The rs1403543 Polymorphism of AGTR2, Which Encodes the Type-2 Angiotensin II Receptor, and Left Ventricular Mass in Polish Full-Term Newborns
Iwona Gorący, Karol Miler, Klaudyna Lewandowska, Monika Rychel, Beata Łoniewska, Andrzej Ciechanowicz

TL;DR
This study explores how a genetic variation in the AGTR2 gene affects heart mass in female Polish newborns.
Contribution
The study identifies a novel association between AGTR2:rs1403543 polymorphism and left ventricular mass in female newborns.
Findings
Female newborns with AGTR2 GA or AA genotypes had lower LVM/body mass ratios than GG homozygotes.
Higher frequency of AGTR2 A allele genotypes was observed in lower tertiles of LVM/body metrics in females.
Abstract
Background/Objectives: Left ventricular hypertrophy is a significant independent risk factor for increased cardiovascular morbidity and mortality. There are some reports indicating an association of rs1403543 (1675G>A) polymorphism in the AGTR2 gene, which encodes the type-2 angiotensin II receptor, with left ventricular hypertrophy or increased left ventricular mass (LVM) in adults. The aim of this study was to analyze the possible association of the AGTR2:rs1403543 polymorphism with LVM in full-term Polish healthy newborns. Methods: The study group comprised 207 consecutive, full-term, healthy newborns. LVM was assessed, on the 3rd day after birth, from the M-mode echocardiographic measurements of left ventricular dimensions using the Penn convention, with the Huwez et al.-modified equation mode. The AGTR2 polymorphism was identified by PCR-RFLP in genomic DNA extracted from cord…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
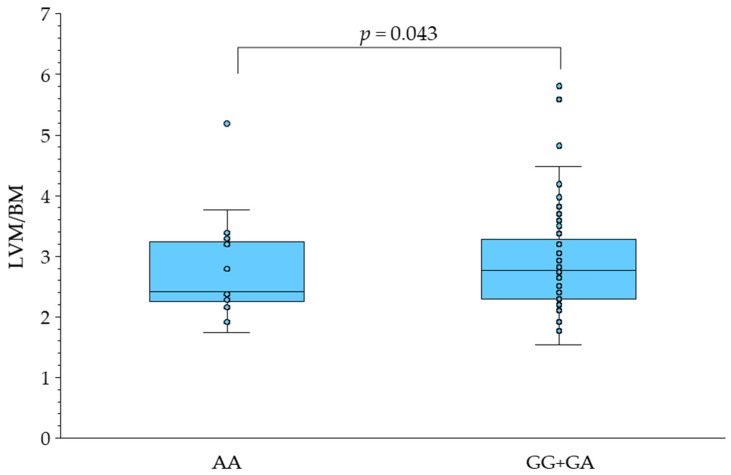 Figure 1
Figure 1Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsHormonal Regulation and Hypertension · Renin-Angiotensin System Studies · Birth, Development, and Health
1. Introduction
Left ventricular hypertrophy (LVH) is recognized when the left ventricular mass index (LVMI), calculated as the left ventricular mass (LVM) divided per body surface area (BSA), is >95 g/m^2^ in women or >115 g/m^2^ in men [1,2]. A number of studies have demonstrated that LVH is a significant independent risk factor for increased cardiovascular morbidity and mortality and, therefore, a major public health burden, especially in light of an aging population [3]. The results of several studies have suggested that approximately 30% to 70% of cardiac mass variability can be explained by heredity [4,5,6,7,8].
The renin–angiotensin system (RAS) is both a major regulator of maternal–fetal health [9] as well as the crucial endocrine element in the regulation of renal and cardiovascular homeostasis [10]. One crucial element of this homeostasis is the action of angiotensin II (ANGII) as a ligand via two types of receptors. Stimulation of the type-1 angiotensin II receptor (AT1R) results, i.a., in an increase of blood pressure and in induction of cardiac hypertrophy and fibrosis, while activation of the type-2 receptor (AT2R) gives opposing effects [10].
