Neurophysiological Markers of Reward Processing Can Inform Preclinical Neurorehabilitation Approaches for Cognitive Impairments Following Brain Injury
Miranda Francoeur Koloski, Reyana Menon, Victoria Krasnyanskiy

TL;DR
This paper explores how brain activity related to reward processing can guide neurostimulation treatments for cognitive issues after brain injury.
Contribution
The paper introduces electrophysiological markers, like cortico-striatal beta oscillations, as potential targets for neurorehabilitation after TBI.
Findings
Electrophysiology can detect cognitive deficits missed by structural measures.
Cortico-striatal beta oscillations are disturbed after frontal TBI and may represent subjective value during reward processing.
Targeting these oscillations with stimulation could improve decision-making post-TBI.
Abstract
Brain stimulation therapies may be used to correct motor, social, emotional, and cognitive consequences of traumatic brain injury (TBI). Neuromodulation applied with anatomical specificity can ameliorate desired symptoms while leaving functional circuits intact. Before applying precision medicine approaches, preclinical animal studies are needed to explore potential neurophysiological signatures that could be modulated with neurostimulation. This review discusses potential neural signatures of cognition, particularly reward processing, which is chronically impaired after brain injury. Electrophysiology, compared to other types of biomarkers, can detect deficits missed by structural measures, holds translational potential between humans and animals, and directly informs neuromodulatory treatments. Disturbances in oscillatory activity underscore structural, molecular, and behavioral…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
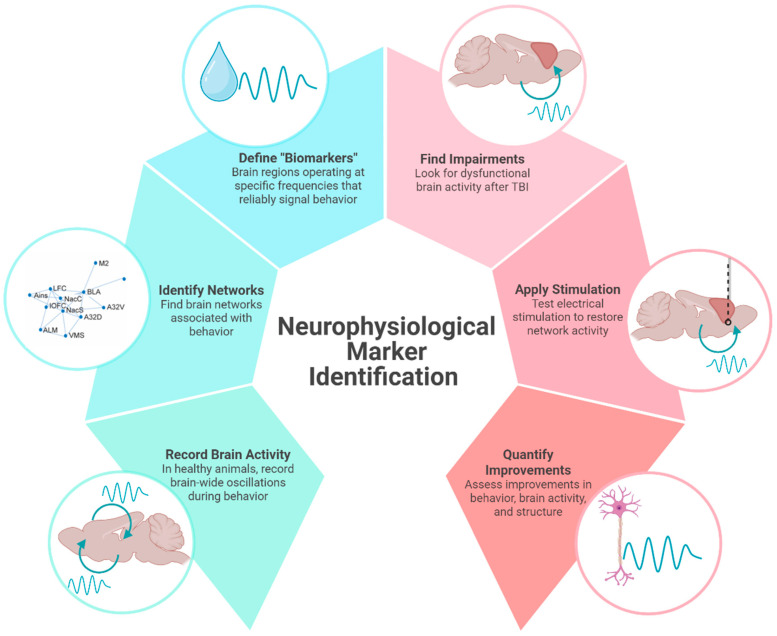 Figure 1
Figure 1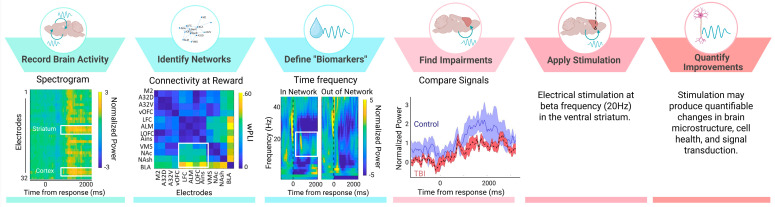 Figure 2
Figure 2- —VA Office of Research and Development
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsEEG and Brain-Computer Interfaces · Functional Brain Connectivity Studies · Neural and Behavioral Psychology Studies
1. Introduction
Biomarkers serve as indicators of biological processes that are objective and reproducible [1]. Categories of biomarkers include neuroimaging (magnetic resonance imaging (MRI)), neurophysiological (Electroencephalography (EEG)), biofluid (blood), and digital biomarkers (wearables), all of which are being implored to study traumatic brain injury (TBI). A number of TBI biomarkers have been cataloged, including serum-based protein, exosomal microRNAs, and metabolic indicators [2,3]. However, these molecular markers are limited in temporal precision and ability to reflect ongoing changes in brain activity, particularly in relation to cognition. Structural imaging (MRI) often fails to detect mild injuries. Although, new imaging advancements like diffusion tensor imaging have improved sensitivity to detect microstructural damage (axonal injury) [4,5]. To address these limitations, the impact of injury on neurophysiological activity (neural oscillations) can be examined. The search for biomarkers in TBI is extremely difficult given the heterogeneous nature of brain injury and the symptoms it produces. Even a single injury propagates multiscale disruptions to local and distributed brain areas. Since behavioral outcomes are hard to predict, establishing a reliable biomarker of injury that can estimate injury progression and response to treatment will be useful.
