Non-Redundant Essential Roles of Proteasomal Ubiquitin Receptors Rpn10 and Rpn13 in Germ Cell Formation and Fertility
Wan-Yu Yue, Yi Zhang, Tian-Xia Jiang, Xiao-Bo Qiu

TL;DR
This study shows that Rpn10 and Rpn13 proteins are crucial for germ cell development and fertility in mice.
Contribution
The study reveals non-redundant roles of Rpn10 and Rpn13 in germ cell formation and fertility.
Findings
Conditional deletion of Rpn13 disrupts proteasome assembly and blocks meiosis in spermatocytes.
Conditional deletion of Rpn10 impairs PGC migration and reduces germ cell numbers.
Loss of either Rpn10 or Rpn13 leads to infertility in both male and female mice.
Abstract
Primordial germ cells (PGCs) undergo proliferation, migration, and sexual differentiation to produce gonocytes, which eventually generate germ cells. The proteasome, which degrades most cellular proteins, is a protein complex with dozens of subunits. The proteasomal ubiquitin receptors Rpn10 and Rpn13 have been shown to play partially overlapping roles in binding ubiquitin chains in vitro and in liver function in vivo. However, the specific role of Rpn10 and Rpn13 in germ cell production remains unclear. We show here that Rpn10 and Rpn13 are each essential for germ cell production and fertility. The conditional deletion of either Rpn10 or Rpn13 in PGCs results in infertility in both male and female mice. Germ cells in testes and ovaries all decreased dramatically in the Rpn13 conditional knockout (cKO) mice. Specifically, the deletion of Rpn13 in PGCs disrupts the assembly of the 26S…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
