area where the compound eyes intersect with the vertex region features multiple pairs of bristles (Figure 4c). The compound eyes are composed of numerous ommatidia, and between these ommatidia, chaetica sensilla type II are distributed (Figure 4d). The antennae are segmented into three parts: the scape, pedicel, and flagellum. The flagellum bears an arista, and unlike the scape and pedicel, it is not further subdivided and is more robust in structure. The arista has a plumose appearance, tapering from the base to the tip (Figure 4e). There are sexual differences in the pedicel and flagellum of the antennae, whereas no sexual dimorphism is observed in the scape, the total length of the antennae, or the arista (Table 1). The maxillary palps possess a sensillar pit and are broadly divided into three sections, with a variety of sensilla distributed at the tip (Figure 4f).
3.5.1. Morphology and Structure of Antennal Sensilla in P. okadai
The antennae of P. okadai are densely populated with a variety of sensilla, which include spinule (Sp), basiconic sensilla (LB: Large basiconic sensilla, TB: Thin basiconic sensilla, and SB: Small basiconic sensilla), trichoid sensilla (T), intermediate sensilla (I), coeloconic sensilla (C), and chaetica sensilla (ChI, ChII). Among these, the flagellum exhibits the greatest diversity and abundance of sensilla types and numbers.
The chaetica sensilla in P. okadai are thick-walled, poreless sensilla, predominantly located on the scape and pedicel of the antennae. Based on a length criterion of 50 μm (µm), they can be categorized into two types, ChI and ChII. The sensilla have smooth shafts with pronounced curvature and are inserted into a movable socket (Figure 5a). There are no significant differences in the morphology or quantity of ChI and ChII between males and females (Table 2). However, the ChI is notably longer than the ChII (Figure 5b,c). The average length of the ChI in males is 56.53 ± 1.62 µm, while in females it is 77.34 ± 7.08 µm, indicating a significant difference (p < 0.05), primarily in the basal diameter of the sensilla (p < 0.05). The average length of the ChII in males is 31.19 ± 2.34 µm, and in females, it is 24.63 ± 0.95 µm, also showing a significant difference (p < 0.05), which is mainly reflected in the apical diameter of the sensilla (p < 0.05) (Table 2).
The coeloconic sensilla are double-walled, characterized by a stout and irregular distribution along the flagellum of the antennae (Figure 6a). They possess numerous fine, longitudinal cuticular fingers (CF) on their surface. These finger-like structures form a small cavity between their inner and outer walls, with multiple such structures creating a central cavity (Figure 6b,c). Each sensilla has a shallow, circular pit at the base and a smooth surface, distinguished by a prominent longitudinal groove that extends from the base to the tip. There are no differences in morphology or quantity between males and females for the coeloconic sensilla (additional file: Table S1). However, the average length from base to tip is significantly different between the sexes, with males measuring 5.56 ± 0.12 µm and females 4.77 ± 0.22 µm (p < 0.05). Additionally, there is a significant difference in the basal diameter of the sensilla between males and females (p < 0.05) (Table 2).
The basiconic sensilla are single-walled, multiporous, generally conical in shape, tapering from the base to the tip, with a bluntly rounded apex, and the sensilla wall is studded with wall pores (Figure 7a). Based on morphology and size, they can be classified into three subtypes: SB, TB, and LB (Figure 7b,c). The antennae are equipped with only the TB and LB. The TB are similar in overall structure to the LB but are flattened, whereas the LB are cylindrical. Both are predominantly located on the flagellum of the antennae, with no distribution observed on the pedicel and scape. There are no differences in morphology or quantity between males and females (additional file: Table S1). However, there is a significant difference in the length of the TB, with females measuring 6.72 ± 0.32 µm and males 9.66 ± 0.34 µm (p < 0.05). A similar difference is observed in the LB (p < 0.05). Regarding the sensilla diameter, the basal diameter of the TB is significantly different between females (2.79 ± 0.17 µm) and males (2.37 ± 0.08 µm), but no differences are noted at the apical end or in the basal and apical diameters of the LB (Table 2).