Brain injury triggers a cascade of insults to the central nervous system including a combination of inflammatory, cellular, and molecular changes [6,7,8]. Due to its superficial position in the brain, prefrontal cortex is particularly susceptible to injury [9,10,11]. Cortical damage leads to changes in reward-guided behavior, including impaired decision making, decreased motivation, impulsivity, and interrupted reinforcement-learning [11,12,13]. Most symptoms are resolved; however, in 5–15% of cases, cognitive deficits endure for years after injury [10,11]. Chronic symptoms are a result of cortical neuron loss, disconnection with networks, and neurotransmitter dysregulation [14,15,16].
The mechanism of injury determines if damage is focal (hematoma, hemorrhage, contusion), diffuse (axonal injury), or a combination. The most common type of injury is diffuse damage to the white matter tracts caused by shearing [17,18,19]. Diffuse axonal injury often fractures cortical-subcortical connectivity that can afflict cognitive networks and perpetuate neuropsychiatric symptoms [18,20]. The symptom profile is dependent on the neural pathways impacted. For example, thalamo-cortical tract damage accounts for significant alteration in executive dysfunction [21].
The goal of TBI treatment is to develop a precision medicine approach that targets a specific brain area or brain function (neurophysiological biomarker) that can be individualized based on each unique trauma pathology. Most treatment strategies center around addressing motor functions or physical impairments through rest and rehabilitation [22], leaving long-term cognitive deficits unaddressed. TBI-induced motor deficits, including gait, coordination, fine motor skills, seizures, and muscle tremors, can be alleviated with a combination of rehabilitation, brain stimulation, and pharmacologic strategies (muscle relaxation/motor inhibition drugs) [23,24,25]. There are no pharmacological treatments that specifically regulate TBI-related changes in mood and cognition or prevent the onset of neuropsychiatric disorders (which have a 2–3 times greater risk of developing) [11,16,26].
In line with a precision medicine approach, using neuromodulation to provide electrical stimulation is thought to activate specific neural circuits, which can strengthen its functional network to improve desired behaviors. Identifying a neurophysiological biomarker associated with specific behaviors is needed to improve the precision and reliability of neuromodulation. As opposed to pharmacological interventions, neuromodulation has the potential to influence a multitude of brain areas while maintaining specificity for functional circuits. This review aims to synthesize findings across preclinical and clinical studies related to neurophysiological biomarkers in TBI, specifically oscillatory associated with reward-guided decision-making impairments seen chronically after injury. We provide a theoretical framework by which preclinical studies can support large-scale electrophysiological recordings in healthy animals to identify brain networks and potential “biomarkers” that can be targeted with electrical stimulation in injured animals (Figure 1). While not a formal meta-analysis, our narrative review offers converging evidence to support the translational potential of using electrophysiological signatures to identify new therapeutic targets for TBI.