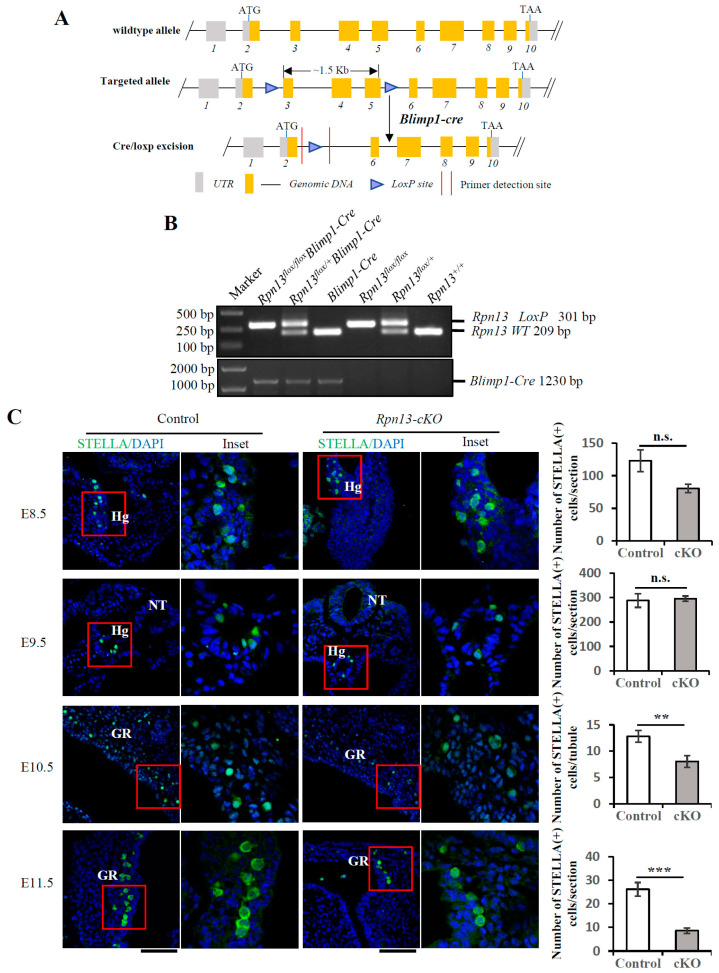 Figure 1
Figure 1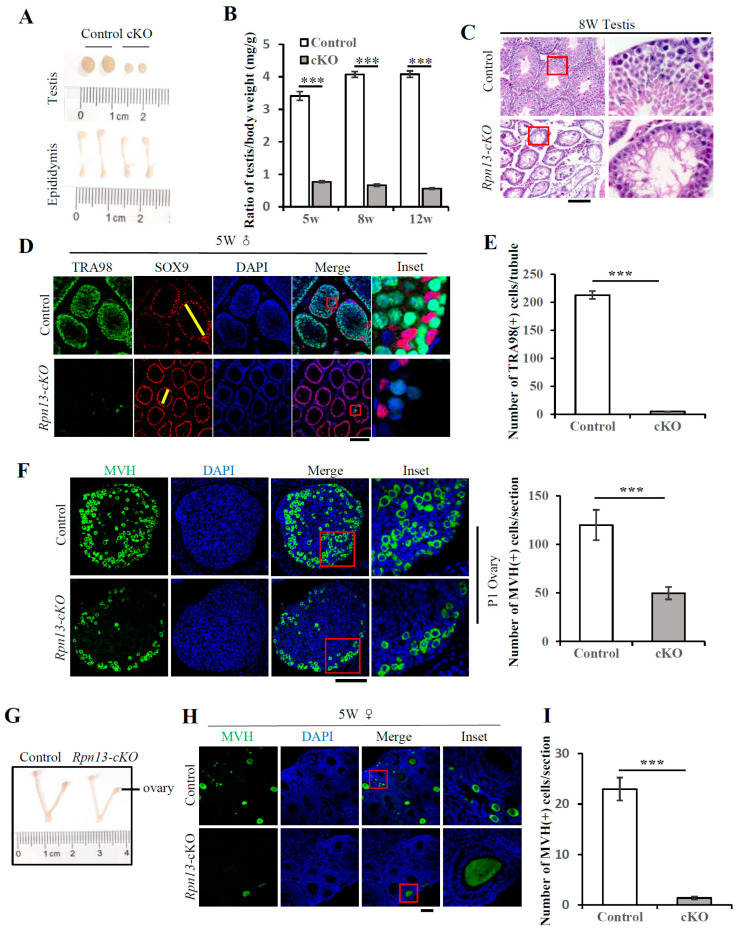 Figure 2
Figure 2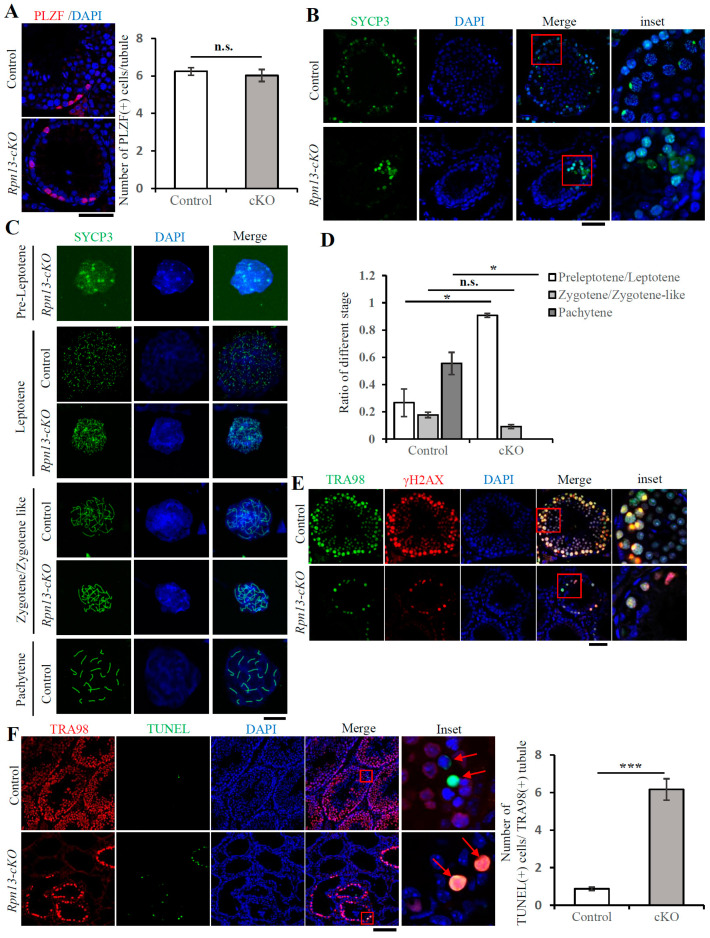 Figure 3
Figure 3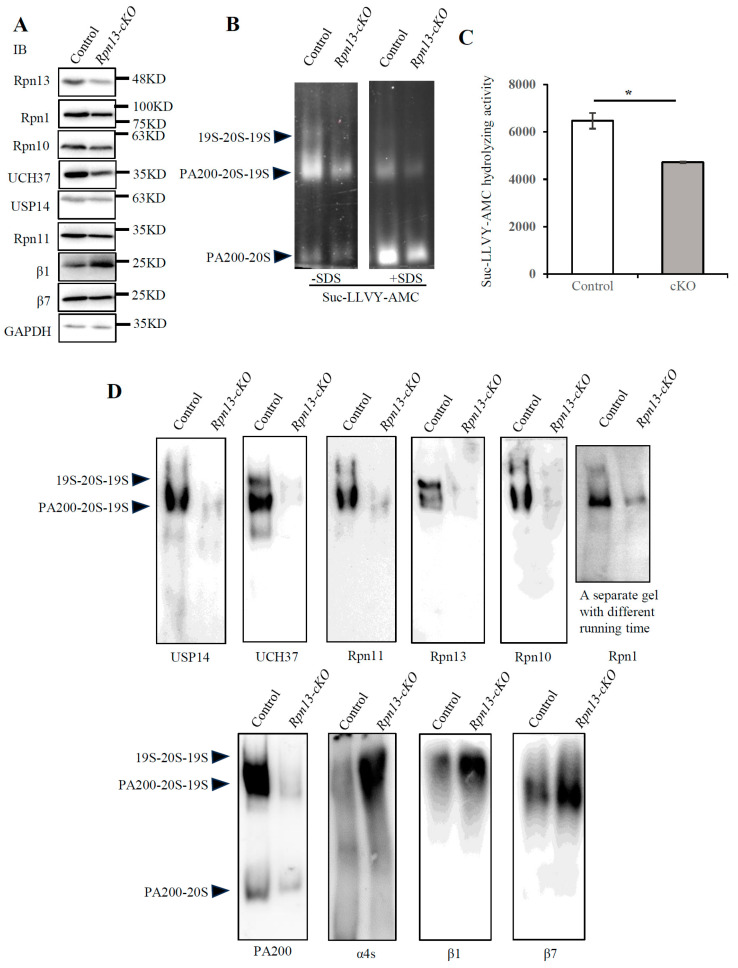 Figure 4
Figure 4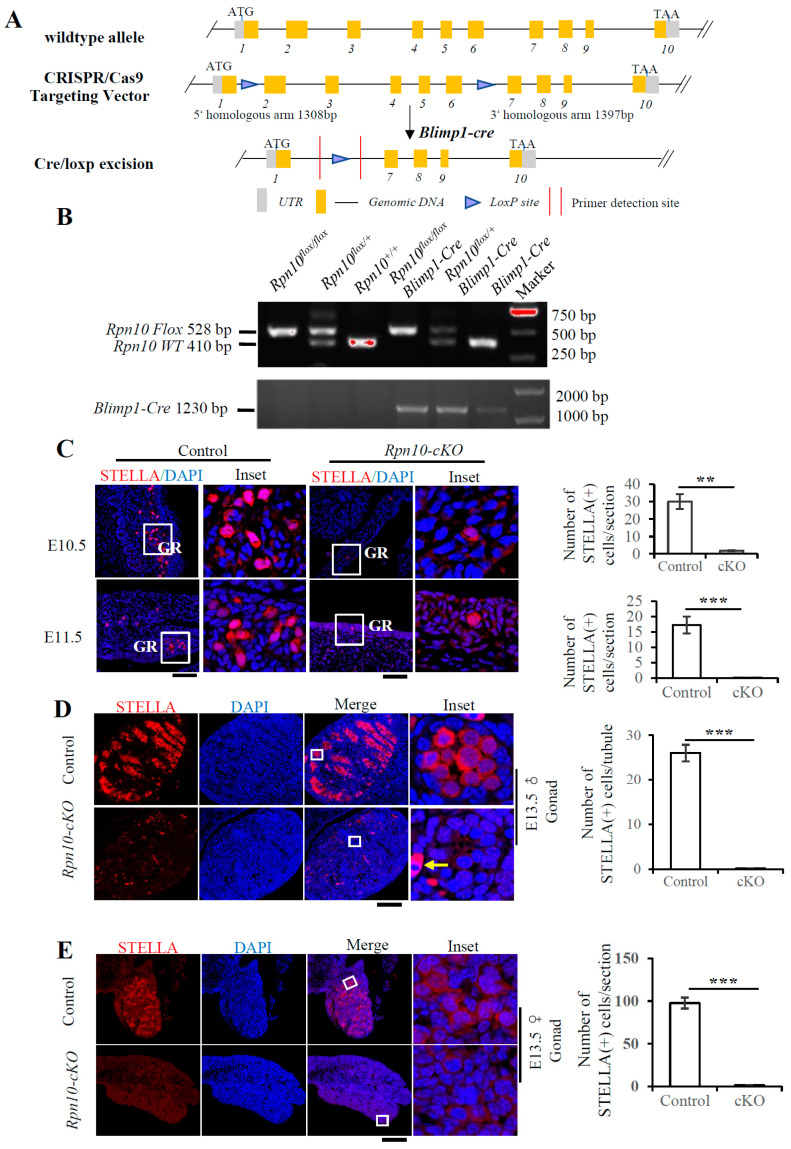 Figure 5
Figure 5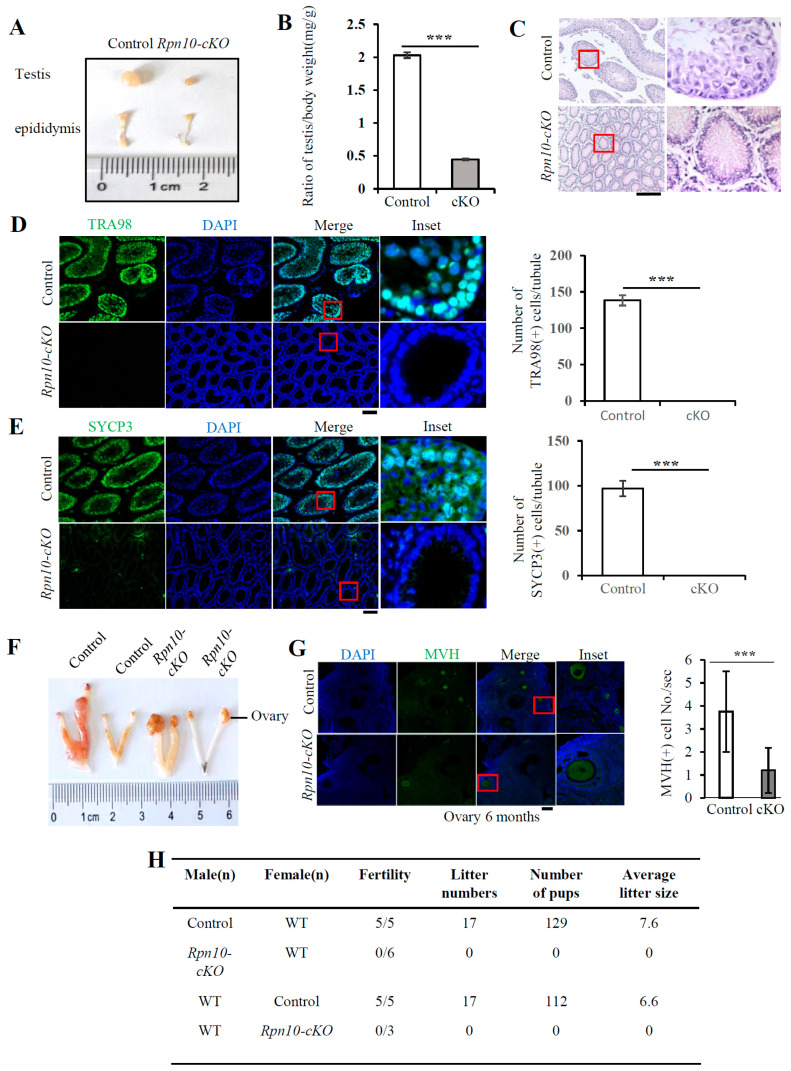 Figure 6
Figure 6- —National Natural Science Foundation of China
- —National Key R & D Program of China
- —Tang scholarship
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsUbiquitin and proteasome pathways · Genetics and Neurodevelopmental Disorders · Epigenetics and DNA Methylation
1. Introduction
Spermatogenesis and oogenesis enable mammals to produce mature sperm and eggs, which are derived from spermatogonial stem cells and primordial follicles, respectively. Both spermatogonial stem cells and primordial follicles initially arise from approximately six primordial germ cells (PGCs) that appear in the proximal posterior ectoderm on embryonic day 6.25 (E6.25) in mice [1]. Following proliferation, specialization, and migration, PGCs localize to the genital ridge and then undergo sex differentiation into gonocytes at E13.5. The female gonocytes go through the preleptotene stage, zygotene stage, and pachytene stage of meiosis and finally stay at the diplotene stage [2]. A portion of the primordial follicles resume meiosis and continue to grow until they enter puberty after birth [3]. The primordial