The trichoid sensilla are single-walled, pore-bearing, and predominantly distributed along the flagellum of the antennae. They exhibit a relatively elongated, hair-like appearance, with their surfaces densely packed with pores and a bluntly rounded tip (Figure 8a,b). There are no differences in morphology, quantity, or length between males and females (Figure 8c,d and additional file: Table S1). However, a significant difference in the basal diameter of the sensilla is observed between the sexes (p < 0.05) (Table 2).
The intermediate sensilla are positioned in length between the trichoid and basiconic sensilla (Figure 9a,b and additional file: Table S1), and like these, they have a surface densely packed with pores and a bluntly rounded tip (Figure 9c). There are no differences in morphology or quantity between males and females (Figure 9 and additional file: Table S1). The length of these sensilla in males is 9.94 ± 0.90 µm, while in females it is 13.65 ± 0.90 µm, indicating a significant difference (p < 0.05) (Table 2). Further measurements reveal that there are also significant differences in both the basal and apical diameters of the intermediate sensilla between the sexes (Table 2).
The antennae of P. okadai are densely covered with spinules across the scape, pedicel, and flagellum (Figure 10a). The cuticular surface of these spinules lacks any pores. Several longitudinal grooves extend along the surface of the spinules (Figure 10b,c), but these grooves do not penetrate the internal lumen of the spinules.
3.5.2. The Morphology and Structure of the Maxillary Palp Sensilla in P. okadai
The maxillary palps of P. okadai are cylindrical in shape, located on either side of the mandibles, and become revealed when the beak is extended (Figure 11a,b). They feature distinct sensillar pits. Compared to the antennae, the maxillary palps have a more limited variety of sensilla, primarily consisting of chaetica sensilla and basiconica sensilla, the specific morphologies of which are described above. The chaetica sensilla on the maxillary palps of P. okadai, similar to those on the antennae, can be divided into two types, ChI and ChII. There are no differences in morphology or quantity between males and females (Figure 11c,d and additional file: Table S1). However, there is a significant difference in the length of ChI between the sexes, with males averaging 56.53 ± 1.62 µm and females 77.34 ± 7.08 µm (p < 0.05), primarily in the basal diameter of the sensilla (p < 0.05). The ChII do not show a significant difference in length between males and females (p > 0.05). The basiconica sensilla on the maxillary palps exhibit greater morphological diversity compared to those on the antennae, with the presence of SB. Among them, only the TB show a significant difference in length between males and females, while the other two types do not exhibit any length differences (Figure 11e,f and Table 2).
4. Discussion
This study employed scanning electron microscopy (SEM) to conduct ultrastructural observations of P. okadai across different developmental stages. The results revealed morphological similarities to Drosophila melanogaster in eggs and larvae, with subtle distinctions [31,32,33]. For instance, two large dorsal appendages were observed near the AP on the dorsal region of D. melanogaster eggs, a unique morphological feature potentially useful for distinguishing P. okadai from its closely related species [31,32]. Notably, P. okadai larvae exhibited one fewer abdominal segment compared to D. melanogaster larvae, a characteristic that can be used for differentiation [33,34]. Pupal morphology in P. okadai showed no significant differences from other drosophilids, though many species shared a prominent respiratory tubercle surrounded by inter-segmental spines arranged in a stepped pattern [25]. The functional significance of these structures remains unclear; however, the existing literature suggests that pupal stages are characterized by exoskeletal remodeling and reorganization of internal organs [35]. Additionally, five types of sensilla were identified on the antennae and maxillary palps of P. okadai, with morphological similarities to those of D. melanogaster [36]. Sexual dimorphism was observed in the size of certain sensilla types in P. okadai between sexes.