2. Neurophysiological Markers Can Identify Potential Therapeutic Targets
2.1. Brain Oscillations and Their Disruption After Injury
Electrophysiological measures, including EEG, can detect neural changes after even a mild TBI that structural imaging may fail to detect [4,22]. For example, EEG can detect reduced amplitude and slowing of brain waves in the acute and subacute phases of mild TBI of patients without visible structural damage on MRI/CT scans [27]. The benefits of electrophysiological measures include good temporal resolution, translatability between humans and animals, affordability, usefulness during recovery, and the capacity to be paired with cognitive tasks [1,12]. Oscillatory activity in an EEG reflects coordinated neural activity that facilitates communication within and between brain areas. Oscillatory bands, which are operationally defined based on functional brain states, range from 0.5 to 500 Hz [28]. The average ionic movements (including synaptic activity, calcium fluctuations, spike after-hyperpolarization, gap junctions, glial, etc.) recorded from an electrode are known as the local field potential (LFP) [28]. LFP activity patterns are associated with distinct behaviors and can predict disease states or response to treatment [28,29].
Numerous studies have identified persistent abnormalities in oscillatory dynamics following TBI, including theta suppression during working memory tasks [30], delta elevations during wakefulness [31], and reductions in beta/gamma frequencies during cognition [32]. Generally, lower frequencies (8–30 Hz) are thought to reflect top–down information processing driven by thalamo-cortical connections and higher frequencies (40–100 Hz) to drive bottom–up processing [33,34]. Importantly, connectivity data, reflecting widespread changes across networks, better predict symptoms severity than local changes in power, suggesting brain injury should be treated as a network disorder (opposed to focal injury) [5]. Changes in oscillations are identified after even mild TBI and therefore may be sensitive to the underlying neural alterations associated with injury [5]. Thus, neurophysiological signals have the potential to aid in TBI diagnosis, prognosis, and treatment response monitoring [1,22,35].
In both humans and animals, TBI disrupts brain oscillations. More recently, studies have shown that electrical stimulation can restore oscillatory rhythms to improve cognitive outcomes following TBI. Generally, after injury, there is a decrease in power across frequency bands which returns to baseline after the acute injury response [1,36]. However, deficits in brain activity can persist chronically (>10 weeks) [36] and, thus, may represent a therapeutic window for clinical intervention. In patients with TBI, altered EEG activity can be seen years after injury [12,32,37,38]. Specifically, alterations of thalamo-cortical circuits are associated with persistent cognitive symptoms seen with TBI [5,21]. The dorsolateral prefrontal cortex, which is normally engaged during cognition, shows reduced power and EEG abnormalities after TBI [8]. Electrophysiological abnormalities occur across brain states (rest, sleep, etc.) and are consistent enough to distinguish between injured and non-injured patients and predict patient outcomes [32,38].
2.2. Neuromodulation to Restore Brain Activity
Modulating brain activity through electrical stimulation can restore rhythmic patterns of the brain disrupted by TBI [39]. To address the widespread neural impairments, stimulation would need to restore activity beyond the focal injury site. Functional MRI in humans and electrophysiological recordings in rodents show that brain stimulation emulates changes across networks [40].
Methods of neuromodulation vary in their level of invasiveness, continuous or transient application, and pairing with functional behavior or rest. Stimulation protocols can be open-loop (applied at/for a certain amount of time, not tied to a behavior or brain function), “on-demand” (triggered by a behavioral event), or closed-loop (triggered by brain feedback) [41,42]. High-frequency stimulation (>5 Hz) is thought to induce excitability and increase cerebral blood flow, whereas low-frequency stimulation (<1 Hz) typically induces inhibitory effects [43]. Factors such as the type of neuromodulation, duration of stimulation, and regularity required will determine the best target for neuromodulation.
Neurostimulation in the form of transcranial magnetic stimulation (TMS), transcranial direct current stimulation (tDCS), and deep brain stimulation (DBS), applied in preclinical models have successfully ameliorated behavioral symptoms of TBI, including motor, attention, memory, mood, and impulsivity impairments [35,44]. In preclinical studies, stimulation applied immediately to several weeks after trauma improved behavioral outcomes, whereas clinical applications of neuromodulation are not usually introduced until later stages of recovery [44]. Repeated TMS applied to the left dorsolateral prefrontal cortex has FDA approval to treat depression and therefore may also be useful in TBI patients exhibiting mood dysregulation, central pain, and blunted cognition or affect [43,45,46]. Importantly, compared to other interventions, growing evidence supports that TMS can have a long-lasting impact on neural circuits by inducing plasticity [47].
