Proteomic Analysis of Bifidobacterium animalis AR668 and AR668-R1 Under Aerobic Culture
Yaping Liu, Xiaoxiao Zhao, Miao Yang, Xin Song, Guangqiang Wang, Yongjun Xia, Liang Zhao, Zhiqiang Xiong, Lianzhong Ai

TL;DR
This study identifies proteins involved in oxygen tolerance in a modified strain of Bifidobacterium animalis, offering insights for improving probiotic applications.
Contribution
The study reveals specific proteins and pathways that enable oxygen tolerance in an evolved B. animalis strain.
Findings
AR668-R1 has 212 upregulated and 390 downregulated proteins compared to AR668 under aerobic conditions.
Key pathways for oxygen tolerance include translation, transmembrane transport, and carbohydrate metabolism.
Overexpression of Mth2 improved AR668-R1 growth by 19.8% under aerobic conditions.
Abstract
Bifidobacterium animalis is a widely used probiotic with significant health benefits, but its application is limited by oxygen sensitivity. Our laboratory previously developed an oxygen-tolerant B. animalis AR668-R1 using adaptive laboratory evolution under aerobic culture, but the molecular mechanism remains unclear. In this work, compared to the wild-type parental strain B. animalis AR668, 212 upregulated and 390 downregulated proteins were identified in AR668-R1 under aerobic conditions through comparative proteomic analysis. Enrichment analysis of the differentially expressed proteins