The TIP60-CD44 Axis Modulates Colorectal Cancer Stemness
Asad Mohammad, Sudhakar Jha

TL;DR
This study shows that TIP60 influences colorectal cancer stemness by regulating CD44, impacting cancer cell growth and spread.
Contribution
The study identifies the TIP60-CD44 axis as a novel regulator of colorectal cancer stemness and metastasis.
Findings
TIP60 depletion reduces colorectal cancer cell proliferation and alters colony formation.
Loss of TIP60 decreases CD44 expression, a key cancer stem cell marker.
CD44 overexpression reverses the effects of TIP60 depletion on cell proliferation and EMT.
Abstract
HIV-1 Tat-interactive protein of 60 kDa (TIP60) is a lysine acetyltransferase protein that can acetylate histone and non-histone proteins. This study highlights TIP60’s role in regulating colorectal cancer (CRC) stemness. The depletion of TIP60 resulted in a marked decrease in cellular proliferation, highlighting TIP60’s involvement in the progression of CRC. Additionally, the loss of TIP60 impacted colony formation, transitioning from densely packed structures to dispersed spindle networks along with the loss of E-cadherin, indicating its role in the epithelial–mesenchymal transition (EMT). Three-dimensional culture models suggest that TIP60 is vital for spheroid formation, highlighting its importance in maintaining cancer stem cell properties in CRC. TIP60-depleted cells showed increased invasion in a 3D basement membrane extract (BME) invasion matrix, demonstrating its essential role…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
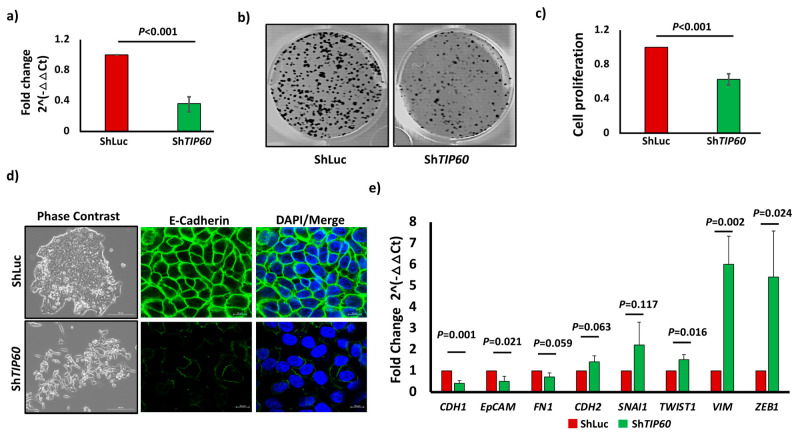 Figure 1
Figure 1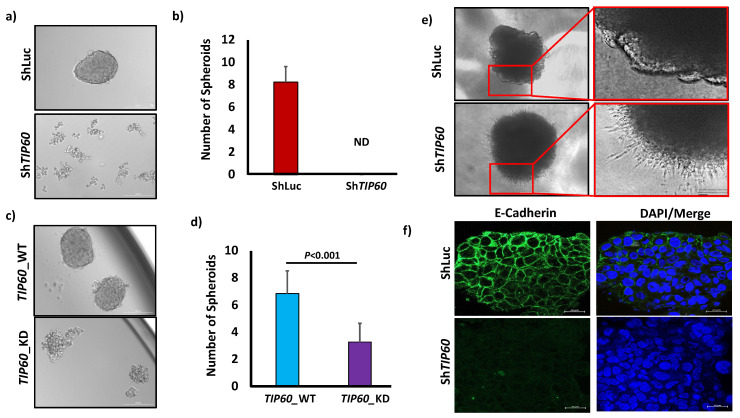 Figure 2
Figure 2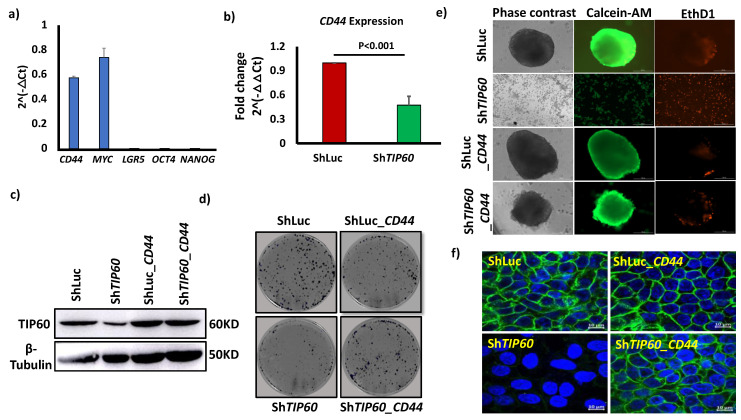 Figure 3
Figure 3- —Oklahoma State University College of Veterinary Medicine, Department of Physiological Sciences
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsImmunotherapy and Immune Responses · Epigenetics and DNA Methylation · Peptidase Inhibition and Analysis
1. Introduction
Colorectal cancer (CRC) is the second leading cause of cancer-related mortality worldwide [1]. Advancements in therapeutic strategies have significantly improved the survival rates of patients diagnosed with CRC [2]. Despite the development of innovative targeted medications and several therapeutic combinations, tumor recurrence and drug resistance remain significant obstacles to conventional, targeted, and immunotherapeutic therapies [3]. Colorectal cancer stem cells (CRCSCs) are the primary determinant of cancer development, recurrence, and dissemination [4]. Cancer stem cells (CSCs) are a distinct group of cells in the tumor bulk that can renew themselves and differentiate into other types of cells [5]. CSCs have a role in tumor recurrence, metastasis, heterogeneity, and multidrug resistance [6,7].