follicles produced by the breakdown of germinal vesicle (GVBD) form primordial follicles pool for their lifelong production. Male gonocytes undergo mitotic arrest after sex differentiation and reenter mitosis until 1–2 days after birth and migrate to the lateral side of the testicular cord, where spermatogonia and spermatocytes are produced [4]. Following two rounds of meiosis, haploid round spermatids are formed, and then mature sperm with acrosome and flagella are produced after spermiogenesis [5].
Proteasomes are responsible for the degradation of most cellular proteins with four types of activators, namely the 19S regulatory particle (RP), PA28α/β, PA28γ, and PA200 [6,7]. The 26S proteasome, which degrades ubiquitinated proteins, is a ~2500 kDa protein complex that consists of the 20S core particle (CP) and the 19S regulatory particle. The CP is composed of α and β subunits, which form a cylindrical structure of heteroheptameric α_1–7_β_1–7_. In the pachytene stage of mammalian mouse testis, a large number of α4 variant α4s appear to form the testis-specific spermatoproteasome [8]. The subunits in the 19S particle control substrate recognition, unfolding, and transport into the open gate of the 20S core particle. The recruitment of the ubiquitinated substrates in the 26S proteasome is mediated directly by the proteasomal ubiquitin receptors, including Rpn1, Rpn10, and Rpn13 [6]. Each of these ubiquitin receptors seems to play distinct roles in binding ubiquitin chains. Rpn10 has two UIM domains, which