between AR668 and AR668-R1 identified the potential oxygen-tolerant related pathways, including the translation process, transmembrane transport system, and carbohydrate metabolism. Furthermore, five potential oxygen-tolerance proteins (DapE, Mth2, MutT, Eno, and MsrAB) were validated by RT-qPCR that…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
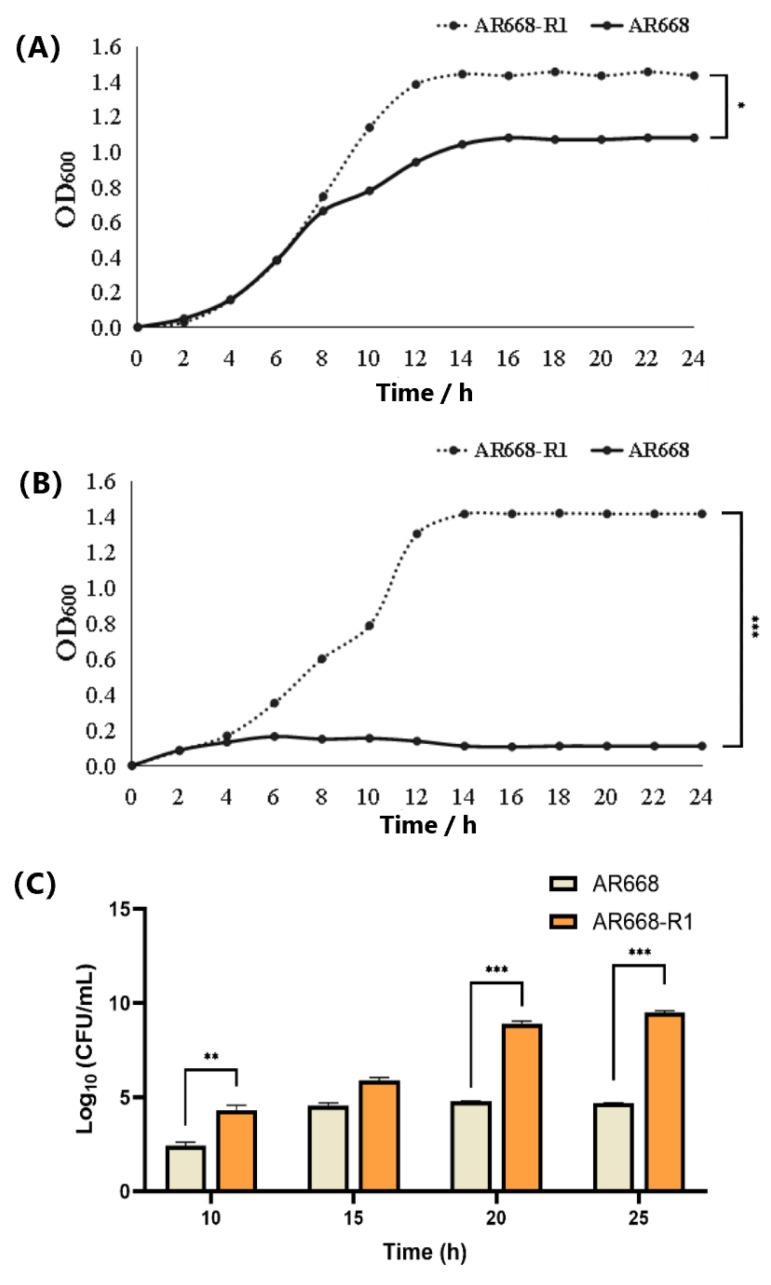 Figure 1
Figure 1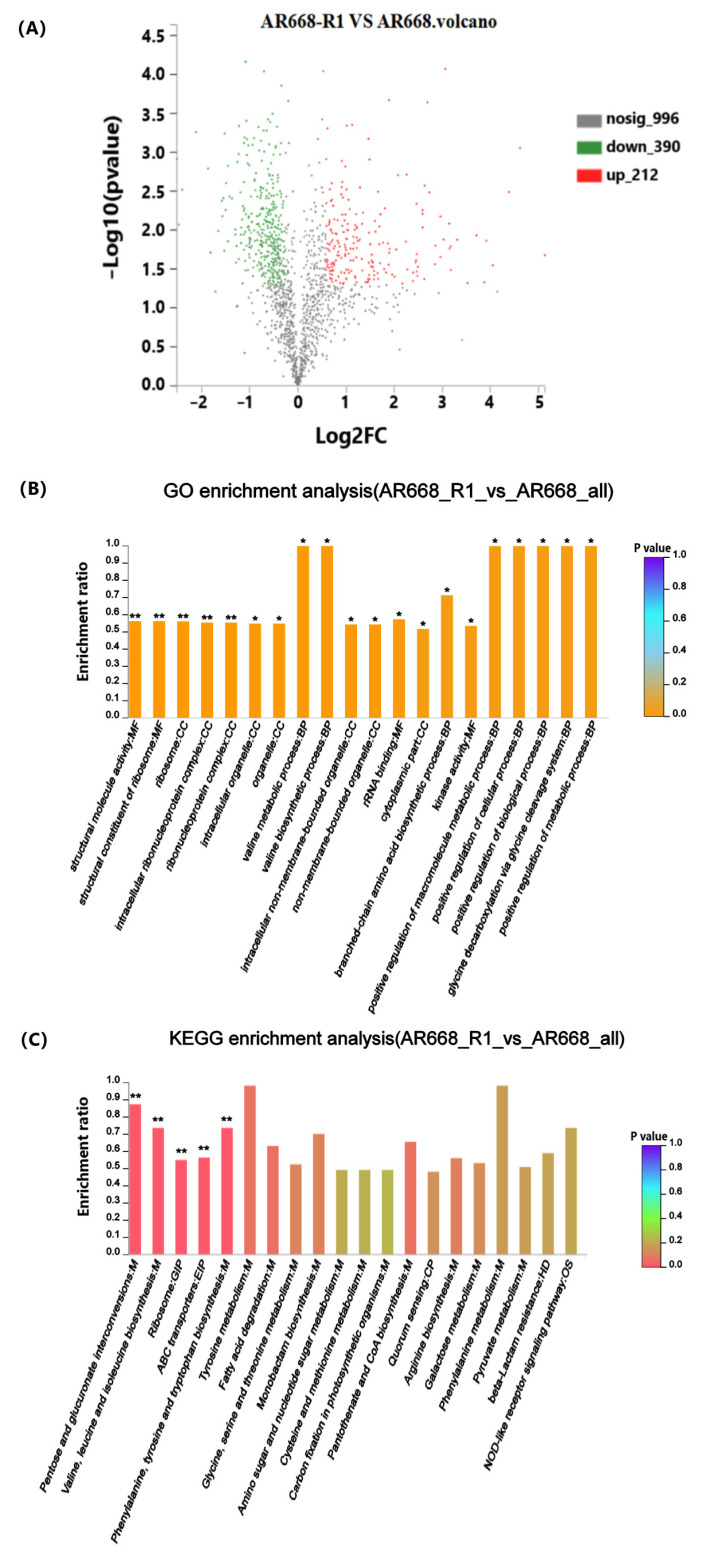 Figure 2
Figure 2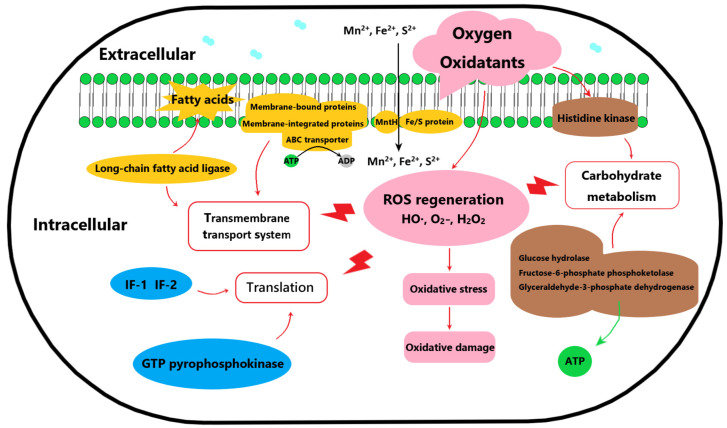 Figure 3
Figure 3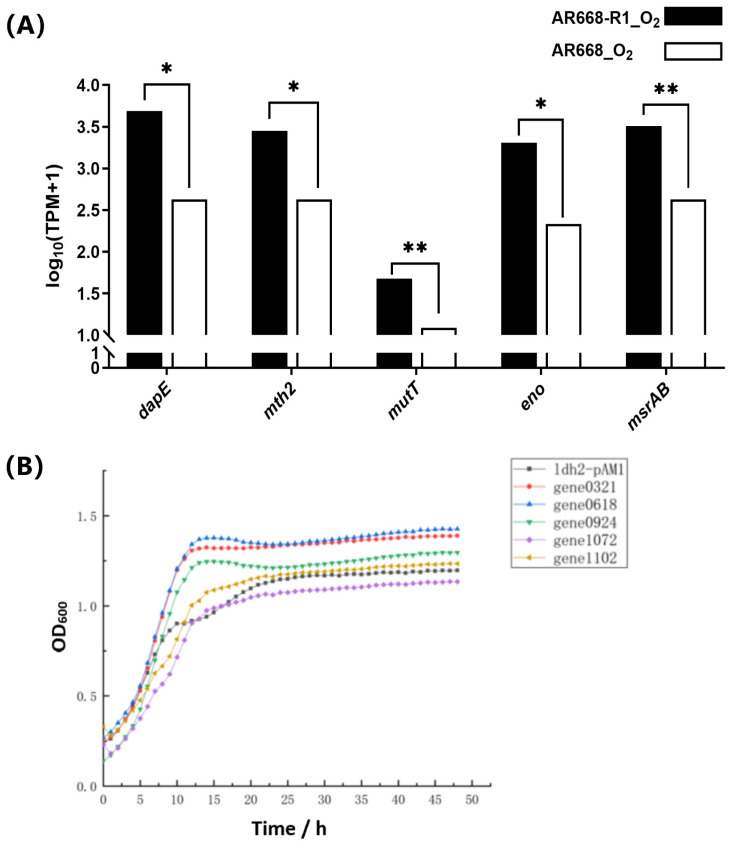 Figure 4
Figure 4- —Key Special Project of Synthetic Biology in Shanghai “Science and Technology Innovation Action Plan”
- —National Science Fund for Distinguished Young Scholars
- —Natural Science Foundation of Shanghai
- —Shanghai Education Committee Scientific Research Innovation Project
- —Yunnan Key Laboratory of Application Technology for Specialty Forest Fruits
- —Key Technology Research and Demonstration Application for Improving Quality and Efficiency of Tamarind Industry
- —Shanghai Engineering Research Center of Food Microbiology
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsProbiotics and Fermented Foods · Microbial Metabolic Engineering and Bioproduction · Genomics and Phylogenetic Studies
1. Introduction
Bifidobacterium animalis, a Gram-positive, anaerobic bacterium, is widely recognized for beneficial effects on human health, particularly the regulation of the gut microbiota. This probiotic strain is characterized by its ability to ferment carbohydrates producing lactic acid and acetic acid, which contributes to the maintenance of a healthy intestinal environment [1]. The physiological properties of B. animalis include the ability to adhere to intestinal epithelial cells, modulate immune responses, and produce antimicrobial compounds that inhibit the growth of pathogenic bacteria [2]. These characteristics make B. animalis a valuable component of functional foods and dietary supplements aimed at promoting gut health. In the food industry, B. animalis has been extensively utilized in the production of fermented dairy products such as yogurt, cheese, and kefir [3], which enhance food nutritional value and sensory properties, such as texture and flavor [4]. Additionally, B. animalis has been incorporated into non-dairy products, including infant formula, cereals, and beverages, to provide probiotic benefits to a wider range of consumers [5]. Despite the widespread application, a significant limitation of B. animalis is sensitivity to oxygen, which restricts viability and functionality in aerobic environments. This oxygen sensitivity poses challenges for the storage, distribution, and incorporation of B. animalis into non-fermented foods and supplements, where exposure to oxygen is inevitable.