Different sensilla types have distinct functions, and they are often distinguished by their unique ultrastructural features [16]. The trichoid and coeloconic sensilla of P. okadai are essentially identical structures to those of D. melanogaster, with the length of D. melanogaster trichoid sensilla being shorter than P. okadai (Table 2) [36,37,38]. Similarly, the chaetica sensilla of P. okadai are morphologically akin to those observed in the Earias vittella (Lepidoptera: Nolidae) [39]. These sensilla, characterized by grooves encircling the base, are notably longer than other types of sensilla. The sensilla basiconic of P. okadai, divided into SB, TB, and LB subtypes based on morphology and length, align with D. melanogaster’s classification but exhibit slightly shorter lengths compared to their counterparts in D. melanogaster [36].
Trichoid and coeloconic sensilla, presumed mechanoreceptors, adeptly convert physical stimuli like touch, pressure, movement, stretch, vibration, and contraction into electrical signals essential for regulating insect behaviors [40]. In the case of Erannis ankeraria Staudinger (Lepidoptera: Geometridae), research underscores the pivotal role of trichoid sensilla in the detection of pheromones and volatile odors, which are crucial for reproductive communication and the species’ survival [41]. Research into Diptera morphology, particularly regarding fruit flies, indicates that coeloconic sensilla may play a crucial role in flight pattern regulation [42]. On the other hand, chaetica and basiconic sensilla, likely functioning as chemoreceptors, enable insects to detect a variety of environmental chemical stimuli, including odors, playing a pivotal role in the olfactory system [38]. In Heortia vitessoides (Lepidoptera: Crambidae), chaetica sensilla are noted for their significant role in chemical information recognition and host plant exploration [43]. Similarly, in beetles, basiconic sensilla are recognized for their importance in oviposition site selection and in sensing environmental changes in chemical substances such as sugars and water, which are crucial for effective host selection [40].
The behavioral responses of insects, including feeding, mate selection, egg laying and communication, are mediated by their major sensory structures, such as the antennae and maxillary palps, which facilitate recognition, signal conversion, and transmission in response to environmental chemical and physical stimuli [21]. The investigation of insect sensilla clarifies of adaptive evolution to some extent and aids in species identification by examining sensilla morphology, for example, the four bumblebee species display differences in antennal length and sensilla types, with Bombus terrestris males having more sensilla than workers [44]. The Torymus sinensis, unlike most Chalcidoidea, has six chaetica sensilla subtypes, whereas species like Megastigmus sichuanensis have relatively small numbers of subtypes [45]. Phortica okadai antennae feature five sensilla types, analogous to the Drosophila model species D. melanogaster, but with distinct flagellum lengths: 204.7 ± 4.83 µm in females (n = 6) and 180.1 ± 9.51 µm in males (n = 6), compared to 171 ± 8 µm in female D. melanogaster (n = 5) and 158 ± 4 µm in males (n = 4).
Existing research has demonstrated that sexual dimorphism in the length of insect antenna and insect sensilla is widespread, which is primarily likely associated with behaviors such as mate-seeking, foraging for food, and environmental recognition in insects [46]. In Gephyraulus lycantha (Diptera: Cecidomyiidae), the pronounced sexual dimorphism in antennal length, with males exhibiting longer antennae and sensilla, particularly trichoid sensilla, potentially enhances male pheromone detection capabilities [47]. Male Bactrocera zonata have a greater number and concentration of trichoid sensilla types I and II on the scape, and uniquely possess trichoid sharp sensilla on the pedicel, absent in females, likely enhancing their olfactory detection of host fruits [48]. Both P. okadai and Phortica variegata, the vector of T. callipaeda in European, exhibit male-biased lachryphagous behavior, which may be linked to olfactory structures within their sensilla [4,24,49,50]. The present study investigated that relative to the gustatory organs like the maxillary palps, the olfactory sensilla on the antennae often display sexual dimorphism, with male P. okadai typically possessing longer sensilla than females, including types ChII, TB, and LB, which may be linked to lachryphagy (Table 2).