The expression of AT2R is high in the neonatal heart and declines after birth (for review, see [11]). It is worth noting that the heart masses of 7-day-old and 14-day-old AT2R-knockout mice were significantly higher than those of age-matched control mice. However, the heart-to-body-mass ratio in AT2R-deficient mice in this early postnatal period was significantly lower than those of age-matched control mice [12]. In addition, the AT2R deficiency resulted in a stronger increase in blood pressure in response to infusion of angiotensin II [13,14,15]. This suggests that the stimulation of AT2R by angiotensin II affects postnatal cardiac growth, possibly via reducing body mass gain and lowering blood pressure [10,12].
The type-2 angiotensin II receptor in humans is encoded by the AGTR2 gene located at chromosome Xq23. The common AGTR2:rs1403543 polymorphism gives a transition of guanine (G) to adenine (A) 1332 base pairs (bp) upstream of the translation start codon (-1332G>A), also known as 1675G>A (e.g., G-to-A transition located 1675 bp downstream of the transcription start) [16]. The dbSNP variant details give AGTR2:rs1403543 as being located at NC_000023.11:g.116170939G>A according to genome assembly GRCh38.p14 chr X and as having two transcripts with locations NM_000686.5:c.-95-29G>A and NM_001385624.1:c.-36+123G>A. In this article, we will refer to this single nucleotide variant as rs1403543 or 1675G>A. The A allele has a global ALFA allele frequency of 52%, and the reference G allele has 48% (a T allele being extremely rare; dbSNP) [17].
So far, only a few reports have focused on the analysis of the association between AGTR2:rs1403543 polymorphism and LVMI and/or LVH and, in addition, these reports have yielded contradictory results [18,19,20,21,22,23,24,25]. In 2001, Schmieder et al. showed, in a group of 120 young males of European descent, that hypertensive (but not normotensive) subjects hemizygous for the AGTR2:1675A allele had a greater LVMI as compared with hypertensives with the reference AGTR2 variant (with the 1675G allele) [18]. One year later, in the GLAOLD (Glasgow Heart Scan Old) study of subjects aged 55–74 years, Herrmann et al. found that the frequency of AGTR2:1675A allele carriers was significantly higher in males with LVH than in those without left ventricular hypertrophy, and this effect was not observed in females [19]. In contrast, Alfakih et al. reported a significantly higher prevalence of GG homozygous females and G hemizygous males among a group with magnetic resonance imaging (MRI)-assessed LVH as compared to a group without LVH (heterozygous females were excluded from the analysis) [20]. Also, in EPOGH men (but not in women), LVMI increased significantly with higher sodium excretion (4.2 g/m^2^ per 100 mmol) in carriers of the reference (1675G) allele, with the opposite tendency in males with the AGTR2:1675A allele [21]. In 2007, Ott et al. reported a lower LVM in males with the 1675G variant who were particularly susceptible to LVM modification by increased salt intake [22]. In contrast, Orlowska-Baranowska et al. found no significant association of AGTR2:1675G>A polymorphism with LVH in Polish patients with aortic stenosis [23]. Huber et al. also revealed no significant association of this AGTR2 variant with LVMI in treated patients with arterial hypertension in Germany [24]. However, in patients with hypertrophic cardiomyopathy, Carstens et al. reported an allele-dependent modulating effect of the AGTR2:rs1403543 polymorphism on LVH (with a decrease in wall thickness by ~0.5 mm with each 1675A allele) which was independent of such covariates as causal mutation of hypertrophic cardiomyopathy or blood pressure [25]. It has not escaped our notice that all the above-mentioned studies concerning the association of the AGTR2:rs1403543 polymorphism and LVM have been carried out in adults. However, it is worthy to underline that reliable evidence suggests that cardiovascular disease (CVD) begins early in life [26,27]. There have been reports of association of CVD risk factors in childhood with both increased cardiac growth in children [28,29] and LVH in adults [30,31]. Therefore, the aim of our study was to analyze the possible association of the AGTR2:rs1403543 polymorphism with echocardiographically measured left ventricular mass in full-term Polish healthy newborns.
