Screening conditions and constructs for attempted genetic transformation of C. elegans by Agrobacterium
Eleanor C. Warren, Andre E. X. Brown, Karen S. Sarkisyan, Ramachandran Srinivasan, Ramachandran Srinivasan, Ramachandran Srinivasan

TL;DR
The paper explores using Agrobacterium to genetically transform C. elegans in a high-throughput way, but routine success has not yet been achieved.
Contribution
A new high-throughput method for C. elegans genetic transformation using Agrobacterium is proposed and partially tested.
Findings
C. elegans can survive on Agrobacterium as a sole food source.
Conditions for Agrobacterium-mediated transformation were screened but routine gene transfer was not achieved.
A set of T-binary constructs was developed as a resource for future studies.
Abstract
Manipulating gene expression within a model organism is important for reverse genetic experimentation, and while techniques to generate transgenic C. elegans are available, they are optimised for creating individual lines. The ability to create libraries of genetically modified animals using C. elegans as a model would make new types of experiments possible and would speed up studies of animal physiology. Here, we describe a range of constructs designed to establish a high-throughput method of C. elegans transformation mediated by gene transfer from Agrobacterium. We demonstrate that C. elegans are able to survive on Agrobacterium as a sole food source, and screen conditions for Agrobacterium-mediated transformation in this organism. While we do not achieve routine gene transfer from Agrobacterium to C. elegans, we suggest that this technique has potential following further…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
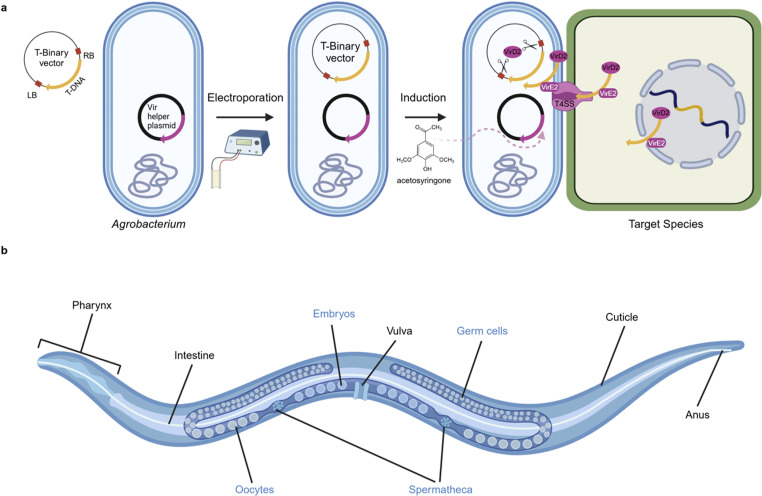 Fig 1
Fig 1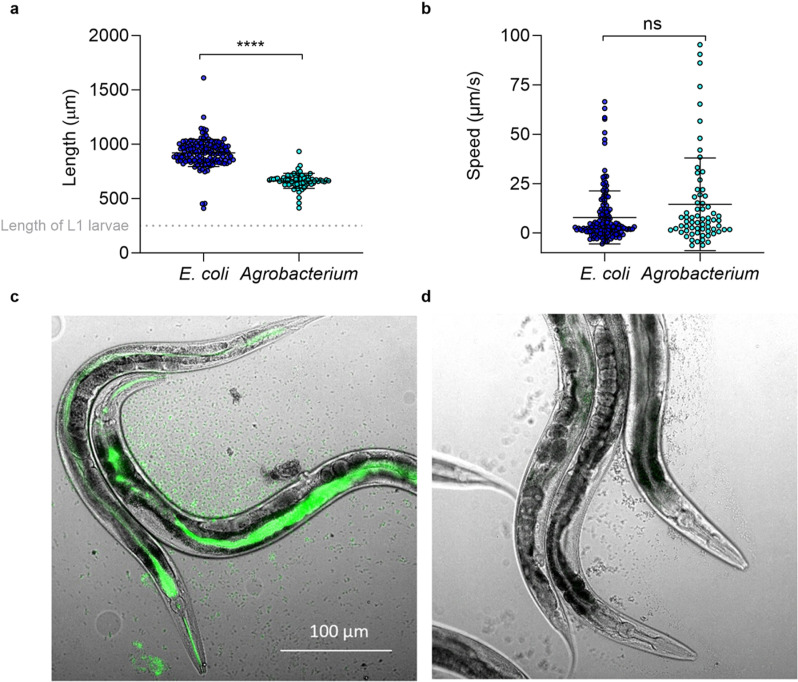 Fig 2
Fig 2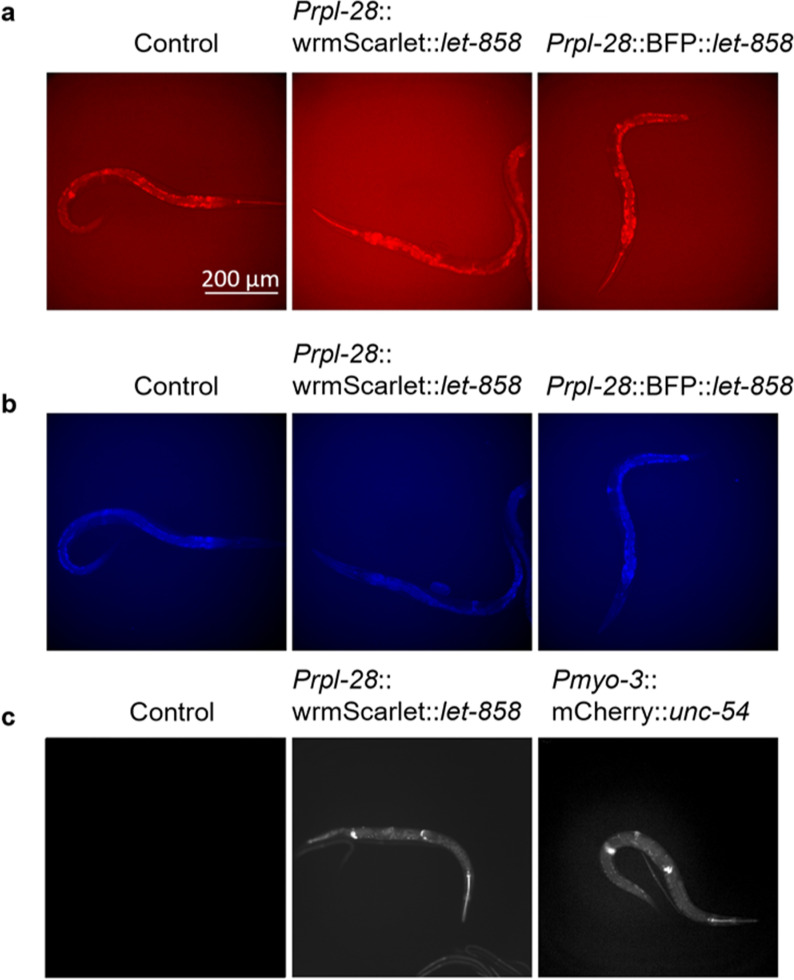 Fig 3
Fig 3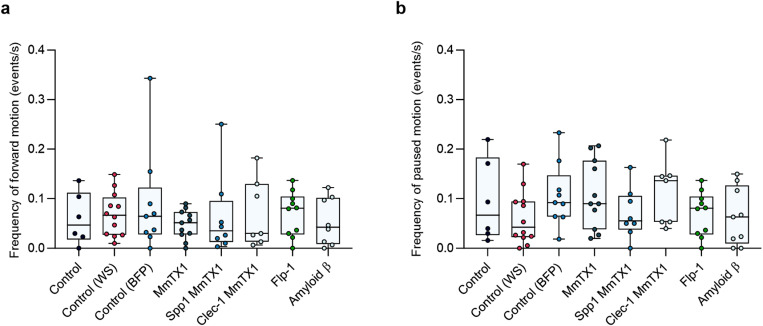 Fig 4
Fig 4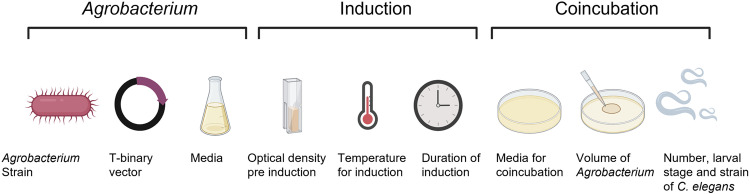 Fig 5
Fig 5- —http://dx.doi.org/10.13039/501100009187Medical Research Foundation
- —http://dx.doi.org/10.13039/501100000265Medical Research Council
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsGenetics, Aging, and Longevity in Model Organisms
Introduction
The nematode Caenorhabditis elegans is an extensively used model organism for genetic analysis, with major discoveries stemming from work on this organism. Despite the popularity of C. elegans as a research model, methods of transgenesis of this organism are limited. The most common method of transgenesis in C. elegans is microinjection of DNA into the gonad, where DNA rearranges to form inherited concatemers, or can be triggered to integrate into the genome [1]. While this technique is widely adopted and effective, it traditionally allows for the injection of only a single animal at a time, which is laborious and thus incompatible with the generation of large libraries of genetically modified animals.