The oxidative stress response in model organisms such as E. coli is well characterized, depending primarily on the OxyR/SoxRS regulatory systems and classical reactive oxygen species (ROS)-scavenging enzymes including catalase and superoxide dismutase [6]. However, Bifidobacterium species are notably lacking in these conserved oxidative defense mechanisms [7,8]. Recent studies revealed that B. breve UCC2003 employs an alternative stress response strategy mediated by a coordinated regulatory network. This network harbors ClgR (stress-responsive transcriptional regulator), LexA (the DNA damage response regulator), and HrcA and HspR (heat shock protein regulators) [9]. For hydrogen peroxide detoxification, B. bifidum primarily relies on alkyl hydroperoxide reductase (Ahp) and the thioredoxin reductase-AhpC (TrxR-AhpC) antioxidant system [10]. Furthermore, oxygen tolerance is facilitated by NADH oxidase (NOX) and HemN-like oxygen reductase activity [11]. Notably, B. longum LTBL16 could enhance oxidative resistance through the upregulation of peroxidases and the activation of the NOX-dependent FoxO signaling pathway [12]. These findings collectively demonstrated that Bifidobacterium species develop diverse and specialized mechanisms to mitigate oxidative stress.
However, despite these intrinsic defense systems, most bifidobacteria remain highly sensitive to oxygen exposure, which severely limits their viability during industrial production and food applications where aerobic conditions are unavoidable. To address this limitation, adaptive laboratory evolution (ALE) strategies for selecting oxygen-tolerant strains have been extensively employed to engineer starter cultures with enhanced robustness for industrial food fermentation processes [13]. Wild-type parental strain B. animalis AR668 was isolated from infant feces [14]. The oxygen-tolerant strain B. animalis AR668-R1 was obtained after the ALE adaptation of AR668 [15] (Figure S1). We previously employed genomic and phenotypic analyses to elucidate the potential oxygen tolerance mechanism in B. animalis AR668-R1, revealing that membrane proteins and ABC transporter are involved in conferring oxygen tolerance of B. animalis. However, there is still a need for a comprehensive understanding of the molecular changes that occur during oxygen adaptation [16]. Here, this study aimed to identify potential oxygen-tolerant proteins in B. animalis AR668-R1 using proteomic analysis, which provides valuable insights into the oxygen-tolerant mechanism by which bifidobacteria cope with oxidative stress at the protein level.
2. Materials and Methods
2.1. Strain, Media, and Culture Condition
The bacterial strains and plasmids are listed in Table 1. B. animalis AR668 and AR668-R1 were cultivated in a BS medium (Catalog #HB0394-1, Hope Bio, Qingdao, China) at 37 °C under anaerobic conditions without shaking or aerobic conditions with shaking at 100 rpm.
2.2. Hydrogen Peroxide Tolerance and Cell Growth of AR668 and AR668-R1
The overnight culture of AR668 or AR668-R1 (OD_600_ = 1.0–1.2) was adjusted to achieve an OD_600_ of 0.60 ± 0.05. The bacterial suspension was then added to sterile BS broth containing hydrogen peroxide (H_2_O_2_, Catalog #H1009, Sigma-Aldrich, St. Louis, MO, USA) at concentrations ranging from 3.5 mM to 10 mM, followed by incubation for 3 h. The control group consists of BS broth without H_2_O_2_. After appropriate dilution, the survival rate of AR668 or AR668-R1 under H_2_O_2_ stress was calculated using the colony counting method on BS agar plates. Each group was repeated three times.
An overnight culture of AR668 or AR668-R1 was inoculated at 5.0% into 100 mL of the BS medium under aerobic conditions for 24 h. The absorbance at 600 nm was measured every 2 h, with three replicates each time. The colony-forming units per milliliter (CFU/mL) of AR668 or AR668-R1 was obtained by plating 10 μL stationary phase cultures on the BS medium at 37 °C for 24 h under anaerobic conditions by 10-fold serial dilutions.