TMS has been used in rodent models with non-invasive coils, but the translational potential is obscured by stimulation site specificity and the use of restraints or anesthesia [48,49]. Instead, “TMS-like” approaches that use implanted electrodes opposed to topical coils, may be optimal when designing preclinical studies [35]. “TMS-like” protocols of 20 Hz stimulation trains applied repeatedly for 10 days to prelimbic cortex normalize depressive-like behaviors and reduce brain-derived neurotrophic factor levels in reward-related regions [50].
2.3. How Does Stimulation Restore Brain Function Following TBI?
Axonal shearing, swelling, chronic inflammation, and microglia activation perpetuate neural loss and tissue degeneration after trauma [22,51,52,53]. Stimulation provides neuroprotection, decreases apoptosis, reduces inflammation, directs cerebral blood flow, and induces neuroplasticity to improve cell health and restore neural circuits [35,43,44]. White matter regeneration (which may be particularly important for cognition) is achieved through optogenetic, pharmacogenetic, or indirect brain stimulation by encouraging glial cells to myelinate active axons [54,55]. In preclinical models of TBI, reversed cortical tissue loss, white matter regeneration, and increased intracellular signaling are directly related to behavioral improvements [56]. TDCS and TMS increase c-fos expression (a marker of neural activity) [57]. TDCS also increases brain-derived neurotrophic factor (BDNF) in stimulated cortical regions of injured brains, indicating a focal neuroplasticity response that is associated with cognitive improvements in spatial memory [58]. Similarly, TMS reduces glial fibrillary acidic protein (GFAP) expression, associated with astroglia cells and neural regeneration [2,59]. Improved neural health and signal transduction can re-establish physiological patterns. DBS reduces spontaneous neural firing and encourages activation of efferent pathways by increasing the release of GABA from interneurons to restore the excitatory/inhibitory balance [45,60].
3. A Potential Biomarker of Reward-Processing Deficits in TBI
The lack of mechanistic information regarding failure of cognitive networks precludes the utility of neuromodulation to treat chronic TBI deficits. Precision medicine approaches to treat TBI depend on finding neurophysiological biomarkers that reliably mark discrete cognitive functions like decision-making, planning, and memory.
3.1. Preclinical Models of Frontal TBI
Rodent models provide information about neurobiological mechanisms which are necessary to inform clinical interventions. Aside from notable differences in brain size, relative volume of brain regions, and presence/absence of cortical folding, the architecture and functional networks are largely preserved between humans and rodents [61]. Animal models of TBI offer control over injury specifications, ability to monitor injury progression, and invasive techniques that are not feasible in humans [62]. Limitations in these models include lack of continuity between protocols, challenge replicating the heterogeneous nature of TBI, broad timeframe of injury (acute, subacute, and chronic), and inability to model exact biomechanical parameters of injury [37,48]. Despite these limitations, rodent models of frontal TBI produce robust and predictable cognitive deficits [62]. Bilateral frontal controlled cortical impact (CCI) injury reliably produces impairments in impulse control and decision-making [63].
Animal models also enable large-scale recordings of in vivo brain activity. Multi-site electrodes simultaneously capture field potentials at different sites across, or between, networks. “Brain-wide” recordings measuring activity from up to 32 brain areas simultaneously, can be used to characterize networks operating at distinct oscillatory frequencies to support unique behaviors [64,65,66]. Although these electrodes record from within the brain, the information they provide (large-scale measure of brain oscillations) is like human EEG. The following section provides an example of how this technique can identify neurophysiological biomarkers and potential targets for neuromodulation.
3.2. Deficient Cortical Beta Oscillations Indicate Reward-Processing Issues After TBI
As shown in humans and animals, an injury to prefrontal cortex results in chronic executive function deficits, including attention, memory, and reward-guided decision making [13,67,68]. Damage causes lasting disruption to brain circuits through inflammation, gliosis, cell death, and alterations in microstructure, ultimately effecting neural communication. Changes in brain oscillations perpetuate disruptions throughout functional networks after cortical damage. Related to executive function deficits, fronto-parietal (attention) and fronto-striatal (decision-making) networks show reduced electrophysiological responses and connectivity consistent with slower reaction times and poor decision making [69,70].