HIV-1 Tat-interactive protein (TIP60), also known as lysine acetyltransferase 5 (KAT5), was first discovered as a cellular acetyltransferase protein interacting with HIV-1 Tat [8]. TIP60 belongs to the MYST (Moz, Ybf2/Sas3, Sas2, and TIP60) family of acetyltransferases and has been shown to acetylate histones [9] and non-histone proteins [10,11]. The MYST family members regulate chromatin remodeling, gene-specific transcription, and DNA damage repair [12]. TIP60 downregulation has been associated with several cancer types, such as colorectal, gastric, and breast cancer [13]. TIP60 has been shown to regulate cellular migration and invasion [14] and inhibit epithelial–mesenchymal transition (EMT) [15]. Apart from cancer, TIP60 has also been shown to be involved in regulating stem cell functions. In hematopoietic stem cells (HSCs), TIP60 is essential for its maintenance [16]. The TIP60 histone acetyltransferase in embryonic stem cells (ESCs) triggers the activation of genes necessary for cell division and suppresses genes that induce differentiation [17]. CD44 belongs to the cell adhesion molecule (CAM) class and is a single-chain transmembrane glycoprotein. CD44 is implicated in various types of cancers, such as breast cancer, prostate cancer, colorectal cancer, and pancreatic cancer. CD44 is involved in tumor growth, EMT, cellular invasion, metastasis, and resistance to chemotherapy [18]. It is an established marker for CSCs [19].
There remains a lack of knowledge regarding the role that TIP60 plays in the control of cancer stemness. Our research sheds light on the significance of TIP60 in the stemness of CRC, demonstrating that TIP60 affects the cancer stemness characteristics in CRC. Mechanistically, we identified that TIP60 controls cancer stemness via CD44.
2. Materials and Methods
2.1. Cell Culture
Colorectal cancer cell lines HCT116 (Cat. No. CCL-247) and 293T (Cat. No. CRL-3216) were purchased from ATCC (Manassas, VA, USA)and were grown in DMEM High glucose (Gibco, Thermo Fisher Scientific, Waltham, MA, USA; Cat. No. 11-995-081) supplemented with 10% Fetal Bovine Serum (Corning, NY, USA; Cat. No. 35-010-CV).
2.2. Generation of Stable Cell Line
An shRNA designed against the TIP60 (TOP 5′-CTGATCGAGTTCAGCTATGAACTCGAGTTCATAGCTGAACTCGATCAG-3′, Bottom 5′-CAAAAACTGATCGAGTTCAGCTATGAACTCGAGTTCATAGCTGAACTCGATCAG-3′) was cloned into a pLKO vector using AgeI and EcoRI restriction sites. For the amplification of the plasmids, the construct (TIP60-targeting, ShTIP60, and control vector, ShLuc) plasmids were transformed into Stbl2 cells. The lentivirus was generated by transfecting 5 × 10^6^ 293T cells with TIP60-targeting (ShTIP60) and control vector (ShLuc) plasmids using polyethylenimine (Fisher Scientific, Waltham, MA, USA; Cat. No. 24314-2) with a 1:3 ratio and incubated for 24 h. After 24 h, the media were changed, and fresh medium was added and incubated for 48 h. After 48 h, the virus was harvested and used to infect 2 × 10^6^ HCT116 cells with polybrene (Millipore Sigma, Burlington, MA, USA; Cat. No. TR1003) reagent (1 mg/mL). After 24 h, the media containing the virus were replaced with fresh growth media, and puromycin (Fisher Scientific, Waltham, MA, USA; Cat. No. AAJ672368EQ) (5 μg/mL) was added to the growth media for selection. The cells were selected until the mock-transfected cells died.