2.3. Protein Sample Preparation, Sequencing, and Enrichment Analysis
The strains AR668-R1 and AR668 were grown in the BS liquid medium with agitation at 100 rpm until reaching the early logarithmic phase. The cells were harvested by centrifugation (4 °C, 4000× g, 10 min), flash-frozen in liquid nitrogen for 10 min, and lysed in a protein lysis buffer (Catalog #B14001, Bimake, TX, USA) containing 8 M urea, 1.0% SDS, and protease inhibitors. Homogenization was performed using a high-throughput tissue grinder (3 cycles, 40 s each), followed by incubation on ice for 30 min with brief vortexing every 5 min. After centrifugation (16,000× g, 4 °C, 30 min), the supernatant was collected. Protein integrity was verified by an SDS-PAGE analysis, followed by concentration quantification using a BCA Protein Assay Kit (Catalog #23227, Thermo Fisher Scientific, Waltham, MA, USA). For digestion, 100 μg of protein was reduced with 10 mM Tris (2-carboxyethyl)phosphine (TCEP, Catalog #20490, Thermo Fisher Scientific, MA, USA) at 37 °C for 60 min, alkylated with 40 mM iodoacetamide (IAM, Catalog #I6125, Sigma-Aldrich, St. Louis, MO, USA) in the dark for 40 min, and digested overnight at 37 °C using trypsin (Catalog #V5111, Promega, Madison, WI, USA) at a 1:50 enzyme–protein ratio. After protein digestion, peptides were desalted using HLB (Hydrophilic-Lipophilic Balanced) cartridges, dried, and reconstituted in 0.1% trifluoroacetic acid (TFA, Catalog #PI85183, Thermo Fisher Scientific, MA, USA).
The peptides were resuspended in a spectrometry loading buffer (2.0% ACN, 0.1% formic acid) with iRT peptides for retention time calibration. An LC-MS/MS analysis was conducted on an EASY nLC-1200 system coupled to a Q Exactive HF-X mass spectrometer (Thermo Fisher Scientific, MA, USA) at Majorbio Bio-Pharm Technology Co., Ltd. (Shanghai, China). The data-independent acquisition (DIA) mode was employed, alternating between full-scan MS and MS/MS. The C18 reversed-phase column (75 μm × 25 cm, Thermo, USA) was equilibrated with solvent A (2% acetonitrile with 0.1% formic acid) and solvent B (80% acetonitrile with 0.1% formic acid) at a flow rate of 300 nL/min. The peptides were eluted using the following gradient: 0–70 min, 5–23% B; 70–90 min, 23–29% B; 90–100 min, 29–38% B; 100–102 min, 38–48% B; 102–103 min, 48–100% B; and 103–120 min, 100% B (maintained).
The raw data were processed using Spectronaut™ (v14) with iRT-based retention time adjustment, in which the database search software automatically performs internal normalization during differential protein quantification. The quantification relied on 6 peptides per protein and 3 daughter ions per peptide, excluding shared/modified peptides. Proteomic data were analyzed on the Majorbio Cloud platform. Differential protein expression was determined using an R-based t-test (with thresholds set at fold change > 1.20 or <0.83, p < 0.05). Functional annotation and enrichment of differentially expressed proteins (DEPs) were performed via GO (http://geneontology.org/, accessed on 10 March 2025) and KEGG (http://www.genome.jp/kegg/, accessed on 10 March 2025) databases.
2.4. RNA Preparations and Gene Expression Analysis by Reverse Transcription Quantitative Polymerase Chain Reaction (RT-qPCR)
Total RNA was isolated using a total RNA extraction reagent (Catalog #B511311, Sangon Biotech, Shanghai, China). Due to the thick cell wall of B. animalis, lysozyme was used for 1 h pretreatment to extract high-quality RNA [18]. The integrity and quantity of RNA were determined by agarose gel electrophoresis and NanoDrop 2000 spectrophotometry (Thermo Fisher Scientific, MA, USA), respectively. Complementary DNA (cDNA) was prepared by the PrimeScriptRT reagent kit (Catalog #RR037A, Takara Bio Inc., Shiga, Japan) and used for RT-qPCR. cDNA synthesis was performed as follows: 1 pg^−1^ μg RNA was mixed with 4 μL 4× g DNA wiper Mix in a total volume of 16 μL RNase-free dH_2_O (42 °C, 2 min), followed by the addition of 4 μL 5× HiScript III qRT SuperMix (37 °C, 15 min) and enzyme inactivation (85 °C, 5 s). cDNA concentration was measured by Nanodrop 2000 at A260. RT-qPCR was performed using SYBR Premix Ex Taq (Catalog #DRR420A, Takara Bio Inc., Shiga, Japan), with each reaction containing 10 μL of 2 × SYBR Green Mix, 0.5 μL of cDNA, 0.5 μL of each primer (20 μmol/L), and 8.5 μL of ddH_2_O, gently mixed in a PCR tube. The PCR reactions were carried out as follows: initial denaturation at 95 °C for 30 s, followed by 40 cycles of denaturation at 95 °C for 5 s and annealing/extension at 60 °C for 30 s, with a final extension at 72 °C for 5 min. The gene-specific qPCR primers are listed in the Supplementary Information (Table S2). The reference gene rplD verified in previous experiments was selected as a control [7]. The qPCR reactions were performed in 96-well (LightCycle 8-tube strips), and three repetitions were made for each sample.