Biolistic bombardment has also been employed to integrate transgenes within the C. elegans genome. This technique involves accelerating DNA-coated microparticles toward a population of worms [2,3]. Although antibiotic resistance markers can be delivered to aid identification of successful transformants, the efficiency of transgenesis using this method remains low [4]. Electroporation, a technique which is widely used for the introduction of exogenous DNA in other organisms, has been successfully applied to C. elegans for the delivery of dsRNA for RNAi silencing [5], however, attempts to achieve transgenesis of C. elegans through electroporation have resulted in only transient, non-inheritable gene expression [5,6], primarily due to challenges in targeting DNA to the germline.
Recently, a higher-throughput library transgenesis approach, termed TARDIS, has been reported [7]. This technique circumvents the low throughput nature of microinjection by transforming a single animal with a library of genetic elements, creating heritable arrays, which are subsequently integrated into the genome to yield genetically diverse individuals. TARDIS relies on the ability of C. elegans to join recombinant DNA into large and unstable extrachromosomal arrays, a process of which is not well understood mechanistically, and cannot be fully controlled by the researcher.
While genome-wide resources of constructs for use in C. elegans, such as a library of constructs for the expression of GFP-tagged proteins, have been created [8], these resources remain under-utilised due to the laborious nature of transforming each construct individually. A simpler and more direct high-throughput transgenesis protocol would be beneficial for research of animal physiology, enabling rapid creation of large libraries of genetically engineered C. elegans for high-throughput screening.
Agrobacterium-mediated transformation is the most widely used method for generating transgenic plants, and has shown success in delivering genes into organisms from other kingdoms, including yeasts [9], filamentous fungi [10], algae [11], human cells [12], and sea urchin embryos [13]. This demonstrates the potential for Agrobacterium-mediated transformation as a tool for genetic engineering across kingdoms, including transgenesis of whole animals. While the value of applying Agrobacterium-mediated transformation to C. elegans has been previously recognized [14], there are no records of this technique successfully producing transformants in this organism.