bind K48 and K63 ubiquitin chains with similar affinity [9], whereas the N-terminal PRU domain of Rpn13 tends to bind the K48-linked ubiquitin chain [10]. Rpn10 and Rpn13 bind distal and proximal ubiquitin, respectively, though they can simultaneously bind the K48-linked ubiquitin chain [11]. On the other hand, Rpn1 might act as a co-receptor of Rpn10 primarily for binding the K63 ubiquitin chain [12]. Rpn10 and Rpn13 have overlapping functions, but Rpn1 is indispensable for development [13]. The conditional deletion of either Rpn10 or Rpn13 in the liver results in mild liver damage, but their simultaneous deletions lead to severe liver damage, suggesting that Rpn10 and Rpn13 play redundant roles in the liver [9]. The global deletion of Rpn13 blocks both oogenesis and spermatogenesis, leading to infertility in mice [14], whereas the global deletion of Rpn10 results in mouse embryonic lethality [15]. However, the specific roles of Rpn10 and Rpn13 in germ cell production remain unclear. We and others have previously identified Rpn13 as a subunit of the mammalian 26S proteasome [16,17,18]. This study demonstrates that the conditional deletion of Rpn10 or Rpn13 in PGCs does not block the formation of PGCs, though leading to infertility in both male and female mice. While the deletion of Rpn10 in PGCs sharply reduces PGC migration, the deletion of Rpn13 in PGCs disrupts the assembly of the 26S proteasome, reduces the number of PGCs, and blocks the meiosis of spermatocytes at the zygotene stage during prophase I.