Related research indicates that the feedback mechanisms within the insect olfactory system are relatively complex, with different neurons potentially conducting information within various sensors to regulate behavioral responses [51]. The information transmission across various sensory modalities in insects is mediated by odorant molecules binding to odorant-binding proteins (OBPs) or odorant receptors (ORs), triggering signal transduction by specific neurons, such as the at1 to at4 neurons in trichoid sensilla of D. melanogaster and ab1 to ab10 neurons in basiconic sensilla [52,53,54,55,56,57,58]. While this study has not thoroughly explored neural conduction beyond the sensilla, further research is needed to understand their functional role. Investigating the insect “odor molecule-sensilla-olfactory signal transduction-behavioral response” model may yield environmentally sustainable biological control strategies beneficial for public health and agricultural pest management.
5. Conclusions
In this study, we conducted a detailed observation of the ultrastructure across various developmental stages and the head region of P. okadai using SEM. Our observations revealed that the eggs of P. okadai are dark brown and cylindrical in shape and lack dorsal appendages. The larvae can be divided into 11 segments, and the pupae have distinct ultrastructural features such as prominent respiratory tubes. We focused particularly on the adult stage, examining the head region, with an emphasis on the sensilla located on the antennae and maxillary palps. A total of five types of sensilla were identified (trichoid, intermediate, coeloconic, basiconic, and chaetic), with sexual dimorphism in the length of ChII, TB, and LB. This research fills a gap in the ultrastructural data of P. okadai across its developmental stages, laying a foundation for further investigation into the functional role of sensilla in the olfactory system.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Jin Y. Liu Z. Wei J. Wen Y. He N. Tang L. Lin D. Lin J. A first report of Thelazia callipaeda infection in Phortica okadai and wildlife in national nature reserves in China Parasites Vectors 2021141310.1186/s 13071-020-04509-033407836 PMC 7789172 · doi ↗ · pubmed ↗
- 2Otranto D. Lia R.P. Cantacessi C. Testini G. Troccoli A. Shen J.L. Wang Z.X. Nematode biology and larval development of Thelazia callipaeda (Spirurida, Thelaziidae) in the drosophilid intermediate host in Europe and China Parasitology 200513184785510.1017/S 003118200500839516336738 · doi ↗ · pubmed ↗
- 3Luo Q. Zhou Y. Wen H. Wang Z. Shen J. Ecological habits of Amiota (Phortica) okadai, a vector of Thelazia callipaeda Chin. J. Zoonoses 200824548550
- 4Shen J. Gasser R.B. Chu D. Wang Z. Yuan X. Cantacessi C. Otranto D. Human thelaziosis—A neglected parasitic disease of the eye J. Parasitol.20069287287510.1645/GE-823R.116995411 · doi ↗ · pubmed ↗
- 5Máca J. Otranto D. Drosophilidae feeding on animals and the inherent mystery of their parasitism Parasites Vectors 2014751610.1186/s 13071-014-0516-425404259 PMC 4243723 · doi ↗ · pubmed ↗
- 6Juhász H. Thury G. Szécsényi M. Tóth-Molnár E. Burián K. Deim Z. Terhes G. Human thelaziosis caused by Thelazia callipaeda eyeworm, hungary Emerg. Infect. Dis.2022282559256110.3201/eid 2812.22075736418009 PMC 9707569 · doi ↗ · pubmed ↗
- 7do Vale B. Lopes A.P. da Conceição Fontes M. Silvestre M. Cardoso L. Coelho A.C. Thelaziosis due to Thelazia callipaeda in Europe in the 21st century—A review Vet. Parasitol.201927510895710.1016/j.vetpar.2019.10895731630050 · doi ↗ · pubmed ↗
- 8Otranto D. Mendoza-Roldan J.A. Dantas-Torres F. Thelazia callipaeda Trends Parasitol.20213726326410.1016/j.pt.2020.04.01332451294 · doi ↗ · pubmed ↗