In nature, Agrobacterium introduces genes into plant genomes, which leads to tumour formation and the biosynthesis of amino acids and opines, creating a beneficial environment for the bacteria to thrive [15]. Because the DNA delivery happens in a sequence-agnostic manner, this ability to integrate genes into host genomes has been harnessed for genetic engineering by replacing tumour-inducing genes with genes of interest: any DNA sequence placed between the two 25-base-pair repeats, T-DNA borders, can be integrated into the target genome.
Agrobacterium-mediated transformation requires the expression of virulence genes which encode the machinery essential for the transfer and integration of T-DNA into the genome of the target organism*.* For the purposes of genetic engineering, a binary system has been developed where the T-DNA is present in a T-binary vector, and the virulence genes are expressed from a second plasmid called the Vir helper plasmid (Fig 1a). Induction of virulence genes by plant wound-induced phenolic compounds, such as acetosyringone, results in transfer of a single-stranded copy of plasmid-encoded T-DNA via a type IV secretion system into the target cell [16]. This system has been optimised for a variety of different plants and other species [12,17–20].
Agrobacterium-mediated transformation of C. elegans.(a) T-binary system. DNA to be integrated into the target genome is cloned between T-DNA borders (LB, and RB) in a T-binary vector, which is introduced to Agrobacterium by electroporation. Induction of virulence genes on a Vir helper plasmid is triggered with acetosyringone, resulting in cleavage of T-DNA and its delivery to the nucleus of the target species for insertion in the genome. (b) Anatomy of a C. elegans hermaphrodite, indicating reproductive components (labelled in blue), and suggested sites for Agrobacterium access (labelled in black).
The discovery that Agrobacterium is able to achieve T-DNA transfer and genome integration in human cells [12] and sea urchin embryos [13] suggests that the processes of DNA delivery and integration are conserved between plants and animals, indicating that this approach could be applied more wildly to other animal models. For Agrobacterium-mediated transformation to produce heritable germline transformants in C. elegans, the T-DNA would require access to the germline. This could be achieved through direct contact of Agrobacterium with germ cells, oocytes or sperm, or through DNA shuttling from tissues accessed by Agrobacterium, such as the intestine or vulva (Fig 1b). While heritable germline transformation would be ideal for the creation of transgenic animals, Agrobacterium-mediated transformation may also result in transient somatic transformation of the cells directly exposed to Agrobacterium, the cuticle, the lumen of the pharynx, the intestine, the vulva and the anus (Fig 1b). Here, we attempted to utilise Agrobacterium to introduce transgenic DNA into C. elegans either transiently or in a heritable manner in order to create a transgenesis approach useful for high-throughput screening.
Results
C. elegans survive with Agrobacterium as a food source
For Agrobacterium-mediated transgenesis to be possible, infection of animals with live Agrobacterium must take place. We initially examined the ability of C. elegans to feed on Agrobacterium as a sole food source and assessed both length and speed of worms fed either on E. coli (OP50), the diet routinely fed to C. elegans in the laboratory, or on Agrobacterium tumefaciens strain LBA4404 (referred to as Agrobacterium). Adult C. elegans fed on Agrobacterium were shorter in length (Fig 2a, p < 0.0001, Unpaired Mann-Whitney test), while speed was not significantly different (Fig 2b, Unpaired Mann- Whitney test). C. elegans were able to survive on Agrobacterium, with survival comparable to C. elegans on E. coli (S5 Fig). C. elegans were able to produce viable progeny on Agrobacterium, with eggs visible 3 days after refeeding L1s, and F1 progeny hatching 4 days after refeeding (S5 Fig). Eggs from the F1 and F2 generation were laid earlier in C. elegans fed Agrobacterium (induced or uninduced) compared to C. elegans fed E. coli (p ≤ 0.01 Kruskal–Wallis test with Dunn’s post hoc test), however, no attraction to Agrobacterium over E. coli was observed from a chemotaxis assay (S5 Fig, p > 0.05, Unpaired Mann-Whitney test). Additionally, we fed C. elegans Agrobacterium expressing GFP and observed fluorescence within the lumen of the pharynx and within the intestine of the worm both in the P0 generation and in the progeny (Fig 2c).
Appearance of C. elegans fed on Agrobacterium.Adult C. elegans fed on Agrobacterium or E. coli were monitored for (a) length and (b) speed (Mann–Whitney test, n > 72. Error bars represent mean + /- SD). Widefield microscopy of C. elegans fed (c) GFP expressing Agrobacterium or (d) non fluorescent Agrobacterium.
Attempt to generate fluorescent C. elegans through the introduction of fluorescent constructs using Agrobacterium-mediated transgenesis
With the aim of introducing detectable transgenic gene expression into C. elegans using Agrobacterium-mediated transformation, we fed worms Agrobacterium containing T-binary vectors with T-DNA comprising transcriptional units designed to drive the expression of fluorescent proteins in C. elegans. Transcriptional units designed with a C. elegans specific promoter (Prpl-28) and 3’UTR (let-858 3’ UTR) expressing either wrmScarlet or mTagBFP2 (referred to herein as BFP) were introduced into T-binary vectors and delivered to Agrobacterium via electroporation. If these constructs are incorporated into worms, this promoter should drive expression ubiquitously in the progeny, alternatively insertion into somatic cells would be expected to produce mosaic animals with fluorescence in specific targeted tissues [21].
Successfully transformed Agrobacterium were induced to express virulence genes, with the aim of triggering transport, insertion, and expression of T-DNA from within C. elegans cells. C. elegans from different life stages (from L1 to adult) were exposed to induced Agrobacterium, and fluorescence was monitored both in the parental generation (P0) and in the progeny (F1). Strong levels of autofluorescence were observed in worms fed Agrobacterium, however no additional fluorescence was reproducibly observed in worms fed Agrobacterium designed to introduce wrmScarlet (Fig 3a) or BFP (Fig 3b) expression.