TIP60 wild-type (TIP60_WT) and TIP60 catalytic dead (TIP60_KD) were generated by retrovirus transfection to 5 × 10^6^ 293T cells with the plasmids (TIP60 wild-type (TIP60_WT) and TIP60 catalytic dead (TIP60_KD)) utilizing Lipofectamine 2000 (Invitrogen, Thermo Fisher Scientific, Waltham, MA, USA; Cat. No. 11668019) following the manufacturer’s instructions. The virus was collected 72 h post-transfection and used to infect 2 × 10^6^ HCT116 cells with polybrene at a 0.4 mg/mL concentration. After 6 h, the virus-containing media were substituted with growth media. Puromycin (5 μg/mL) was added to the growth medium for selection after 24 h. The cells were selected until the mock-transfected cells died, and the puromycin-selected cells were maintained for two weeks to generate stocks.
2.3. CD44 Overexpression
To overexpress CD44 in HCT116 cell lines, a plasmid containing the full-length CD44s (Cat. No.137812) gene was obtained from Addgene (Watertown, MA, USA). HCT116 control (ShLuc) and TIP60-depleted (ShTIP60) cells were placed in 6-well plates. When the cell culture reached 60% confluency, the plasmid (1:3) was introduced into the cells using polyethylenimine. After 72 h post-transfection, the media were substituted with a fresh medium. The cells were selected utilizing neomycin (1 mg/mL) (Research Product International, Mount Prospect, IL, USA; Cat. No. N20040-25).
2.4. RNA Isolation and Quantitative PCR (qPCR)
HCT116 control (ShLuc) and TIP60-depleted (ShTIP60) cells were on a 10 cm dish. After reaching 70% confluency, the cells were trypsinized (Corning, NY, USA; Cat. No. 25-053 CI) and counted utilizing a hemacytometer (Fisher Scientific, Waltham, MA, USA; Cat. No. 22-600-100). One hundred thousand HCT116 control cells (ShLuc) and TIP60-depleted (ShTIP60) cells were seeded into 6-well ultra-low attachment plates (Corning, NY, USA; Cat. No. 07-200-601) and incubated for 7 days. The cells were collected and washed with ice-cold PBS, and the whole RNA was extracted using an RNeasy Kit (Qiagen, Hilden, Germany; Cat. No. 74104), following the manufacturer’s instructions, and reverse-transcribed to cDNA using the iSCRIPT cDNA synthesis kit (Bio-Rad Laboratories, Hercules, CA, USA; Cat. No. 1708891). Quantitative real-time PCR (RT-qPCR) was performed using iTaq Universal SYBR Green Supermix (Bio-Rad Laboratories, Hercules, CA, USA; Cat. No. 1725124) on Quant Studio 6 Pro (Applied Biosystem, Thermo Fisher Scientific, Waltham, MA, USA). The amplification conditions were as follows: 50 °C for 2 min, 95 °C for 5 min; 40 cycles of amplification (95 °C for 15 s, 60 °C for 1 min); and the melt curve step (95 °C for 15 s, 60 °C for 1 min, 60 °C → 95 °C (0.1 °C/s)). The mRNA expression of the gene was normalized to the average of the two housekeeping genes ACTB and GAPDH. Gene expression analysis was performed using qPCR and computed using ΔΔCt. The ΔCt was calculated by subtracting the housekeeping gene’s Ct value from the target gene’s Ct value (ΔCt = Ct_target − Avg Ct_housekeeping, ACTB and GAPDH). To compute ΔΔCt, the values were subtracted from the control sample’s ΔCt from each experimental sample’s ΔCt (ΔΔCt = ΔCt_experimental − ΔCt_control). Finally, gene expression in fold change was calculated using the method 2^−ΔΔCt^ [20]. The primers utilized for detecting mRNA expression levels were purchased from Integrated DNA Technologies (IDT), and the sequences of each forward primer (F) and reverse primer (R) are as Table 1.