2.5. Plasmid Construction
The gene sequences and primers are listed in the Supplementary Information (Tables S1 and S2). The DNA fragments of gene0321, gene0618, gene0924, gene1072, and gene1102 were amplified from the B. animalis genome by PCR using primers and inserted into the E. coli-B. animalis shuttle plasmid pAM1-ldh2 with digestion by HindIII and XbaI using the ClonExpress one-step cloning kit (Catalog #C113, Vazyme Biotech Co. Ltd., Nanjing, China). The overexpression plasmids were validated by PCR (Figure S2). All the plasmids were constructed in E. coli Top10, cultured in Luria–Bertani (LB) broth supplemented with 100 μg/mL ampicillin (Amp) at 37 °C overnight with shaking (200 rpm). The amplified plasmids were electroporated into B. animalis, and cultured in BS medium supplemented with 5 μg/mL erythromycin (Em) at 37 °C under anaerobic conditions without shaking. Empty plasmid ldh2-pAM1 was used as a negative control.
2.6. Statistical Analysis
The results were statistically analyzed by one-way analysis of variance using the statistical software package SPSS 26.0. The statistical degree of significance was set at * p < 0.05, ** p < 0.01, and *** p < 0.001.
3. Results and Discussion
3.1. Hydrogen Peroxide Tolerance and Cell Growth of AR668-R1 and AR668
The maximum lethal concentration of H_2_O_2_ for AR668 and AR668-R1 was determined using the plate counting method (Table S3). It was found that AR668 completely died at 6.0 mM H_2_O_2_, while AR668-R1 retained viability (9.7% survival) even at 8.5 mM H_2_O_2_. Under anaerobic conditions, both AR668 and AR668-R1 grew normally (Figure 1A). AR668-R1 reached a maximum OD_600_ of 1.46, which was significantly higher than that of AR668 (1.08, p < 0.05). The anaerobic growth improvement observed in AR668-R1 may result in ALE-driven metabolic reorganization, which has been demonstrated to provide oxidative stress tolerance in S. cerevisiae [19]. Under aerobic conditions, the maximum growth OD_600_ of AR668-R1 reached 1.44, corresponding to 3.09 × 10^9^ CFU/mL (Figure 1B,C). This indicates that AR668-R1 could grow normally in the aerobic environment. In contrast, the growth of AR668 was severely inhibited under aerobic conditions. Therefore, the oxygen tolerance of AR668-R1 showed a significant improvement through ALE adaptation compared to AR668.
3.2. Functional Enrichment Significance Analysis
To understand the oxygen tolerance mechanism of AR668-R1, the protein expression levels of AR668-R1 and AR668 under aerobic conditions were determined by proteomic analysis, followed by functional annotation and pathway analysis using GO and KEGG. A total of 1598 proteins were detected in AR668 and AR668-R1, with 212 upregulated proteins and 390 downregulated proteins in AR668-R1 using AR668 as the control (Figure 2A). The top five significant (p < 0.01) GO functional enrichment results were all related to ribosomes, which function to synthesize polypeptide chains from amino acids according to mRNA instructions (Figure 2B). It indicated that the cultivation of B. animalis under an aerobic environment primarily affects the translation process, including protein synthesis and amino acid metabolism. In addition, the GO analysis revealed significant enrichment (p < 0.05) in valine metabolism and synthesis, rRNA binding, branched-chain amino acid synthesis, kinase activity, glycine cleavage, and the positive regulation of metabolic processes, which were potentially associated with oxygen tolerance in B. animalis. Similarly, the KEGG pathway analysis showed significant (p < 0.01) enriched pathways including pentose and glucuronate interconversions; valine, leucine, and isoleucine biosynthesis; ribosomes; and ABC transport systems (Figure 2C). It suggested that in addition to protein synthesis, the pathways related to carbohydrate metabolism and transmembrane transport systems are also important to oxygen tolerance.