Autofluorescence of Agrobacterium infected C. elegans.C. elegans fed on Agrobacterium control or Agrobacterium containing T-binary vectors (Prpl-28::wrmScarlet::let-858 or Prpl-28:BFP::let-858) induced to transfer T-DNA to host, were monitored for fluorescence using widefield microscopy in the (a) DsRed channel or (b) blue channel. (c) Under certain conditions C. elegans fed Agrobacterium with the T-binary vector Prpl-28::wrmScarlet::let-858 or Pmyo3::mcherry::unc-54 showed strong fluorescence in specific regions.
On two separate occasions, worms fed Agrobacterium with T-DNA designed to introduce wrmScarlet expression exhibited remarkable fluorescence (Fig 3c). This pattern of atypically bright fluorescence was also observed on 10 occasions with a construct (Pmyo3::mCherry::unc-54), where the fluorescent gene was designed to contain introns to preclude any expression of the fluorescent gene by the Agrobacterium (Fig 3c). The conditions used to obtain these results are highlighted in S1 Table. On these twelve occasions, this phenotype was displayed by all C. elegans observed, including in the P0 and the F1 generation (S3 Fig), however this bright fluorescence was not consistently reproducible across different experiments even when these conditions were reproduced. This bright fluorescence appeared to localise to the pharynx, the vulva, and two regions approximately 100 µm anterior and posterior to the vulva. Additionally, fluorescence was sometimes observed in the intestine. In order to assess the background fluorescence of Agrobacterium, cells with and without T-binary vectors were monitored for fluorescence (S2 Fig). Presence of T-DNA designed for the expression of fluorescent proteins in C. elegans did not lead to increased fluorescence of Agrobacterium, both in constructs with introns (encoding mCherry) or without introns (encoding wrmScarlet), indicating that fluorescent proteins with *C. elegans-*specific promoters were not expressed by Agrobacterium. Agrobacterium did, however, display higher autofluorescence when grown on certain media types, such as Todd Hewitt (TH) as compared to LB-Nematode Growth Media agar (LB-NGM) (S2 Fig).
Phenotyping C. elegans behaviour fed Agrobacterium containing test constructs
As an additional metric to assess Agrobacterium-mediated gene delivery to C. elegans, we selected a range of genes that – if exogenously expressed in C. elegans – would be expected to result in severe paralysis. Genes chosen for inclusion within T-DNA were the neuropeptide flp-1 (dna1959), and Amyloid-β (Aβ1–42) (dna1977), both of which have been demonstrated to cause inhibited movement or paralysis when expressed in C. elegans [22,23], as well as MmTX1, a peptide from coral snake venom which has been reported to paralyse worms when applied topically [24]. These genes were included as transcriptional units within T-DNA in Agrobacterium, with the sequence for MmTX1 either alone (dna1948) or with signal peptide sequences, clec-1 (dna1947), or Spp1(dna1946) [25]. Controls of wrmScarlet (WS) (dna1401), BFP (dna1621) or no plasmid were included. C. elegans larvae were fed on these strains of induced Agrobacterium, and at adulthood their behaviour was monitored to assess the frequency of forward motion and frequency of pausing as a read out of paralysis. We found no significant differences in forward motion (Fig 4a), or pausing (Fig 4b) under any condition tested, suggesting that the genes of interest were not successfully expressed, at least not at levels sufficient to paralyse worms.
Locomotion of C. elegans fed on Agrobacterium control strain, or strains containing T-binary vectors engineered to paralyse C. elegans.(a) Frequency of forward motion, (b) frequency of paused motion (Kruskal–Wallis test with Dunn’s post hoc test, n ≥ 6, boxes represent IQR, line shows median value). Each independent replicate represents an average of 3 worms per well.
Screening conditions for Agrobacterium induction and coincubation with C. elegans
We screened several parameters of the transformation protocol trying to achieve successful gene delivery, including strain of Agrobacterium and T-binary vector used, induction conditions, and conditions for C. elegans coincubation with Agrobacterium (Fig 5).
Schematic of the transformation protocol parameters varied during testing for C. elegans transformation using Agrobacterium.Expanded details of experimental conditions are provided in S1 Table.
Since infection by Agrobacterium is likely necessary for bacterial attachment and T-DNA transfer, we also investigated whether some of the available C. elegans mutant strains would be potentially susceptible to infection. Strains tested included a mutant defective in grinding bacteria (phm-2(ad597) I.), which has previously been shown to allow live bacteria to accumulate in the intestine [26], a mutant with a disrupted cuticle integrity (bus-17(e2800) X.) [27], and a mutant with intestinal invaginations (bbln-1(mib70) X.) [28]. Given that the vulva may serve as an entry point for Agrobacterium, we also explored Agrobacterium-mediated transgenesis in C. elegans using the him-5(e1490) V. strain, which exhibits a high incidence of males. Our aim was to facilitate mating between male worms and hermaphrodites, thereby enhancing agrobacterial entry through this route and potentially increasing access to the germline. However, none of the tested conditions or their combinations resulted in successfully transformed C. elegans in our hands. A summary of all the conditions tested is shown in S1 Table.