2.5. Immunofluorescence
Cells were grown in 6-well plates (Corning Costar, NY, USA; Cat. No. 07-200-83) on 15 mm coverslips. At 70% confluency, the cells were fixed in 100% ice-cold methanol for 15 min. Cells were blocked in 3% BSA (Sigma-Aldrich, St. Louis, MO, USA; Cat. No. A2153) for 1 h and incubated with mouse anti-E-cadherin (BD Biosciences, San Jose, CA, USA; Cat. No. 610182) primary antibody at a 1:200 ratio for 1 h at room temperature. Cells were washed twice with PBS, and the secondary antibody, goat anti-mouse (Invitrogen, Thermo Fisher Scientific, Waltham, MA, USA; Cat. No. A11029), was added at a 1:1000 ratio and incubated for 1 h. After incubation, the cells were washed twice with 1× PBS. The stained cells were then mounted onto the glass slides using DAPI (Vector Laboratories, Newark, CA, USA; Cat. No. NC1848443). Images were captured using LSM 980 with Airyscan 2 confocal laser scanning microscope (Carl Zeiss Microscopy GmbH, Jena, Germany).
2.6. Cell Invasion Assay
Two hundred HCT116 ShTIP60 and ShLuc cells were seeded into a 96-well ultra-low attachment round-bottom plate (Corning, Cat. No. 07-201-680). Immediately after seeding, the cells were centrifuged at 1200 rpm for 5 min and incubated for 72 h for spheroid formation. After 72 h, 100 µL of BME invasion matrix (R&D Systems, Minneapolis, MN, USA; Cat. No. 3500096K) was added to each well. The cells were further incubated for 72-96 h. Images were taken at 10× magnification on a BioTek Cytation 5 imager (Agilent Technologies, Santa Clara, CA, USA).
2.7. Spheroid Formation Assay (3D Culture)
HCT116 control (ShLuc) and TIP60-depleted (ShTIP60) cells were cultured in a 10 cm dish. After reaching 70% confluency, the cells were trypsinized (Corning, Cat. No. 25-053 CI) and counted utilizing a hemacytometer (Fisher Scientific, Waltham, MA, USA; Cat. No. 22-600-100). One hundred cells were seeded in each well of 96-well flat ultra-low attachment plates (Corning, NY, USA; Cat. No. 07-200-603). The cells were incubated for 7 days, and images were taken using a BioTek Cytation 5 imager (Agilent Technologies, Santa Clara, CA, USA). For live and dead cell staining of spheroid culture, Calcein AM (Thermo Fisher Scientific, Waltham, MA, USA; Cat. No. C1430) and Ethidium Homodimer-1 (EthD1) (Thermo Fisher Scientific, Waltham, MA, USA; Cat. No. E1169) were added at a concentration of 1 μg/mL. The cells were incubated for 30 min at 37 °C in a cell culture incubator. After the incubation, images were taken using a BioTek Cytation 5 imager (Agilent Technologies, Santa Clara, CA, USA).
2.8. Colony Formation Assay and Absorbance Assay
HCT116 control (ShLuc) and TIP60-depleted (ShTIP60) cells were grown in a 10 cm dish. The cells were trypsinized and counted using a hemacytometer after 70% confluency. One thousand HCT116 control (ShLuc) and TIP60-depleted (ShTIP60) cells were seeded in 6-well plates and incubated for 11 days. On day 11, cells were fixed with a 0.5% Crystal Violet (Fisher Scientific, Waltham, MA, USA; Cat. No. NC0827019) staining solution. The cells were washed twice with 1× PBS, and 250 µL of Crystal Violet solution was added to each well for 5 min. After incubation, the cells were washed three times with PBS. The plates were air-dried, and images were taken using a BioTek Cytation 5 imager (Agilent Technologies, Santa Clara, CA, USA). For the absorbance assay, 1% SDS (Bio-Rad Laboratories, Hercules, CA, USA; Cat. No. 1610302) was used to destain Crystal Violet. Five hundred microliters of 1% SDS were added to each well of 6-well plates and incubated for 6 h on a shaker at room temperature. After the incubation, 100 μL of the destained solution was added to 96-well plates for HCT116 ShLuc and ShTIP60 cells. Absorbance readings were taken at 590 nm on a BioTek Cytation 5 imager (Agilent Technologies, Santa Clara, CA, USA).