3.3. Analysis of Differential Expressed Proteins
Protein expression with significant differences (p < 0.01) were analyzed in the proteomic data (Table 2). Compared to AR668, 37 upregulated (FC > 2.0) and 38 downregulated (FC < 0.5) proteins were identified in AR668-R1. Based on the function of the differential proteins, a summary of the involved pathways associated with oxygen tolerance was found in AR668-R1 (Figure 3). For example, the ribosome serves as the exclusive site for protein biosynthesis, and proteins associated with ribosome modification and translation, including small subunit ribosomal RNA methyltransferase G (24.48-fold), 30 S ribosomal protein S6 (8.34-fold), and 50 S ribosomal proteins L24 and L33 (3.23-fold and 3.17-fold, respectively), were significantly upregulated in AR668-R1. Translation initiation factors IF-1 and IF-2 were also upregulated, suggesting the enhanced translation efficiency in AR668-R1. During the translation process, the elongation factor (EF) promotes peptide chain elongation, while the RelE factor cleaves messenger RNA at specific ribosomal sites to terminate translation. Therefore, the upregulation of RelE inhibits global protein synthesis, which leads to persistence and enhances aerobic survival capacity [20]. Notably, the GTP pyrophosphokinase protein enhances RelE activity, thereby significantly reducing translation efficiency [21]. Collectively, EF, IF−1, and IF-2 were upregulated 2.31- to 5.52-fold, while the GTP pyrophosphate kinase protein was downregulated by 4.34-fold. These differential expressions demonstrated faster translation speed and higher precision in protein synthesis under aerobic culture, which were directly associated with oxygen tolerance in B. animalis.
The cell membrane fatty acid composition is critical for stress resistance in Bifidobacterium [22]. In AR668-R1, long-chain fatty acid ligase and membrane-bound proteins were obviously downregulated, while ABC transporter proteins, implicated in substrate transport and oxygen tolerance [23], were upregulated (5.52-fold, 4.77-fold, and 3.03-fold for putative transporter protein, conserved membrane protein, and ABC transporter permease, respectively). Additionally, AR668-R1 exhibited enhanced carbohydrate metabolism, with a 2.8-fold upregulation of glyceraldehyde-3-phosphate dehydrogenase, a key enzyme in glycolysis [24], leading to elevated NADH/NADPH generation and ATP production under oxygen stress [25]. The two-component system in AR668-R1 was also more active, evidenced by a 2.44-fold upregulation of histidine kinase, enabling rapid environmental signal response and stress adaptation. As revealed by the proteomic analysis, the potential oxygen tolerance mechanism of AR668-R1 involves enhanced translation efficiency, improved transmembrane transport for membrane repair, and accelerated carbohydrate metabolism, which constitute intrinsic defense systems against oxidative stress (Figure 3). The coordinated extracellular protection and intracellular metabolic and translational adaptations enable AR668-R1 to maintain cellular integrity and function under aerobic conditions.
3.4. Validation of Potential Oxygen-Tolerance Proteins of B. animalis
According to the above proteomic analysis of differentially expressed proteins and our previous genomic analysis [15], we excluded certain proteins based on their functional annotation and literature evidence in metabolic pathways, including ribosome biogenesis-related proteins, non-oxygen-tolerant related proteins (e.g., cold-shock protein), and uncharacterized proteins. Notably, the upregulation of ribosomal proteins and translation factors in AR668-R1 indicated an enhanced translational capacity, which may contribute to oxygen tolerance through the rapid synthesis of stress-response proteins during oxidative challenge. This observation was supported by a previous report on E. coli, in which oxidative stress-resistant mutants increased the expression of translation machinery components to maintain proteostasis under ROS exposure [26]. The remaining candidates were evaluated through their functional contributions to essential oxygen-tolerance mechanisms, including membrane integrity maintenance demonstrated by membrane-bound transporters, oxidative DNA damage repair involving 8-oxo-dGTPase, energy metabolism through glycolytic enzymes, and redox homeostasis maintained by methionine sulfoxide reductases (Table 3). We further selected the following five proteins for experimental validation to confirm their roles that may be involved in the oxygen tolerance of AR668-R1. DapE (membrane protein) and Mth2 (7,8-dihydro-8-oxoguanine-triphosphatase) were selected based on the function in maintaining membrane stability under oxidative stress, as demonstrated in studies of oxygen stress responses in lactic acid bacteria [27]. MutT (DNA mismatch repair protein) was prioritized for its function as a crucial regulator in mitigating DNA damage during oxidative stress, reducing stress-induced oxidative damage by enhancing gene expression levels [11]. Eno (enolase) was included due to its regulatory function in diverse stress responses [28], while MsrAB (peptide methionine sulfoxide reductase) was chosen as a key component of the ModRS-regulated oxidative stress defense system [29].