T-binary vectors for use in C. elegans
A range of T-binary vectors for use in C. elegans have been generated (Table 1). DNA sequences are available (S1 Fig). We employed modular cloning vectors designed for Golden Gate construction of multigene constructs [29,30]. We selected Modular cloning vectors MoClo level 1 position 1 (pICH47732), position 2 (pICH47742) and position 3 (pICH47751) (for singular transcription units) or MoClo level 2 (pAGM4723) (for multiple transcription units) which contain origins of replication for E. coli and Agrobacterium, left and right borders for T-strand transfer, and Type IIS restriction sites compatible with Golden Gate cloning [29,30]. All T-DNA sequences were designed with well documented promoter and 3’UTR pairs for ubiquitous expression, Prpl-28 and let-858 3’UTR [31], Peft-3 and tbb-2 3’UTR [32], and Prps-27 and unc-54 3’UTR [33] or targeted expression Pmyo-3 and unc-54 3’UTR [34]. A range of fluorescent proteins were chosen: wrmScarlet, BFP, mCherry, GFP and eYFP, to enable visual identification of transformed organisms. Transcription units containing the genes for flp-1, Amyloid-β (Aβ1–42) and MmTX1 (with and without secretion signals) were constructed, cloned into the backbones alone or in combination with wrmScarlet. Finally T-DNA sequences containing a transcription unit designed to express a puromycin resistance gene [35] were created, and are available in combination with other transcription units in MoClo level 2.
Table 1: Constructs for testing Agrobacterium-mediated transformation.
Discussion
Agrobacterium have previously been shown to transfer genes to a range of organisms under a variety of conditions, suggesting that Agrobacterium-mediated transformation could be successfully applied to a wide range of species. In this study we have designed a range of plasmids for use in Agrobacterium-mediated transformation of C. elegans, a technique which could vastly increase the speed and throughput of transgenesis of nematodes. We have systematically varied conditions for Agrobacterium growth, induction, and coincubation with C. elegans, however, have not successfully generated routine transgenesis of nematodes using this method. Here we are reporting our results and test conditions, and are sharing the plasmids created for Agrobacterium-mediated transformation.
C. elegans is a widely used model, with over 1000 labs worldwide studying this organism. Transgenic C. elegans are most commonly created via individual microinjection of worms [36]. Currently, large-scale forward genetic screens are possible where mutagenesis or RNAi is used to disrupt gene function, and phenotypes are subsequently identified. However, the ability to generate vast numbers of C. elegans-expressing libraries of functional transgenes, through a technique such as Agrobacterium-mediated transformation, would enable large scale screens of mutants expressing endogenous, or exogenous proteins or peptides [37,38]. Given the high efficiency of Agrobacterium transformation in plant systems, as demonstrated by the ability to achieve near-simultaneous delivery of over 20 unique T-DNAs to a given plant cell [39] we believe Agrobacterium holds significant potential for use in C. elegans transformation.
Here we have failed to generate transgenic C. elegans using Agrobacterium-mediated transgenesis, however in some conditions we have observed intense fluorescence in specific regions of the worms, such as the vulva and two regions anterior and posterior to the vulva (Fig 3c and S4 Fig). While this fluorescence is noticeably brighter, it is located in comparable regions where autofluorescence has been observed in control Agrobacterium lacking a T-binary vector (Fig 3a). We have not observed this same pattern of autofluorescence in C. elegans fed E. coli (S4 Fig). Using GFP-expressing Agrobacterium we demonstrated that the Agrobacterium localise within the pharynx and intestine of C. elegans with no noticeable accumulation within these regions where we have observed fluorescence (Fig 2c). This indicates that the observed fluorescence is not caused by Agrobacterium accumulation, but may result from an interaction with the bacteria, complicating the interpretation of the red fluorescence as a potential T-DNA expression signal.
Genetic modification of C. elegans via traditional methods is well established, thus there are many well described regulatory elements for engineering targeted gene expression in this organism. For the generation of functional transcription units to be delivered as T-DNA from T-binary vectors we selected a range of promoters for targeted (Pmyo-3) [39], or ubiquitous (Peft-3, Prpl-28 and Prps-27) expression [1,32,33]. Well established 3’ UTRs were chosen for ubiquitous expression (unc-54, tbb-2) [1], or permissiveness for germline expression (tbb-2, let-858) [40,41]. Genes chosen for easy to observe phenotypes (either fluorescence or behavioural paralysis) were chosen for expression from T-DNA, and in order to aid identification of successfully transformed nematodes, a puromycin resistance gene was designed to be ubiquitously expressed from an additional modular transcription unit [35]. The constructs designed here (Table 1) are suitable for in vivo genetic transformation of C. elegans animals, or transformation of ex vivo cultured C. elegans germ cells and should provide a useful resource for optimizing Agrobacterium-mediated transformation of C. elegans.
Most Agrobacterium-mediated transformation protocols share common features such as induction of virulence genes with acetosyringone and co-cultivation within a narrow temperature range of 22–26°C [17,42–44]. However, Agrobacterium-mediated transformation with sea urchin embryos has been successful in the absence of phenolic inducer or defined induction media [13] and co-incubation of HeLa cells with Agrobacterium has been successfully carried out at 37°C [12], much higher than has been established for plant protocols. This suggests that optimal conditions for Agrobacterium-mediated transformation differ according to species, and indicates that modification of existing protocols could result in successful T-DNA transfer from Agrobacterium to C. elegans.
Following the discovery of Agrobacterium-mediated transformation, and widespread use of this technique for generating genetically modified plants, optimised Agrobacterium strains have been developed to enhance transformation efficiency. Since certain strains appear to have differing efficacies in different species [45], we selected a range of Agrobacterium strains for testing with C. elegans: LBA4404 [46], C58C1[47], GV3101 [48], AGLO [49], EHA105 and A281 [50]. This includes strains used in transformation of HeLa (C58C1) and sea urchin embryos (GV3101), however, transformation into C. elegans was unsuccessful with all of these strains. For future optimization of Agrobacterium-mediated transformation, modifications to widely used strains could be made by improving the attachment of Agrobacterium to host cells. Work into enhancing attachment of Agrobacterium to non-natural hosts has been carried out by engineering adhesins to the surface of Agrobacterium [51], applying similar techniques for attachment to C. elegans cells could result in successful infection and T-DNA transfer. Furthermore, strains of Agrobacterium with reporters for virulence gene induction such as lacZ [52] or GFP [53], could be beneficial for optimizing induction conditions.