2. Materials and Methods
2.1. Animals
The C57BL/6J mice of Rpn13^flox/+^ were provided by Nanjing Biomedical Research Institute of Nanjing University. The C57BL/6N mice of Rpn10^flox/+^ were provided by Biocytogen Pharmaceuticals Co., Ltd. (Beijing, China); Blimp1-Cre mice were kindly provided by Professor Jinsong Li. All mice were housed in specific pathogen-free-grade animal houses under alternating conditions of 12 h of light and 12 h of darkness.
Rpn13^flox/flox^ mice were obtained by mating Rpn13^flox/+^ and Rpn13^flox/+^ mice. Rpn13^flox/flox^ and Rpn13^flox/+^Blimp1-Cre were further mated to generate Rpn13^flox/flox^Blimp1-Cre (Rpn13-cKO), Rpn13^flox/+^, Rpn13^flox/flox^, and Rpn13^flox/+^Blimp1-Cre (Rpn13-control) mice. Rpn10-cKO and control mice were obtained by similar processes. For genotyping, DNA was extracted from the mouse tail or tissue and analyzed by PCR (98 °C, 2 min; 98 °C,10 s, 60 °C, 20 s, 72 °C, 30 s, 35 cycles; 72 °C, 5 min). The primers used are listed in Table S1.
2.2. Tissue Collection and Immunofluorescence or Immunohistochemical Staining
Embryos or gonads were fixed in 4% paraformaldehyde at 4 °C, dehydrated, embedded in paraffin, and sectioned (5 μm). The sections were dewaxed, hydrated, and boiled in the antigen retrieval buffer (10 mM sodium citrate, pH 6.0) using a microwave oven for about 10–15 min. Then, the immunohistochemical staining was performed as per the instructions of the general SP kit SP-9000 (Beijing Zhongshan-Golden Bridge Biological Technology Co., Ltd., Beijing, China). Immunofluorescence analysis was performed following washing the sections in PBS three times and blocking with goat serum in 0.2% TritonX-100. The sections were then incubated with primary antibodies at 4 °C overnight. After washing with PBS three times, the sections were incubated with secondary antibodies for 1 h at room temperature. Next, they were mounted with DAPI (5 μg/mL) after washing with PBS three times. The images were taken under a regular microscope or a confocal microscope (Zeiss, Oberkochen, Germany, LSM 700, or LSM 880). The primary and secondary antibodies used are listed in Table S2.
2.3. Immunoblotting
Testes were ground in a mortar, sonicated, and then centrifugated at 16,000× g for 10 min. The proteins were separated by SDS-PAGE or by Native-PAGE, followed by incubating on ice for 30 min. For SDS-PAGE, the testes were homogenized and lysed in the buffer (50 mM Tris-HCl, pH8.0, 100 mM KCl, 1 mM EDTA, 1 mM EGTA, 1% TritonX-100, 2.5 mM Na_4_P_2_O_7_, 1 mM β-Glycerophosphate, 1 mM Na_3_VO_4_, and a protease inhibitor mixture). As for native-PAGE, testes were homogenized and lysed in the buffer (50 mM Tris-HCl, PH7.5, 250 mM Sucrose, 5 mM MgCl_2_, 0.5 mM EDTA, 2 mM ATP, 1 mM DTT). After proteins were transferred to the polyvinylidene fluoride (PVDF) membrane (Millipore, Burlington, MA, USA), they were blocked with 5% non-fat milk and incubated with primary antibodies at 4 °C overnight. The secondary antibodies were horseradish peroxidase-labeled anti-rat, anti-rabbit, or anti-mouse IgG. The primary and secondary antibodies used for immunoblotting are listed in Table S2.
2.4. Hematoxylin and Eosin (H & E) Staining
Testes and ovaries were fixed in 4% paraformaldehyde at 4 °C overnight, dehydrated in 50–100% gradient ethanol, transparentized in xylene twice, dipped in warm wax four times, embedded in paraffin, and sectioned (5 μm). Then, the sections were dewaxed with xylene three times until the paraffin was removed, hydrated with ethanol and H_2_O three times, and stained with hematoxylin and eosin. The nucleus was stained with hematoxylin (blue), and the cytoplasm was stained with eosin (red).
2.5. Proteasome Activity Assay, Spermatocyte Spread and Immunolabeling
The proteasome activity assay, spermatocyte spread, and immunolabeling were performed as described previously [19]. The proteasome activity was analyzed using the peptide substrate succinyl-Lue-Leu-Val-Tyr-7-amino-4-methylcoumarin (Suc-LLVY-AMC). The primary and secondary antibodies used for immunolabeling are listed in Table S2.