2. Materials and Methods
2.1. Newborns
The criteria for newborn eligibility in the present study as well as newborn cord blood (500 μL) collection for the isolation of genomic DNA have been described before [32,33]. In brief, the study group comprised 207 healthy Polish newborns (96 females and 111 males), born between 38 and 40 weeks (inclusive) of gestation to healthy women with uncomplicated pregnancies. The sex of the newborn, body mass (BM; kg) and body length (BL; m) were taken from standard hospital records. Body surface area (BSA; m^2^) was calculated using the Mosteller formula [34]:
All the studied newborns, on the third day after delivery, underwent transthoracic echocardiography as described previously [6,35]. Measurement techniques followed American Society of Echocardiography conventions. Left ventricular masses were estimated from echocardiographic left ventricular dimensions using the Penn convention, with the equation modified by Huwez et al. [36]:
where IVST, LVPWT and LVID denote interventricular septal thickness, left ventricular posterior wall thickness and left ventricular internal dimension, respectively. To give standardized parameters, the left ventricular mass was divided by body mass (LVM/BM, g/kg), body length (LVM/BL, g/m) or body surface area (LVM/BSA, g/m^2^). The study was conducted in accordance with the latest Declaration of Helsinki (2024) and was approved by the bioethics committee at the Pomeranian Medical University in Szczecin, Poland. Parental informed consent for newborn control data was obtained for each newborn.
2.2. AGTR2:rs1403543 Genotyping
Genomic DNA was isolated from cord blood as described previously [32,33]. For the genotyping of the AGTR2:rs1403543 polymorphism, a PCR-RFLP (polymerase chain reaction–restriction fragment length polymorphism) method was applied with 5′-GGAAAGTAGAACATACATTAAATG-3′ as the forward primer, and with 5′-CCTGTAAGAGAA ACAGCAGCTAAAGAATT′-3′ as the reverse primer (both primers from TIB MOL BIOL, Poznań, Poland). The AGTR2 amplicons were subsequently digested with the EcoRI restriction enzyme (Thermo Fisher Scientific, Waltham, MA, USA). The PCR product was cut into fragments of 95 base pairs (bp) and 25 bp if the 1675G reference allele was present or remained uncut (with an amplicon of 120 bp in length) with the 1675A allele. Both electrophoretic separation of the restriction products and the verification of PCR-RFLP results were carried out as described before [33].
2.3. Statistical Analyses
Normality of the quantitative data was assessed by Kolmogorov–Smirnov or Lilliefors tests. As most quantitative variables were not normally distributed, all are presented as medians with minimum and maximum values. Quantitative data were compared using Mann–Whitney tests or Kruskal–Wallis tests, if necessary. Categorical data were assessed using chi-squared tests. Statistical significance was defined as p < 0.05. All data were analyzed using a data analysis software system (Statistica, version 13, TIBCO software, Palo Alto, CA, USA).
3. Results
There were 28 (29.2%) GG homozygotes, 45 (46.9%) GA heterozygotes and 23 (23.9%) AA homozygotes among female newborns, and the frequency of the AGTR2:1675A allele in this group was 47.4%. The AGTR2:rs1403543 genotype distribution in female newborns was consistent with the Hardy–Weinberg equilibrium (p = 0.557). There were 58 (52.2%) male newborns hemizygous for the AGTR2:1675G reference allele and 53 (47.8%) male newborns with the 1675A allele.
Clinical and echocardiographic newborn characteristics are shown in Table 1. There were no significant differences between females and males in terms of body length, left ventricular dimensions (IVST, LVPWT and LVID) or LVM and LVM indices (LVM/BM, LVM/BL and LVM/BSA). Only body mass (BM) and body surface area (BSA) in male newborns were significantly higher than in females.
There were no significant differences in the values of clinical and echocardiographic variables in female newborns in regard to AGTR2:rs1403543 genotype except for LVM/BM. The LVM/BM in female newborns carrying at least one 1675A allele (with a GA or AA genotype) was significantly lower as compared to its value in females homozygous for the reference AGTR2 allele (Table 2 and Figure 1).