In addition to the strains of potentially-susceptible C. elegans mutants tested here, other C. elegans strains could be worth investigating, such as immunocompromised mutants (e.g., sek-1(ag1) or nsy-1(ag3)) with reduced innate immunity [54]. Strains of C. elegans such as vitellogenin mutant (*vit-1(ok2616)*X) with reduced eggshell integrity [55] could be useful to enable Agrobacterium to access and infect the germline. Alternatively, exogenous chemicals such as caffeine [56] could be used to physically permeabilise the eggshell, allowing Agrobacterium to transform cells of the germ line, thereby enabling heritable genetic modifications.
Several other factors in the procedure for Agrobacterium-mediated transformation, aside from Agrobacterium and host strain*,* have been shown to influence gene transfer efficiency. Such essential conditions include, the pH of the co-cultivation medium [57,58], the cell density of Agrobacterium [58,59], and media used for both induction and co-cultivation [60]. We systematically altered these conditions along with other variables in the Agrobacterium-mediated transformation protocol (S1 Table), with the aim of developing a procedure compatible with C. elegans. While we didn’t succeed in developing an Agrobacterium-mediated transformation protocol for C. elegans, we hope sharing details of our optimization strategy may prove useful for others.
Materials and methods
Construct construction
Sequences encoding genes, promoters and terminators were ordered as gene fragments from Twist Biosciences, or Integrated DNA Technologies (IDT), and cloned by golden gate assembly into MoClo level one plasmids (pICH47732: position 1, pICH47742: position 2 or pICH47751: position 3) before assembly of multiple transcription units into MoClo level 2 (pAGM4723). Except for Pmyo-3::mCherry::unc-54 in L1 which was generated using Gibson assembly with DNA amplified from pCFJ104 [34], and MoClo L1 (pICH47732).
Bacteria strains and vectors
Agrobacterium tumefaciens strain LBA4404 (ThermoFisher Scientific, 18313015) was used as a host for all experiments shown in this paper. During screening, a selection of Agrobacterium strains were tested (EHA105, GV3101, A281, AGLO and C58), all strains were maintained with Streptomycin 100 µg/ml and Rifampicin 25 µg/ml. Agrobacterium competent cells were transformed with T-binary plasmids according to manufacturer instructions (ThermoFisher, Scientific 18313015), and selected for using 50 µg/ml Carbenicillin (Level 1 backbones) or 50 µg/ml Kanamycin (level 2 backbones). PCR using T-DNA specific primers was used to confirm successfully transformed Agrobacterium colonies*.* Untransformed Agrobacterium lacking any T-binary vector was used as a negative control.
C. elegans strains
C. elegans strains N2, phm-2(ad597) I. (DA597), bus-17(e2800) X. (CB6081) and him-5(e1490) V. (CB4088) were obtained from the Caenorhabditis Genetics Center (CGC). C. elegans strain bbln-1(mib70) X. was generously provided by Mike Boxem (Utrecht University).
Agrobacterium-mediated transformation
A standardized protocol is available at dx.doi.org/10.17504/protocols.io.5qpvo9emzv4o/v2. Agrobacterium was streaked to LB or Yeast Extract Peptone (YEP) agar plates containing appropriate antibiotics to maintain plasmid vectors (100 µg/ml streptomycin, 50 µg/ml Carbenicillin or Kanamycin and 25 µg/ml Rifampicin), and plates were incubated for 3 days at 28°C. A single colony of Agrobacterium was used to inoculate 20 ml YEP media (with appropriate antibiotics) which was incubated overnight at 28°C and 250-rpm. Overnight cultures were diluted 1:100 in 20 ml YEP media or AB sucrose media ((0.5% sucrose, 3% K_2_HPO_4_, 1% NaH_2_PO_4_, 1% NH_4_Cl, 0.145% MgSO_4_, 0.15 KCl, 0.01% CaCl_2_, and 0.0025% FeSO_4_-7H_2_O, 2 mM NaPO_4_)(with appropriate antibiotics), and these cultures were incubated at 28°C and 250-rpm until the Agrobacterium reached an OD600 of 0.3. Agrobacterium cells were pelleted by centrifugation (5 min 4000g), before resuspension in 1 ml induction medium containing acetosyringone (100 µM or 200 µM), and shaking at 50-rpm at room temperature for varying durations (S3 Fig). 0.5 ml of Agrobacterium in induction media was added directly to LB-NGM (or alternative) plates with appropriate antibiotics. A mixed population of worms (containing C. elegans at different life stages (from L1 to adult), were washed three times in M9 buffer before 15 worms were transferred, by pipetting, to plates with Agrobacterium. Alternatively, C. elegans were bleach synchronised (dx.doi.org/10.17504/protocols.io.2bzgap6) and either L1s were added to Agrobacterium directly, or worms were refed until L4 stage before washing three times in M9 buffer and adding to Agrobacterium. Plates were incubated at 20 degrees for 4–10 days, such that the progeny of the exposed C. elegans can be observed. C. elegans fluorescence was observed for 100s of nematodes directly on plates using an Olympus U-RFL-T microscope, or for 10’s of worms on microscope slides using a Olympus IX70 fluorescence microscope. To anaesthetise C. elegans to prevent them moving during microscopy, C. elegans were transferred to microscope slides with 3% agar pads and 1ul of 25 mM sodium azide.