2.6. TUNEL Assay
Apoptosis was detected by the TUNEL assay that was performed according to the DeadEnd Fluorometric TUNEL system (Promega, Madison, WI, USA, G3250) protocol.
2.7. Quantification and Statistical Analysis
All data were presented as the mean ± standard error of the mean (SEM). Unless stated otherwise, the significance level between the control and the conditional knockout groups was determined by the two-tailed unpaired t test, and the statistical significance was defined as p < 0.05 (n.s., not significant; * p < 0.05; ** p < 0.01; *** p < 0.001). All of the images were chosen blind and randomly and quantitated by Image J (ImageJ 1.52n).
3. Results
3.1. Conditional Deletion of Rpn13 in PGCs Reduces the PGC Number at E10.5
Germ cell formation is rigorously regulated by the ubiquitin–proteasome pathway. The global deletion of the proteasomal ubiquitin receptor Rpn13 leads to infertility in both male and female mice [14]. Primordial germ cells (PGCs) are the sources of hermaphroditic germ cells and undergo germline lineage restriction, specification, migration, and proliferation. To explore the mechanism underlying the role of Rpn13 in reproduction, we selectively knocked out the Rpn13 gene in PGCs. Using CRISPR-Cas9-assisted homologous recombination [20], two LoxP sites were inserted, respectively, into intron 2 and intron 5 of the Rpn13 gene to generate mice carrying the flox allele (Rpn13^flox/+^) (Figure 1A).
Although Blimp1 is a key transcription factor controlling terminal plasma cell differentiation [21], it is essential for the formation of PGCs [1]. To disrupt the Rpn13 gene in PGCs, the Rpn13^flox/+^ mice were further mated with Blimp1-Cre mice, which express the Cre recombinase in PGCs driven by a Blimp1 promoter as early as E6.25 [1]. Six genotypes of Rpn13 alleles, including Rpn13^+/+^, Blimp1-Cre, Rpn13^flox/+^, Rpn13^flox/flox^, Rpn13^flox/+^Blimp1-Cre, and Rpn13^flox/flox^Blimp1-Cre, were generated by mating the Rpn13^flox/flox^ with Rpn13^flox/+^Blimp1-Cre mice, as verified by PCR (Figure 1B). Rpn13 was conditionally deleted in PGCs in Rpn13^flox/flox^Blimp1-Cre mice (i.e., Rpn13-cKO), but not in Rpn13^flox/+^, Rpn13^flox/flox^ and Rpn13^flox/+^Blimp1-Cre mice. Stella (also known as Dppa3 or PGC7), a gene expressed in lineage-restricted germ cells, can usually mark PGCs [22]. The total number of Stella-positive PGCs had no difference between the control and the Rpn13-cKO embryo at E8.5 and E9.5 but was markedly reduced at both E10.5 and E11.5 in the Rpn13-cKO embryo (Figure 1C). Thus, the deletion of Rpn13 reduces the number of PGCs starting from E10.5, though it does not completely deplete PGCs.
3.2. Conditional Deletion of Rpn13 Disrupts Germ Cell Development and Leads to Infertility in Both Males and Females
We further found that both male and female Rpn13-cKO mice were infertile (Supplementary Figure S1A). Rpn13-deficient testes were much smaller with thinner seminiferous tubules, though the sizes of their epididymis were only slightly reduced (Figure 2A,B). Hematoxylin and eosin (H & E) staining showed that the seminiferous tubules in Rpn13-cKO testes did not contain any mature sperm, which usually possesses a flagellum- and a falciform-shaped head with the nucleus and the acrosome [23,24] (Figure 2C and Supplementary Figure S1E). Immunostaining with the germ cell marker antibody TRA98 [25] showed that the number of germ cells in seminiferous tubules reduced dramatically (Figure 2D,E and Supplementary Figure S1C–F). There were no detectable germ cells on most seminiferous tubule sections, though a few of them were present on certain sections. The germ cells in testes and ovaries at postnatal day 1 (P1), as marked by the TRA98 and Vasa homolog (MVH), respectively, all decreased dramatically in Rpn13-cKO mice (Figure 2E and Supplementary Figure S1B). MVH usually begins to be expressed in oocytes after PGCs colonize at the gonads and is detectable throughout the entire life of oocytes after E12.5 [26]. We found that the ovary of the Rpn13-cKO mice at 5 weeks was smaller than that of the control group (Figure 2G and Supplementary Figure S1G) with fewer MVH-positive follicles and oocytes (Figure 2H,I and Supplementary Figure S1G,H) in the Rpn13-cKO mice. These results demonstrate that the deletion of Rpn13 in PGCs disrupts germ cell development, leading to infertility in both male and female mice.