2.9. Western Blot Analysis
Cell lysates were extracted using cold RIPA buffer (Cell Biolabs Inc., San Diego, CA, USA; Cat. No. AKR-190) with protease inhibitor and quantified using Pierce BCA Protein Assay Kit (Thermo Fisher Scientific, Waltham, MA, USA; Cat. No. PI23225). Denatured lysates were subjected to SDS-PAGE, transferred to PVDF membranes, and blocked with 3% BSA (Sigma-Aldrich, St. Louis, MO, USA; Cat. No. A8022) for 1 h. The membranes were then incubated with CD44 primary antibody (Thermo Fisher Scientific, Waltham, MA, USA; Cat. No. MA5-13890) at a dilution of 1:1000 and incubated with the membrane overnight at 4 °C in 1% BSA in TBST. Beta-tubulin primary antibody (Santa Cruz Biotechnology, Dallas, TX, USA; Cat No. sc9104) was incubated overnight at 4 °C with the diluted 1% BSA in TBST buffer. HRP conjugated secondary anti-mouse antibody (Santa Cruz Biotechnology, Dallas, TX, USA; Cat. No. sc516102) against CD44 and anti-rabbit antibody (Santa Cruz Biotechnology, Dallas, TX, USA; Cat. No. sc2357) against β-tubulin were used at a concentration of 1:5000 and incubated at room temperature for 1 h. Pierce ECL (Thermo Fisher Scientific, Waltham, MA, USA; Cat. No. 32106) was used for chemiluminescence. Images were acquired using an Amersham Imager 600 (GE Healthcare Life Sciences, Chicago, IL, USA).
2.10. Statistical Analysis
All the experiments were conducted with three independent biological repetitions. The evaluation of data, calculation of mean values, and standard deviation were performed utilizing Microsoft Excel. Statistical significance (p ≤ 0.05) was assessed through an unpaired, two-tailed Student’s t-test. Error bars indicate the standard deviation derived from three biological replicates.
3. Results and Discussion
3.1. TIP60 Drives Cell Proliferation and Negatively Regulates EMT in CRC
To investigate the functional role of TIP60 in CRC, we utilized shTIP60 to deplete TIP60 expression in CRC cell lines. We achieved a knockdown efficiency of 64% (p = 0.0002) for HCT116 cell lines, as analyzed by qPCR (Figure 1a). To identify the role of TIP60 in CRC proliferation, we performed a colony formation assay (CFA) by seeding 1000 cells in 6-well plates and incubating them for 11 days. The CFA shows a significant reduction in cell proliferation (Figure 1b), which was further confirmed by the absorbance assay showing reduced cell proliferation of 37% (p = 0.0005) in TIP60-depleted (ShTIP60) cells (Figure 1c). This finding is in agreement with previous studies in squamous cell carcinoma [21] and prostate cancer [22], which suggests that TIP60 is necessary for cellular proliferation in CRC. Since TIP60 regulates cellular invasion and metastasis, we looked at the cell morphology to find out if there are any changes between control (ShLuc) and TIP60-depleted (ShTIP60) cells. The phase-contrast image shows that depletion of TIP60 results in a phenotypic change from compact cell colonies to random spindle networks in cultures, suggesting that TIP60-depleted cells undergo EMT. We also tested additional conditions to understand the role of TIP60 in CRC by overexpressing the TIP60 wild-type (TIP60_WT) and mutating its catalytic domain (TIP60_KD), which is essential for its lysine acetyltransferase activity. The phase-contrast image shows a compact colony in HCT116 TIP60_WT cells, whereas in TIP60_KD, it resembles a mesenchymal phenotype, suggesting a dominant negative effect due to mutation in the catalytic domain (Figure S1a). To further confirm TIP60-depleted (ShTIP60) cells undergoing EMT, we performed immunofluorescence staining of E-cadherin and qPCR analysis for EMT genes in HCT116 ShLuc and ShTIP60 cells. E-cadherin staining in ShLuc showed linear junctional staining, whereas in ShTIP60 cells, there was a loss of junctional staining with a punctate pattern, confirming that TIP60-depleted cells undergo EMT (Figure 1d, right panel). qPCR analysis for EMT genes revealed that epithelial markers such as E-cadherin (CDH1) (p = 0.0013) and EpCAM (p = 0.0218) were significantly downregulated. In contrast, mesenchymal markers such as TWIST1 (p = 0.0163), Vimentin (VIM) (p = 0.0028), and ZEB1 (p = 0.0243) were significantly upregulated (Figure 1e). In breast cancer, it has been demonstrated that TIP60 is essential for the epithelial phenotype [15]. The findings of our study indicate that the loss of epithelial phenotype is a consequence of the depletion of TIP60, which suggests that it plays a role as an epithelial gatekeeper in CRC. Together, these data suggest that TIP60 is essential for cell proliferation and epithelial phenotypes in CRC.