Multiple different conditions were tested for Agrobacterium-mediated transformation (S1 Table). Multiple Agrobacterium strains (LBA4404, EHA105, GV3101, A281, AGLO, C58), and T-binary vectors were utilised. Various conditions for Agrobacterium induction were screened, such as the optical density (OD600) of Agrobacterium for induction, the density of Agrobacterium in induction medium, the concentration of the phenolic inducer acetosyringone, the duration of induction, and the presence of antibiotics. Multiple media types were tested for co-incubation of Agrobacterium and C. elegans. These were LB-NGM (1% tryptone, 0.5% yeast extract, 1% NaCl, 1mM CaCl_2_, 1mM MgSO_4_, 1mM KPO_4_, 1.7% BioAgar, 5 µg/mL Cholesterol), LB (1% tryptone, 0.5% yeast extract, 1% NaCl, 1.5% agar), Low salt LB (1% tryptone, 0.5% yeast extract, 0.05% NaCl, 1.5% BioAgar), SOB (2% tryptone, 0.5% Yeast extract, 0.05% NaCl, 0.0186% KCl, 0.203% MgCl, 0.24% MgSO_4_, 1.5% BioAgar), YEP (1% Bacto peptone, 1% Yeast extract 0.5% NaCl, 1.5% BioAgar), Terrific (2% Tryptone, 2.4% Yeast extract, 0.4% Glycerol, KPO_4_, 1.7 mM KH_2_PO_4_, 7.2 mM K_2_HPO_4_, 1.5% BioAgar), 2x Yeast extract Tryptone (2xYT) (1.6% Tryptone, 1% Yeast extract, 0.5% NaCl, 1.5% BioAgar), Yeast Mannitol (YM) (0.04% Yeast extract, 1% mannitol 1.7 mM NaCl, 0.8mM MgSO_4_, 2.2 mM K_2_HPO_4_, 1.5% BioAgar), liquid Induction Media (IM)(1% NH_4_Cl, 0.145% MgSO_4_, 0.15 KCl, 0.01% CaCl_2_, and 0.0025% FeSO_4_-7H_2_O, 2 mM NaPO_4_ (pH 5.6), 50 mM 2-(4-morpholino)-ethane sulfonic acid (MES), 0.5% glucose, 100 μM acetosyringone), Induction Media agar (IM + 1.5% BioAgar), Dulbecco’s Modified Eagle Medium (DMEM Gibco +1.5% BioAgar), Yeast Peptone (YP) (2% Bacto peptone, 1% Yeast extract, 1.5% BioAgar). All media was supplemented with 5 µg/ml cholesterol. The concentrations of streptomycin and carbenicillin in these plates were varied, as was the presence of surfactant and the hydration of the plates. For LB-NGM plates, the brand of LB and Agar was varied as were the potassium phosphate salts added. Additionally, the strain of C. elegans, number of worms, and temperature of incubation were systematically varied.
Behavioural feature extraction
A detailed protocol is available at dx.doi.org/10.17504/protocols.io.n92ldro9og5b/v1. Briefly, tierpsy tracker was used to obtain behavioural features for worms in wells of a 96 well plate [61–63]. We extracted features describing morphology, “length_50th” (median worm length), and movement, “speed_50th” (median worm speed), “motion_mode_forward_frequency” (frequency of forward motion in events/second) “motion_mode_pasued_frequency” (frequency of paused motion in events/second).
C. elegans progeny, lifespan and chemotaxis assays
A detailed protocol is available at dx.doi.org/10.17504/protocols.io.e6nvwbx59vmk/v2. For progeny assays, approximately 50 synchronized L1 N2 C. elegans were transferred onto 90 mm LB-NGM plates seeded with 1 mL of either E. coli (OP50), uninduced Agrobacterium, or induced Agrobacterium, all normalized to an OD600 of 1.8. Plates were monitored every 24 hours to record the number of eggs and progeny visible per plate. Data collection continued until F2 generation eggs were observed (day 6).
For lifespan assays, approximately 50 synchronized L4 N2 C. elegans were transferred onto LB-NGM plates supplemented with 400 µM 5-Fluorodeoxyuridine and seeded with 1 mL of E. coli (OP50), uninduced Agrobacterium, or induced Agrobacterium, all normalized to an OD600 of 1.8. C. elegans were monitored daily, and individuals were classified as dead if they failed to respond to a gentle touch to the head using an eyebrow pick.
Chemotaxis assays were performed on 90 mm LB-NGM plates seeded with 100 µL of E. coli (OP50) or induced Agrobacterium at opposite ends of the plates. Both cultures were normalized to an OD600 of 1.8. Approximately 40 synchronized L4 C. elegans were placed at the center of each plate. After 4 hours, the number of worms on each bacterial lawn was recorded and the Chemotaxis Index (CI) was calculated as (number of worms on E. coli - number of worms on Agrobacterium/ total number of worms).
Statistics
The distribution of all experimental data was tested using the D’Agostino and Pearson omnibus normality test. Two-tailed non-parametric Mann-Whitney tests were used to assess significance between independent samples from two groups not showing a Gaussian distribution. The Kruskal-Wallis test with Dunn’s post hoc test was used to test significance between three or more independent groups of data not showing a Gaussian distribution. Statistical analysis was carried out using GraphPad Prism Software.**** indicates p ≤ 0.0001, p ** indicates p ≤ 0.01, * indicates p ≤ 0.05. Where no significance is indicated in a figure, p > 0.05.