Other than for BSA, there were no significant differences in clinical and echocardiographic characteristics of male newborns in regard to AGTR2:rs1403543 hemizygosity. The BSA in male newborns hemizygous for the AGTR2:1675A allele was significantly lower as compared to males with the reference variant (with the 1675G allele) (Table 3). Further, separately for females and males, the distributions of AGTR2:rs1403543 polymorphism in tertiles of LVM indices were analyzed. No significant associations were found both in females or males between the distributions of AGTR2:1675G>A variants or tertiles of LVM indices except for LVM/BL and LVM/BSA in female newborns (Table 4 or Table 5, respectively). The frequency of AGTR2 genotypes with at least one AGTR2:1675A allele was significantly higher for both LVM/BM and LVM/BSA in female newborns from the lower tertile as compared with subjects from the upper tertile (Table 4).
Finally, after exclusion of GA heterozygous female newborns (due to the presumed effect of random inactivation of the one of the X chromosomes in females), we assessed the differences between GG/G and AA/A subjects (GG homozygous females + G hemizygous males versus AA homozygous females + A hemizygous males).
After the exclusion of GA heterozygotes, no significant differences between female newborns and male newborns were found in the values of body length, left ventricular dimensions or LVM and LVM indices. Only the body mass (BM) and body surface area (BSA) in male newborns were significantly higher as compared to females (Table 6).
There were no significant differences in the values of clinical and echocardiographic variables between subjects carrying the 1675G allele and carriers of the 1675A variant (Table 7).
No significant differences in the frequency distributions of the AGTR2 polymorphism with regard to the tertiles of LVM indices were found in the comparison of GG/G with AA/A newborns (Table 8).
4. Discussion
This study has focused on the analysis of the association between the AGTR2:rs1403543 polymorphism and left ventricular mass (LVM) in full-term healthy newborns. The results revealed no significant association of AGTR2 variants with LVM indices in male newborns, but they were associated with a lower LVM in female newborns carrying the 1675A allele. Our results with female newborns are consistent with those of a previous study by Alfakih et al. [20], who reported both a lower LVM index in AA/A subjects (i.e., in AA females and A males) than in GG/GA females and an excess of GG/G subjects among a group of patients with MRI-determined left ventricular hypertrophy. In addition, in a family cohort of patients with hypertrophic cardiomyopathy (HCM), Carstens et al. found a decrease in average wall thickness of ~0.5 mm with each AGTR2:1675A allele independent of the effects of the primary HCM causal mutation, blood pressure and other hypertrophy covariates [24]. However, it is also worthy to note that other studies either did not confirm in women an association of the 1675A AGTR2 allele with LVM [21] or LVH [19,23,25] or revealed such association in males only [18,19,22]. In contrast, Kuznetsova et al. found that LVMI increase (related to higher salt intake) was significantly higher in males with the 1675G variant as compared to males with the A allele [21]. Therefore, taking into consideration the above discrepancies, we decided to carry out our study in healthy newborns born at term. The reasons for this are that newborns have a relative lack of exposure (or at least no long-term exposure) to confounding environmental factors, coexisting diseases or medication, and full-term healthy newborns are a more appropriate target group to study the association between genetic polymorphisms and cardiovascular intermediate phenotypes such as cardiac mass [37]. Last, but not least, an argument in favor of performing an association analysis of X-chromosome-located genes (such as AGTR2) in female newborns is the low frequency of nonrandom (skewed) X-chromosome inactivation (XCI) in female newborns [38] (although, even so, in one analysis, heterozygotes were excluded). In addition, for the rs1403543 biallelic polymorphism of the AGTR2 gene located at chromosome X, women have three possible genotypes (GG, GA or AA), while men are hemizygous either for 1675G or the 1675A allele. Therefore, to ensure equal gene dosage, we carried out an association analysis of AGTR2 polymorphism with LVM indices in the combined group (consisting of female and male newborns) after exclusion of heterozygous female newborns as in the study by Alfakih et al. [20].