3.2. TIP60 Modulates Cancer Stem Cell Characteristics in CRC
To determine TIP60’s functional significance in regulating CRCSCs, we performed a spheroid formation assay by seeding 100 cells per well of ShLuc and ShTIP60 cells in 96-well ultra-low attachment plates. Spheroid formation assay shows that TIP60-depleted cells completely abrogate sphere-forming ability in 3D culture (Figure 2a,b). It has been shown that TIP60 is involved in stem cell function [16,17]. In an in vitro condition, sphere-forming ability is one of the assays for determining the self-renewal and multipotency of CSC subpopulations inside tumors or cancer cell lines, suggesting the importance of TIP60 in regulating CRC stemness. To understand the role of TIP60 in CRC stemness, we also performed a spheroid culture of TIP60_WT and TIP60_KD. Spheroid formation assay in TIP60_WT shows no significant differences compared to ShLuc cells. However, quantification shows that TIP60_WT forms bigger spheroids than ShLuc. In contrast, TIP60_KD shows a reduced number of spheroid (p = 0.0009) formations, smaller in size, indicating the partial loss of TIP60 functions (Figure 2c,d). This suggests that TIP60 plays a crucial role in CRC stemness. The 2D culture in Figure 1d,e shows that TIP60-depleted cells undergo EMT. To find EMT regulation in the 3D culture condition, we seeded 200 cells per well in 96-well round-bottom ultra-low attachment plates, resuspended in a spheroid-formation extracellular matrix (ECM). Since ShTIP60 fails to form spheroids in 3D culture, after seeding 200 cells per well, we spun down the cells at 1500 rpm for 5 min and incubated them for 3 days at 37 °C in sphere-forming conditioned media so the cells could assemble to form aggregates. Our results show that control (ShLuc) cells fail to invade in a 3D invasion matrix, whereas TIP60-depleted (ShTIP60) cells infiltrate in 3D culture (Figure 2e), suggesting TIP60 is involved in maintaining the epithelial phenotype. Moreover, immunofluorescence labeling of E-cadherin in 3D culture (Figure 2f) exhibits a linear junctional staining similar to that shown in 2D culture, confirming the role of TIP60 in cellular migration and invasion. These data suggest that TIP60 regulates CRC stemness and inhibits cellular invasion in CRC.
3.3. The TIP60-CD44 Axis Regulates Cancer Stem Cell Properties
To have a mechanistic insight on how TIP60 regulates CRC stemness and to identify downstream targets for TIP60, we selected genes that are involved in the regulation of pluripotency, self-renewal, and proliferation in embryonic stem cells/adult stem cells and cancer stem cells such as LGR5 [23], MYC [24], OCT4 [25], NANOG [26], and CD44 [19] (Figure 3a). qPCR results shows that CD44 (p = 0.0011) (Figure 3b) and MYC (p = 0.0038) (Figure S1b) were significantly downregulated in TIP60-depleted cells (ShTIP60) in 3D culture, whereas the expression of LGR5, OCT4, and NANOG was undetected. Since CD44 is a marker for CSCs across various solid tumors [19], we were interested in how TIP60 downregulation changes CD44 expression level and whether CD44 acts as a downstream target for TIP60. The role of MYC and TIP60 has already been established, where MYC recruits TIP60 to chromatin [27]. Meanwhile, the TIP60-CD44 axis has not yet been explored. To identify the role of CD44 in CRC, we overexpressed CD44 by transfecting CD44 full-length in HCT116 control (ShLuc) and TIP60-depleted (ShTIP60) cells. Western blot results show that overexpression of CD44 upregulates TIP60 expression levels (Figure 3c). To identify the role of the TIP60-CD44 axis in CRC cell proliferation, we performed a colony formation assay that shows CD44 overexpression restores cell proliferation in HCT116 cells (Figure 3d and S1c). To further confirm the role of CD44 in cancer stemness, we performed a spheroid formation assay. Overexpression of CD44 in TIP60-depleted (ShTIP60) cells restores spheroid formation efficiency (Figure S1d,e), suggesting that the TIP60-CD44 axis maintains the CSC property. We also performed CalceinAM (stains live cells) and EthD1 (stains dead cells) staining. CalceinAM and EthD1 staining show that TIP60-depleted cells (ShTIP60) fail to survive in 3D culture (Figure 3e), suggesting that TIP60-depleted cells undergo anoikis in 3D culture. Furthermore, immunofluorescence staining (Figure 3f) and the phase-contrast image (Figure S1f) also show that TIP60-depleted cells overexpressing CD44 (ShTIP60_CD44) undergo colony compaction, suggesting an EMT reversal phenotype. Taken together, these data suggest that the TIP60-CD44 axis regulates CRC stemness.