The gene expression of these five potential oxygen-tolerance proteins was analyzed using RT-qPCR in B. animalis AR668 and AR668-R1 under aerobic conditions (Figure 4A). The expression levels of gene0321 (dapE), gene0618 (mth2), gene0924 (mutT), gene1072 (eno), and gene1102 (msrAB) were significantly (p < 0.05) upregulated, which indirectly indicated the correctness of our proteome data. To further validate these five potential oxygen-tolerant proteins, we overexpressed their corresponding genes in B. animalis AR668-R1 using the ldh2 promoter. The RT-qPCR analysis showed that all the target mRNAs were upregulated, with transcript levels increasing >3-fold compared to the control (Figure S3). Under aerobic conditions, compared to the control strain, the overexpression of gene0321, gene0618, and gene0924 resulted in better growth (Figure 4B). Specifically, the OD_600_ of the strain overexpressing gene0321 and gene0618 were 17.6% and 19.8% higher than the control during the stationary phase, respectively. It confirmed that the DapE and Mth2 proteins are associated with the oxygen tolerance of B. animalis, providing valuable insights into the oxygen-tolerant mechanisms of B. animalis at the protein level. By analyzing the oxygen tolerance mechanism of the aerotolerant B. animalis AR668-R1 strain, this study provides a theoretical foundation for developing oxygen-resistant Bifidobacterium. Additionally, the inherent oxygen sensitivity of B. animalis has previously limited its applications in food products. Its wider application in fermented foods will be made possible by clarifying its oxygen tolerance mechanism. Furthermore, this insight can serve as a valuable reference for the genetic engineering of other oxygen-resistant strains, expanding the use of probiotics in a broader range of applications.
4. Conclusions
Bifidobacterium animalis AR668-R1, developed by ALE, exhibited high oxygen tolerance in comparison to the parental strain AR668. Comparative proteomic analysis identified 212 upregulated and 390 downregulated proteins in AR668-R1, revealing that the potential oxygen tolerance mechanism involves extracellular membrane components and intracellular transport systems to mitigate oxidative damage. Five potential oxygen-tolerance proteins (DapE, Mth2, MutT, Eno, and MsrAB) were validated, with Mth2 overexpression significantly improving growth by 19.8%. These findings provide valuable insights into the molecular basis of oxygen tolerance in B. animalis.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Evdokimova S.A. Karetkin B.A. Guseva E.V. Gordienko M.G. Khabibulina N.V. Panfilov V.I. Menshutina N.V. Gradova N.B. A study and modeling of Bifidobacterium and Bacillus coculture continuous fermentation under distal intestine simulated conditions Microorganisms 20221092910.3390/microorganisms 1005092935630373 PMC 9147766 · doi ↗ · pubmed ↗
- 2Ji J. Lin W. Liu S.J. Jiao Z. Li X. Probiotics, prebiotics, and postbiotics in health and disease Med Comm 20234 e 42010.1002/mco 2.42037929014 PMC 10625129 · doi ↗ · pubmed ↗
- 3Sun Y. Guo S. Kwok L.Y. Sun Z. Wang J. Zhang H. Probiotic Bifidobacterium animalis ssp. lactis Probio-M 8 improves the fermentation and probiotic properties of fermented milk J. Dairy Sci.20241076643665710.3168/jds.2024-2486338825144 · doi ↗ · pubmed ↗
- 4Zhang Y. Hou Y. Zhang S. Jing N. Zhang H. Xie Y. Liu H. Yan J. Ren J. Jin J. Bifidobacterium animalis A 12, a probiotic strain that promotes glucose and lipid metabolism, improved the texture and aroma of the fermented sausage Foods 20231233610.3390/foods 1202033636673428 PMC 9858581 · doi ↗ · pubmed ↗
- 5Pimentel T.C. de Assis B.B.T. dos Santos Rocha C. Marcolino V.A. Rosset M. Magnani M. Prebiotics in non-dairy products: Technological and physiological functionality, challenges, and perspectives Food Biosci.20224610158510.1016/j.fbio.2022.101585 · doi ↗
- 6Baussier C. Oriol C. Durand S. Py B. Mandin P. Small RNA Oxy S induces resistance to aminoglycosides during oxidative stress by controlling Fe–S cluster biogenesis in Escherichia coli Proc. Natl. Acad. Sci. USA 2024121 e 231785812110.1073/pnas.231785812139495911 PMC 11572966 · doi ↗ · pubmed ↗
- 7Cronin M. Zomer A. Fitzgerald G. van Sinderen D. Identification of iron-regulated genes of Bifidobacterium breve UCC 2003 as a basis for controlled gene expression Bioengineered 2012315916910.4161/bbug.18985 PMC 337093422179149 · doi ↗ · pubmed ↗
- 8Schöpping M. Vesth T. Jensen K. Franzén C.J. Zeidan A.A. Genome-wide assessment of stress-associated genes in bifidobacteria Appl. Environ. Microbiol.202288 e 02251-2110.1128/aem.02251-2135311508 PMC 9004370 · doi ↗ · pubmed ↗