Oscillatory activity, particularly at beta frequencies, deriving from the prefrontal cortex is important for top–down attention [71], executive control [72], sensorimotor integration [73], motor planning [74], and decision making [75]. Beta oscillations are correlated with the cortical microstructure (i.e., myelin density/integrity) and therefore susceptible to damage through TBI [76]. Even mild TBI leads to localized and widespread disruptions in beta oscillatory activity (measured by magnetoencephalography) [5]. Beta activity in cortico-striatal regions (prefrontal cortex, orbitofrontal cortex, anterior insula, ventral striatum, and basolateral amygdala) marks positive valence (i.e., rewarded outcomes) [77,78,79,80]. In humans, reward-evoked beta oscillations correlate with activation of (and coupling between) ventral striatum and medial prefrontal cortex, suggesting that beta frequency oscillations may coordinate the neural circuits involved in reward processing [80]. In rats, across multiple tasks of reward-guided decision-making, we consistently find increases in beta-frequency (15–30 Hz) oscillations during reward processing that reflects reward magnitude, reward probability, and subjective value [77].
Due to its superficial position in the brain, cortex is particularly susceptible to injury [10], giving rise to socially inappropriate behavior, poor impulse control, and trouble decision-making [21]. When cortical areas are damaged, their participating networks are also dysfunctional. As a network, cortico-striatal brain areas mediate adaptive reward-guided decision-making by creating action-outcome associations, controlling impulsive choices, and responding flexibly to changing conditions [81]. Bifrontal TBI caused by CCI decreases the ability of rats to detect reinforced outcomes, impairs behavioral flexibility, and increases impulsivity [63,82].
Consistent with these behavioral changes, we find that CCI-TBI also blunts reward-locked beta oscillations and cellular activity in lateral orbitofrontal cortex (measured by c-Fos staining) [82]. Beta activity in the orbitofrontal cortex does not discriminate between reward outcomes as efficiently following injury (effects observed up to 12 weeks after injury). Beta oscillations are correlated with structural deficits observed after TBI, including myelin density and morphology [76]. Local changes in beta power and deficits in functional connectivity following TBI has also been characterized in thalamo-cortical circuits [5,30,83]. Interestingly, we found that reductions in orbitofrontal activity were rescued by a behavioral intervention in which reinforced outcomes were cued, thus suggesting reward signals in the brain are malleable and can be altered to improve cognitive behavior [82]. Brain-based interventions may work similarly to increase beta oscillations during positive reward outcomes thereby reversing reward-related deficits from TBI. For these reasons, reward-evoked beta oscillations in the cortico-striatal network may represent a neurophysiological marker that can be targeted with electrical stimulation to improve cognitive symptoms of TBI. In accordance with the theoretical framework for neurophysiological marker identification (Figure 1), we have shown that large-scale recordings of brain activity can identify behaviorally relevant networks to define putative “biomarkers” with region and frequency specificity (Figure 2). We have found impairments of reward-evoked beta oscillations following TBI that may be remediated by electrical stimulation (Figure 2).
Although, we have not tested electrical stimulation on this putative beta signal in TBI animals, we have stimulated beta oscillations to influence decision making in healthy animals. Beta oscillations were modified in healthy rats performing a delayed discounting task, in which they chose between a small, immediate reward or a large, delayed reward [77]. In total, 20 Hz electrical stimulation was applied “on-demand”, triggered by an animal selecting the large, delayed reward outcome. Stimulation biased behavior toward the large reward choice, despite the temporal delay (2–10 s). Multiple cortico-striatal targets (including OFC) had this effect [77]. Although these results are limited due to lack of control/replication, they warrant testing beta-frequency modulation in an injured cohort.