The AGTR2 gene spans approximately 5 kb and consists of three exons. Only the third exon encompasses the complete protein-coding sequence of 363 amino acids of the G-protein-coupled receptor [39]. The AGTR2 gene 5′-flanking region contains typical eukaryotic promoter sequence motifs but also an Interferon Consensus Sequence Binding Protein (ICSBP) site and a putative Embryonal, Long Terminal Repeat Binding Protein (ELP) site what suggests a possible unusual regulation of AGTR2 expression [39]. In addition, Warnecke et al. have indicated that sequence elements in intron 1 of the AGTR2 gene are necessary for its efficient transcription [40]. Kuznetsova et al. have suggested that the AGTR2:rs1403543 polymorphism located in intron 1 is probably functional [21]. However, the molecular effects of AGTR2:1675G>A transition have been not fully elucidated and remain unclear. In 1999, Nishimura et al. pointed out that the AGTR2:rs1403543 transition is located within the lariat branchpoint motif of intron 1, which disrupts the efficiency of mRNA splicing. The authors also reported that cDNA from code containing the A allele consists of all three exons but cDNA from code with the G allele is 60 nucleotides shorter due to a lack of exon 2. In addition, the amount of mRNA transcribed from the G allele code was significantly lower as compared with mRNA from the A allele code [41]. However, six years later Warnecke et al. indicated that the AGTR2:1675G>A polymorphism modulates receptor protein expression rather than mRNA splicing. Using in vitro transfection assays with a luciferase reporter gene, the authors revealed that subjects carrying the 1675G allele may express higher levels of the type-2 angiotensin II receptor protein than those with the A allele [42]. It is not possible to rule out that the intronic rs1403543 polymorphism affects transcription of this gene per se but, additionally, it is in close linkage disequilibrium with other AGTR2 variants of real functional importance, as discussed below.
Our analyses (using https://www.ensembl.org/Homo_sapiens/Tools/LD, accessed on 3 April 2025) in European-descent populations from the 1000 Genomes (1KG) Project have indicated that rs14003543 is in tight (r^2^ 0.785, D′ 0.957), very tight (r^2^ 0.842, D′ 0.999) or even in complete (r^2^ 1.000, D′ 1.000) linkage disequilibrium (LD) with the AGTR2:rs11091046 polymorphism (GRCh38.p14 chr X:NC_000023.11:g.116173873A>C (dbSNP); also known as 3123C>A or 4599C>A [37] for TSI (Toscani in Italy), CEU (Utah Residents with Northern and Western European ancestry) or GBR (British in England and Scotland), respectively). The rs11091046 polymorphism is located in the 3′-untranslated region of AGTR2 exon 3, at a microRNA-response-element position possibly corresponding to the hsa-miR-208a-5p and hsa-miR-208b-5p binding sites [43]. The miR-208a and miR-208b elements are encoded in introns of the myosin heavy chain 6 gene (MYH6) or the myosin heavy chain 7 gene (MYH7), respectively [44,45]. Van Rooij et al. showed that miR-208a is required, i.a., for cardiomyocyte hypertrophy and fibrosis [44], while miR-208b is rather involved in the control of muscle myosin content [45]. Therefore, Yvert et al. suggested that AGTR2 expression may be influenced partly by miR-208 a- and/or b-5p and that the C allele of the AGTR2:rs11091046 polymorphism (in linkage disequilibrium with the 1675A allele) can reduce the binding affinity of miRNA to mRNA, leading to an increased amount of type-2 angiotensin II receptor protein [43]. It is also worthy to note that the AGTR2:rs11091046 polymorphism has been previously found to be associated with several cardiovascular phenotypes such as acute myocardial infarction [46], hypertension [47] or hypertrophic cardiomyopathy [48]. In the latter study, carried out with 103 genetically independent subjects with HCM, Deinum et al. showed a significant decrease in LVMI with the number of AGTR2:3123C alleles in women only but no association of this index with rs11091046 in men [48].
A major limitation of our study is its relatively low statistical power mainly due to a relatively small sample size (96 female newborns and 111 male newborns). Such limited numbers of subjects in our study were due to the fact that the recruitment of newborns to the study had been completed several years ago, and good-quality genomic DNA samples remained only in these 207 newborns for which we also had the results of ultrasound measurements of heart dimensions. To further depict the issue, we have computed (using the Open Epi tool, version 3.01, a free and open-source software available at www.openepi.com, accessed on 12 February 2025) the minimum sample size of male newborns for 80% statistical power and 5% type I error rate with the assumptions that the ratio of G to A hemizygotes is equal to 1.09 (58/53) and LVM/BL is lower than 15.5 g/m in 29.3% (17 of 58) of G subjects and in 37.7% (20 of 53) of A subjects (Table 5). Under the above assumptions, the minimum sample size necessary varied from 992 to 1039 male newborns (517–542 G subjects and 475–497 A subjects). On the other hand, the sample size in our study (n = 211) was even greater than in four of eight previously published reports focusing on association analyses of AGTR2:rs14003543 or AGTR2:rs11091046 with LVM or LVH [18,19,20,21,22,23,24,25]. It is also worth noting that the number of female newborns in our study (n = 96) was greater than in the study by Alfakih et al. (n = 72) or by Deinum et al. (n = 40), who previously revealed the association of lower LVM in women with the 1675A allele [20] or with the 3123C variant [48], respectively.