Our findings reveal that TIP60 regulates CRC stemness, cell proliferation, epithelial phenotype, and cellular invasion. Our results are consistent with the knockdown of TIP60 in non-small-cell lung cancer, showing reduced cell proliferation, migration, and invasion [28]. It has also been shown that in CRC, TIP60 expression is downregulated (13%) in tumor samples with peritoneal dissemination, distant metastasis, and a higher stage of TNM classification [29]. Similarly, in gastric cancer, where TIP60 is downregulated in 61% of specimens, it correlates with tumor invasion and lymph node metastasis [30]. These data suggest that TIP60 plays a critical role during tumor initiation and progression, but its downregulation may occur during the transition to a metastatic and advanced stage of CRC. In breast cancer, TIP60 acts as a tumor suppressor gene, and its loss induces genomic instability, leading to the development of cancer [31]. These studies highlight that TIP60 is a dynamic epigenetic regulator that may induce tumor-promoting and tumor-suppressing actions. In summary, this study reveals that TIP60 regulates cancer stem cells’ properties, cellular proliferation, and EMT in CRC. Targeting TIP60 will offer a novel therapeutic approach against CRCSCs.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Bray F. Laversanne M. Sung H. Ferlay J. Siegel R.L. Soerjomataram I. Jemal A. Global cancer statistics 2022: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries CA Cancer J. Clin.20247422926310.3322/caac.2183438572751 · doi ↗ · pubmed ↗
- 2Gmeiner W.H. Recent Advances in Therapeutic Strategies to Improve Colorectal Cancer Treatment Cancers 202416102910.3390/cancers 1605102938473386 PMC 10930828 · doi ↗ · pubmed ↗
- 3Hammond W.A. Swaika A. Mody K. Pharmacologic resistance in colorectal cancer: A review Ther. Adv. Med. Oncol.20168578410.1177/175883401561453026753006 PMC 4699262 · doi ↗ · pubmed ↗
- 4Lin C.C. Liao T.T. Yang M.H. Immune Adaptation of Colorectal Cancer Stem Cells and Their Interaction with the Tumor Microenvironment Front. Oncol.20201058854210.3389/fonc.2020.58854233312953 PMC 7708331 · doi ↗ · pubmed ↗
- 5Yu Z. Pestell T.G. Lisanti M.P. Pestell R.G. Cancer stem cells Int. J. Biochem. Cell Biol.2012442144215110.1016/j.biocel.2012.08.02222981632 PMC 3496019 · doi ↗ · pubmed ↗
- 6Phi L.T.H. Sari I.N. Yang Y.G. Lee S.H. Jun N. Kim K.S. Lee Y.K. Kwon H.Y. Cancer Stem Cells (CS Cs) in Drug Resistance and their Therapeutic Implications in Cancer Treatment Stem Cells Int.20182018541692310.1155/2018/541692329681949 PMC 5850899 · doi ↗ · pubmed ↗
- 7Marzagalli M. Fontana F. Raimondi M. Limonta P. Cancer Stem Cells-Key Players in Tumor Relapse Cancers 20211337610.3390/cancers 1303037633498502 PMC 7864187 · doi ↗ · pubmed ↗
- 8Yamamoto T. Horikoshi M. Novel substrate specificity of the histone acetyltransferase activity of HIV-1-Tat interactive protein Tip 60J. Biol. Chem.1997272305953059810.1074/jbc.272.49.305959388189 · doi ↗ · pubmed ↗