Supporting information
S1 FigGenbank sequence files for all constructs for testing Agrobacterium-mediated transformation.DNA Sequence files for constructs described in Table 1.(ZIP)
S2 FigAutofluorescence of Agrobacterium and C. elegans on LB-NGM or Todd Hewitt media.(a) Autofluorescence of Agrobacterium containing different T-binary vectors grown on LB-NGM or Todd Hewitt (TH) agar was assessed by pelleting a known number of cells and visualizing the pellet in an Eppendorf tube using widefield fluorescence microscopy. Autofluorescence of C. elegans fed Agrobacterium grown on LB-NGM or TH agar was visualized using widefield fluorescence microscopy, using the DsRed channel. (b) Quantification of autofluorescence from Agrobacterium and C. elegans grown on LB-NGM or TH agar (Mann–Whitney test, n = 4, error bars represent mean + /- SEM).(TIF)
S1 TableExperimental parameters for attempted C. elegans transformation using *Agrobacterium.*C. elegans fed Agrobacterium carrying T-binary vectors engineered for fluorescence expression in worms occasionally exhibited fluorescence in specific body regions, which appeared noticeably brighter than the typically observed autofluorescence. Experimental conditions associated with these phenotypes are highlighted in green, and representative images are displayed in S3 Fig.(TIF)
S3 FigRepresentative fluorescence microscopy images of C. elegans from the highlighted conditions in S1 Table.Images were captured using widefield fluorescence microscopy in the DsRed channel. Wild-type worms without Agrobacterium treatment served as controls, with three representative images shown.(TIF)
S4 FigFluorescence observed in C. elegans following exposure to *Agrobacterium.*C. elegans fed Agrobacterium carrying T-DNA engineered for fluorescence expression occasionally exhibited atypically bright fluorescence in the vulva and in two regions approximately 100 µm anterior and posterior to the vulva. (a) Fluorescence was detected in the vulva (labeled with *), as well as in two lateral regions (labeled with x), following exposure to Agrobacterium strain LBA4404 containing either the Pmyo-3::mCherry::unc-54 or Prpl-28::wrmscarlet::let-858 T-binary vector. Control worms exposed to Agrobacterium lacking the T-binary vector are also shown. (b) Autofluorescence observed in C. elegans fed E. coli (OP50) or Agrobacterium lacking the T-binary vector is compared to the fluorescence observed in C. elegans fed Agrobacterium carrying the Prpl-28::wrmscarlet::let-858 construct. Images were captured using widefield fluorescence microscopy in the DsRed channel and brightfield microscopy.(TIF)
S5 FigComparison of C. elegans lifespan and fecundity on Agrobacterium and E. coli.(a) Survival curves of C. elegans fed E. coli (OP50), uninduced Agrobacterium, or induced Agrobacterium (Kruskal–Wallis test with Dunn’s post hoc test, n = 6). (b) Quantification of eggs and progeny laid over time by C. elegans fed E. coli (OP50), induced Agrobacterium or uninduced Agrobacterium (Kruskal–Wallis test with Dunn’s post hoc test, n = 6). (c) Chemotaxis index of C. elegans comparing E. coli (OP50) against induced Agrobacterium (Mann–Whitney t test, n = 6. Box represents IQR, line shows median value).(TIF)
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Nance J, Frøkjær-Jensen C. The Caenorhabditis elegans Transgenic Toolbox. Genetics. 2019;212(4):959–90. doi: 10.1534/genetics.119.301506 31405997 PMC 6707460 · doi ↗ · pubmed ↗
- 2Isik M, Berezikov E. Biolistic Transformation of Caenorhabditis elegans. In: Sudowe S, Reske-Kunz AB, Editors. Biolistic DNA Delivery: Methods and Protocols. Totowa, NJ: Humana Press; 2013. p. 77–86.10.1007/978-1-62703-110-3_723104335 · doi ↗ · pubmed ↗
- 3Praitis V, Casey E, Collar D, Austin J. Creation of low-copy integrated transgenic lines in Caenorhabditis elegans. Genetics. 2001;157(3):1217–26. doi: 10.1093/genetics/157.3.1217 11238406 PMC 1461581 · doi ↗ · pubmed ↗
- 4Radman I, Greiss S, Chin JW. Efficient and rapid C. elegans transgenesis by bombardment and hygromycin B selection. P Lo S One. 2013;8(10):e 76019. doi: 10.1371/journal.pone.0076019 24130756 PMC 3794024 · doi ↗ · pubmed ↗
- 5Khodakova AS, Vilchis DV, Blackburn D, Amanor F, Samuel BS. Population scale nucleic acid delivery to Caenorhabditis elegans via electroporation. G 3 (Bethesda). 2021;11(7):jkab 123. doi: 10.1093/g 3journal/jkab 123 33872353 PMC 8495937 · doi ↗ · pubmed ↗
- 6Thekke-Veetil T, Mc Coppin NK, Domier LL, Hajimorad MR, Lambert KN, Lim H-S, et al. Transient expression of a luciferase m RNA in plant-parasitic and free-living nematodes by electroporation. Mol Biochem Parasitol. 2022;250:111489. doi: 10.1016/j.molbiopara.2022.111489 35640846 · doi ↗ · pubmed ↗
- 7Stevenson Z, Moerdyk-Schauwecker M, Banse S, Patel D, Lu H, Phillips P. High-throughput library transgenesis in Caenorhabditis elegans via transgenic arrays resulting in diversity of integrated sequences (TARDIS). e Life. 2023;12:RP 84831.10.7554/e Life.84831 PMC 1032850337401921 · doi ↗ · pubmed ↗
- 8Sarov M, Murray J, Schanze K, Pozniakovski A, Niu W, Angermann K, et al. A genome-scale resource for in vivo tag-based protein function exploration in C. elegans. Nat Methods. 2014.10.1016/j.cell.2012.08.001PMC 397930122901814 · doi ↗ · pubmed ↗