While much of the current evidence regarding beta oscillations observed after TBI stems from our research, other studies have identified beta activity for reward processing and cognitive control across both humans and animal models [71,72,73,78,79]. Beta-evoked oscillations are seen during decision-making, particularly in situations with high cognitive demand [73]. In humans with mild TBI, deficits in frontal beta power (measured with magnetoencephalography) reflect thalamo-cortical network damage [5]. Beta-functional connectivity, compared with other frequencies, best predicted symptoms severity and mild TBI classification [5].
Contrary to these findings, some researchers have found evoked beta power following the omission of expected rewards [84]. Thus, although beta likely plays a role in reward processing, under what conditions it signals expectancy and reward outcome remain unclear. However, the role of beta oscillations following TBI warrants further investigation.
3.3. Other Potential Neurophysiological Biomarkers
Beta oscillations in the cortico-striatal network may represent a promising neurophysiological biomarker tied to a specific cognitive domain (reward valuation). Other neurophysiological markers pertaining to different cognitive domains may also be used to improve TBI outcomes with neuromodulation. For example, theta oscillations are suppressed after TBI and return over the course of recovery [30,83]. Theta oscillations are an appealing target to consider, as they are believed to modulate long-term potentiation underlying learning and memory [85,86]. Stimulating the medial septum (increasing theta oscillations) restored cognitive performance in rats with TBI [83,86,87]. Likewise, in rodent TBI models, gamma activity is decreased near the focal injury site and signal (and associated behavior consequences) is rescued with 40 Hz modulation (Blue LED photobiomodulation therapy) [88].
Another avenue that has been explored is modulating delta oscillations to change sleep dynamics (and thereby wakeful neural function) that are interrupted by TBI. Delta oscillations typically present in deep sleep are elevated during wakefulness after TBI [31]. Hypothalamic stimulation recovers delta oscillations to restore organized sleep after TBI [31].
4. Limitations and Challenges (In Translation and Beyond)
Despite promising preliminary findings, research on beta oscillations as a biomarker in TBI is still in its infancy. Generally, a role for beta oscillations in facilitating cognition (and reward evaluation) is accepted, but the disruption of beta signals in the case of TBI is less known. Beta oscillations are believed to facilitate top–down control of cognitive behaviors [89], and although these coincide with TBI sequalae, the involvement of beta oscillations explicitly still needs to be explored. Studies examining neurophysiological markers pertaining to cognitive deficits in TBI are sparse and often limited by methodological variability (injury models, severity, timeline, region of interest) across studies. Compared with deficits in motor function, cognitive networks are harder to precisely target and are often confounded by emotional/neuropsychiatric symptoms [90]. More preclinical research is needed to delineate the neurobiological substrates of cognitive behavior. Neurophysiological measures sometimes show weak or no associations with behavioral or functional outcomes. Barone et al., 2024, find that EEG alterations in mild TBI did not significantly correlate with clinical outcomes measures, underscoring the need to further refine which markers can best predict patient outcomes [27]. Although EEG is used clinically in an intensive-care setting to monitor seizure activity or drug effects, it is not common in outpatient settings [90]. Clinical assessment of EEG activity over a more chronic time course will help identify electrophysiological signatures related to individual pathologies.
Moreover, the success of neuromodulation to ameliorate cognitive symptoms of TBI is occluded by inconsistent protocols and translational barriers. Animal stimulation protocols are inconsistent in how they apply neuromodulation to TBI. Methodological factors like the type of stimulation (deep brain v. external), location, duration, and onset of stimulation relevant to TBI must be considered [44]. Further, factors like the use of anesthesia complicate methodological similarities and obscure translatability. While rodent models offer a high degree of experimental control, their translational value is often obscured by differences in anatomical division, cytoarchitecture, network structure, and behavior complexity. Moreover, clinical safety profiles and ideal stimulation parameters may be hard to gleam from rodent studies. The risk of seizures and adverse side effects with TMS or tDCS in TBI patients must be considered when generalizing across species [39,44,45]. Since patients with TBI present a high-risk population, the risk profile and safety guidelines of clinical neuromodulation must be carefully considered. Future work will need to identify homologous neural circuits, validate neurophysiological markers across species, and refine neuromodulation parameters for clinical application.