3.3. Deletion of Rpn13 Blocks Meiosis of Spermatocytes at Zygotene Stage During Prophase I
During spermatogenesis, spermatogonia undergo mitosis and differentiation into primary spermatocytes, which further undergo meiosis, leading to the formation of haploid spermatids and sperm [27]. The number of undifferentiated spermatogonia marked by the promyelocytic leukemia zinc-finger protein (PLZF) [28] had no difference between the control and Rpn13-cKO groups (Figure 3A). The synaptonemal complex protein 3 (SYCP3) is partially retained along chromosome arms until metaphase I, while SYCP1 is only just present in autosomes [29]. The number of SYCP3-positive cells decreased sharply (Figure 3B), and spermatocytes marked with either SYCP3 or SYCP1 were only detectable until the zygotene stage, but not the pachytene stage, during prophase I in Rpn13-cKO testes (Figure 3C and Supplementary Figure S2A). The proportions of preleptotene/leptotene, zygotene/zygotene-like, and pachytene were 26.7%, 17.8%, and 55.5%, respectively, in the control group, but their proportions became 90.8%, 9.2%, and 0%, respectively, in the Rpn13-cKO group (Figure 3D). The expression of phosphorylated H2AX (γH2AX) is usually strong at the leptotene stage, gradually weakened at the zygotene stage, and enriched in sex-body at the pachytene stage [30]. However, no γH2AX-positive sex-body was found, while the levels of γH2AX at the zygotene stage were reduced in Rpn13-cKO testis (Figure 3E and Supplementary Figure S2B). Finally, the number of apoptotic cells detected by the TUNEL assay in the TRA98-positive cells increased markedly, suggesting that Rpn13 cKO increases the ratio of apoptosis in spermatocytes (Figure 3F). Together, these results indicate that the conditional deletion of Rpn13 in PGCs blocks meiotic progression at the zygotene stage of prophase I in spermatocytes.
3.4. Conditional Deletion of Rpn13 Markedly Reduces Activity of the 26S Proteasome
The deletion of Rpn13 in PGCs dramatically reduced the levels of Rpn13 in testes (Figure 4A), apparently due to the depletion of Rpn13 in germ cells, which are the primary group of cells in the testis. Using the specific proteasomal fluorogenic peptide substrate, Suc-LLVY- AMC, we found that the activities of the 26S proteasome were sharply reduced in the Rpn13-cKO testes (Figure 4B,C). The deletion of Rpn13 in PGCs dramatically reduced the levels of 19S subunits, including the ubiquitin receptors (Rpn13, Rpn1, and Rpn10) and deubiquitinating enzymes (UCH37, Usp14, and Rpn11), in addition to the proteasome activator PA200, and increased the levels of 20S subunits, including β1, β7 and the spermatoproteasome subunit α4s, as analyzed by immunoblotting following native-PAGE (Figure 4D). In contrast, SDS-PAGE displays a decrease in the total level of β7, including the free form of the β7 subunit (Figure 4A). There is probably a negative feedback regulation of the free β7 expression caused by the increase in β7 in the assembled 26S proteasome. These results suggest that the conditional deletion of Rpn13 reduces proteasomal activities, probably by disrupting the assembly of the 26S proteasome.
3.5. Deletion of Rpn10 in PGCs Sharply Reduces PGC Migration and Leads to Infertility
Rpn10 and Rpn13 were suggested to play redundant roles in recognizing ubiquitinated proteins and maintaining cellular homeostasis [9]. The global deletion of Rpn10 leads to early-embryonic lethality in mice [15], but the specific role of Rpn10 in germ cell development remains unclear. Thus, we constructed mice with the conditional deletion of Rpn10 in PGCs. Two LoxP sites were inserted in non-coding regions between exons 1 and 6 of the Rpn10 gene by CRISPR-Cas9. The deletion of the Rpn10 gene in PGCs was achieved using Blimp1-Cre for the selective expression of Cre recombinase in PGCs (Figure 5A,B). Except for the Rpn10-cKO mice with the genotype of Rpn10^flox/flox^Blimp1-Cre, mice with other genotypes including Rpn10^+/+^, Blimp1-Cre, Rpn10^flox/+^Blimp1-Cre, Rpn10^flox/+^, and Rpn10^flox/flox^ were also obtained. PGCs were only sporadically found at the genital ridges at E10.5 and E11.5 in Rpn10-cKO mice, suggesting that PGCs were mostly lost along the migration process in Rpn10-cKO mice (Figure 5C). Furthermore, there were few Stella-positive gonocytes in male and female gonads from the Rpn10-cKO mice at E13.5, as analyzed by immunostaining (Figure 5D,E). These results suggest that the specific deletion of Rpn10 in PGCs sharply reduces the migration of PGCs.
The testis of male Rpn10-cKO mice was apparently much smaller on postnatal day 32 (Figure 6A,B). H & E staining showed that the mouse testicular seminiferous tubules were essentially empty except for a ring of cells near the basement membrane in the Rpn10-cKO testis (Figure 6C). Immunofluorescence staining with the germ cell marker TRA98 revealed that there were no germ cells in the seminiferous tubules of the Rpn10-cKO testis (Figure 6D). Immunofluorescence staining with SYCP3 demonstrated that there were no spermatocytes in the Rpn10-cKO testis (Figure 6E). Thus, the conditional deletion of Rpn10 in PGCs leads to the loss of spermatocytes.