5. Conclusions
The results of our study suggest that the AGTR2:rs1403543 polymorphism may be associated with physiological variability in cardiac mass in female newborns. However, we are fully aware that the inclusion of AGTR2 polymorphism analysis into a cardiovascular risk assessment will require prior prospective longitudinal studies in much larger groups of newborns of specific ethnic origin, taking into account both the period and the degree of exposure to cardiovascular risk factors.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Bula K. Ćmiel A. Sejud M. Sobczyk K. Ryszkiewicz S. Szydło K. Wita M. Mizia-Stec K. Electrocardiographic criteria for left ventricular hypertrophy in aortic valve stenosis: Correlation with echocardiographic parameters Ann. Noninvasive Electrocardiol.201924 e 1264510.1111/anec.1264530896064 PMC 6931645 · doi ↗ · pubmed ↗
- 2Lang R.M. Bierig M. Devereux R.B. Flachskampf F.A. Foster E. Pellikka P.A. Picard M.H. Roman M.J. Seward J. Shanewise J.S. Recommendations for chamber quantification: A report from the American Society of Echocardiography’s Guidelines and Standards Committee and the Chamber Quantification Writing Group, developed in conjunction with the European Association of Echocardiography, a branch of the European Society of Cardiology J. Am. Soc. Echocardiogr.2005181440146310.1016/j.echo.2005.10.00516376782 · doi ↗ · pubmed ↗
- 3Bornstein A.B. Rao S.S. Marwaha K. Left Ventricular Hypertrophy Stat Pearls [Internet]Stat Pearls Publishing Treasure Island, FL, USA 2025 Available online: https://www.ncbi.nlm.nih.gov/books/NBK 557534/(accessed on 12 February 2025)32491466 · pubmed ↗
- 4Swan L. Birnie D.H. Padmanabhan S. Inglis G. Connell J.M. Hillis W.S. The genetic determination of left ventricular mass in healthy adults Eur. Heart J.20032457758210.1016/S 0195-668X(02)00524-912643891 · doi ↗ · pubmed ↗
- 5Sharma P. Middelberg R.P. Andrew T. Johnson M.R. Christley H. Brown M.J. Heritability of left ventricular mass in a large cohort of twins J. Hypertens.20062432132410.1097/01.hjh.0000202815.18083.0316508579 · doi ↗ · pubmed ↗
- 6Bella J.N. Mac Cluer J.W. Roman M.J. Almasy L. North K.E. Best L.G. Lee E.T. Fabsitz R.R. Howard B.V. Devereux R.B. Heritability of left ventricular dimensions and mass in American Indians: The Strong Heart Study J. Hypertens.20042228128610.1097/00004872-200402000-0001115076185 · doi ↗ · pubmed ↗
- 7Assimes T.L. Narasimhan B. Seto T.B. Yoon S. Curb J.D. Olshen R.A. Quertermous T. Heritability of left ventricular mass in Japanese families living in Hawaii: The SAPPHI Re Study J. Hypertens.20072598599210.1097/HJH.0b 013e 32809 bd 74017414662 · doi ↗ · pubmed ↗
- 8Verhaaren H.A. Schieken R.M. Mosteller M. Hewitt J.K. Eaves L.J. Nance W.E. Bivariate genetic analysis of left ventricular mass and weight in pubertal twins (the Medical College of Virginia twin study)Am. J. Cardiol.19916866166810.1016/0002-9149(91)90361-N 1877484 · doi ↗ · pubmed ↗