Finally, due to the heterogenous nature of TBI, multimodal treatments will likely be effective. Guided by biomarkers, stimulation may be paired with therapy or pharmacological treatment to address multiple symptoms. In humans, neuromodulation (tDCS) paired with cognitive training can enhance its success [90]. In some cases, enhancements in cognition were small and non-significant, further supporting the need to identify the networks to apply neuromodulation that will have maximal benefit when paired with cognitive rehabilitation [91]. Multimodal treatments create a challenge in ascertaining the exact mechanism, pathological target, and duration/frequency of treatment needed [44,90]. Without knowledge about which networks to target, it is possible that each treatment modality would target opposing networks and produce a null effect [90].
5. Conclusions
Even after identifying a reliable biomarker for intervention, important questions, like when in the disease progression to intervein and how long benefits persist, will need to be researched. Electrophysiology allows us to measure impairments which are undetectable by other methods, but using multi-modal diagnostic biomarkers (blood, structural, physiological, genetic) offers the most precision to treat individual pathologies by capturing the heterogeneous nature of TBI. The current lack of objective biomarkers for TBI poses a significant challenge for translational research. Future work should employ biomarkers to confirm that injuries induced in preclinical models are comparable to humans and that a proposed treatment would be clinically effective.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Wilde E.A. Wanner I.-B. Kenney K. Gill J. Stone J.R. Disner S. Schnakers C. Meyer R. Prager E.M. Haas M. A Framework to Advance Biomarker Development in the Diagnosis, Outcome Prediction, and Treatment of Traumatic Brain Injury J. Neurotrauma 20223943645710.1089/neu.2021.009935057637 PMC 8978568 · doi ↗ · pubmed ↗
- 2Ghaith H.S. Nawar A.A. Gabra M.D. Abdelrahman M.E. Nafady M.H. Bahbah E.I. Ebada M.A. Ashraf G.M. Negida A. Barreto G.E. A Literature Review of Traumatic Brain Injury Biomarkers Mol. Neurobiol.2022594141415810.1007/s 12035-022-02822-635499796 PMC 9167167 · doi ↗ · pubmed ↗
- 3Gutierre M.U. Telles J.P.M. Welling L.C. Rabelo N.N. Teixeira M.J. Figueiredo E.G. Biomarkers for Traumatic Brain Injury: A Short Review Neurosurg. Rev.2021442091209710.1007/s 10143-020-01421-033078327 · doi ↗ · pubmed ↗
- 4Bigler E.D. Neuroimaging Biomarkers in Mild Traumatic Brain Injury (m TBI)Neuropsychol. Rev.20132316920910.1007/s 11065-013-9237-223974873 · doi ↗ · pubmed ↗
- 5Zhang J. Safar K. Emami Z. Ibrahim G.M. Scratch S.E. Da Costa L. Dunkley B.T. Local and Large-Scale Beta Oscillatory Dysfunction in Males with Mild Traumatic Brain Injury J. Neurophysiol.20201241948195810.1152/jn.00333.202033052746 · doi ↗ · pubmed ↗
- 6Freire M.A.M. Rocha G.S. Bittencourt L.O. Falcao D. Lima R.R. Cavalcanti J.R.L.P. Cellular and Molecular Pathophysiology of Traumatic Brain Injury: What Have We Learned So Far?Biology 202312113910.3390/biology 1208113937627023 PMC 10452099 · doi ↗ · pubmed ↗
- 7Ladak A.A. Enam S.A. Ibrahim M.T. A Review of the Molecular Mechanisms of Traumatic Brain Injury World Neurosurg.201913112613210.1016/j.wneu.2019.07.03931301445 · doi ↗ · pubmed ↗
- 8Risbrough V.B. Vaughn M.N. Friend S.F. Role of Inflammation in Traumatic Brain Injury–Associated Risk for Neuropsychiatric Disorders: State of the Evidence and Where Do We Go From Here Biol. Psychiatry 20229143844810.1016/j.biopsych.2021.11.01234955170 PMC 12128916 · doi ↗ · pubmed ↗