The ovaries of Rpn10-cKO mice were larger than those of the control mice at approximately 18 months after birth (Figure 6F), but the numbers of follicles and oocytes, which are MVH-positive, were markedly reduced in the ovaries from Rpn10-cKO female mice (Figure 6G and Supplementary Figure S3). Consequently, there were no offspring mice following the mating of Rpn10-cKO female mice with wild-type male mice or the mating of wild-type female mice with Rpn10-cKO male mice (Figure 6H). These results suggest that Rpn10 deletion causes infertility in both male and female mice, primarily by sharply reducing PGC migration.
4. Discussion
The conditional deletion of Rpn10 or Rpn13 alone in the liver causes minor damage, in contrast to the severe liver damage caused by their double deletion, suggesting their redundant roles in liver development [9]. Our study shows that unlike the reduced PGC migration caused by the deletion of Rpn10 in PGCs, the deletion of Rpn13 in PGCs primarily blocks the meiosis of spermatocytes at the zygotene stage during prophase I, suggesting their non-redundant roles in germ cell formation. The role of Rpn13 in germ cells is obviously different from that in the liver, and the reason for this could be its different mechanisms in regulating the 26S proteasome. These results strongly support the notion that the roles of Rpn10 and Rpn13 vary with tissue or biological processes.
The global deletion of Rpn13 blocks both oogenesis and spermatogenesis [14], but the specific role of Rpn13 in germ cell production remains unclear. Our study demonstrates that Rpn13 is required for germ cell production in both testes and ovaries. As analyzed in the testes, the conditional deletion of Rpn13 reduces proteasomal activities, probably by disrupting the assembly of the 26S proteasome. It has been shown previously that the global deletion of Rpn10 leads to embryonic lethality in mice [15]. We show here that while the number of PGCs reduced at E10.5-11.5, very few PGCs remained at E13.5 in the mice with conditional deletion of Rpn10 in PGCs, suggesting that Rpn10 is required for the programmed migration of PGCs in the embryo.
5. Conclusions
The deletion of Rpn13 in PGCs disrupts the assembly of the 26S proteasome, reduces the number of PGCs, and blocks meiosis of spermatocytes at the zygotene stage during prophase I, whereas the deletion of Rpn10 in PGCs sharply reduces PGC migration. The numbers of germ cells in the testes and ovaries all decreased dramatically in Rpn13-cKO mice. The conditional deletion of either Rpn10 or Rpn13 in PGCs results in infertility in both male and female mice. In conclusion, Rpn10 and Rpn13 play non-redundant essential roles in germ cell formation and fertility in both male and female mice. These results are important for understanding the roles of Rpn10 and Rpn13 in germ cell development and related reproductive diseases.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Ohinata Y. Payer B. O’Carroll D. Ancelin K. Ono Y. Sano M. Barton S.C. Obukhanych T. Nussenzweig M. Tarakhovsky A. Blimp 1 is a critical determinant of the germ cell lineage in mice Nature 200543620721310.1038/nature 0381315937476 · doi ↗ · pubmed ↗
- 2Western P.S. Miles D.C. van den Bergen J.A. Burton M. Sinclair A.H. Dynamic regulation of mitotic arrest in fetal male germ cells Stem Cells 20082633934710.1634/stemcells.2007-062218024419 · doi ↗ · pubmed ↗
- 3He M. Zhang T. Yang Y. Wang C. Mechanisms of Oocyte Maturation and Related Epigenetic Regulation Front. Cell Dev. Biol.2021965402810.3389/fcell.2021.65402833842483 PMC 8025927 · doi ↗ · pubmed ↗
- 4de Rooij D.G. Russell L.D. All you wanted to know about spermatogonia but were afraid to ask J. Androl.20002177679810.1002/j.1939-4640.2000.tb 03408.x 11105904 · doi ↗ · pubmed ↗
- 5Mc Laren A. Meiosis and differentiation of mouse germ cells Symp. Soc. Exp. Biol.1984387236400220 · pubmed ↗
- 6Jiang T.-X. Zhao M. Qiu X.-B. Substrate receptors of proteasomes Biol. Rev. Camb. Philos. Soc.2018931765177710.1111/brv.1241929732666 · doi ↗ · pubmed ↗
- 7Finley D. Recognition and processing of ubiquitin-protein conjugates by the proteasome Annu. Rev. Biochem.20097847751310.1146/annurev.biochem.78.081507.10160719489727 PMC 3431160 · doi ↗ · pubmed ↗
- 8Qian M.-X. Pang Y. Liu C.H. Haratake K. Du B.-Y. Ji D.-Y. Wang G.-F. Zhu Q.-Q. Song W. Yu Y. Acetylation-mediated proteasomal degradation of core histones during DNA repair and spermatogenesis Cell 20131531012102410.1016/j.cell.2013.04.03223706739 PMC 3983474 · doi ↗ · pubmed ↗