Chemical Composition, Antioxidant, and Enzyme Inhibitory Activities of Artemisia schmidtiana Maxim. Essential Oil
Xinyu Zhu, Xu Liu

TL;DR
This study analyzes the chemical makeup and health benefits of Artemisia schmidtiana essential oil, showing it has antioxidant properties and can inhibit certain enzymes.
Contribution
The study identifies key compounds in Artemisia schmidtiana essential oil and evaluates its antioxidant and enzyme inhibitory effects for potential pharmaceutical use.
Findings
The essential oil contains germacrene D, falcarinol, and β-caryophyllene as major constituents.
The oil showed significant inhibition of α-glucosidase and β-lactamase enzymes.
Molecular docking confirmed interactions between the oil's compounds and target enzymes.
Abstract
Artemisia schmidtiana Maxim., a plant belonging to the Asteraceae family, is renowned for its extensive ethnomedicinal applications and distinctive aromatic qualities. This study evaluated the chemical composition, antioxidant capacity, and inhibitory effects on acetylcholinesterase (AChE), α-glucosidase, and β-lactamase of its essential oil (EO). The major constituents of the EO were identified as germacrene D (16.29%), falcarinol (11.02%), β-caryophyllene (9.43%), α-zingiberene (7.93%), phytol (6.06%), and α-humulene (4.04%). The EO demonstrated radical scavenging activity against DPPH (44.9% at 5 mg/mL) and ABTS (IC50 = 0.72 ± 0.02 mg/mL) radicals, with a FRAP antioxidant capacity of 126.61 ± 0.59 μmol·g−1. Additionally, the EO exhibited modest AChE inhibition (16.7% at 250 μg/mL) and significant inhibition of α-glucosidase and β-lactamase, with IC50 values of 178.80 ± 17.02 μg/mL…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
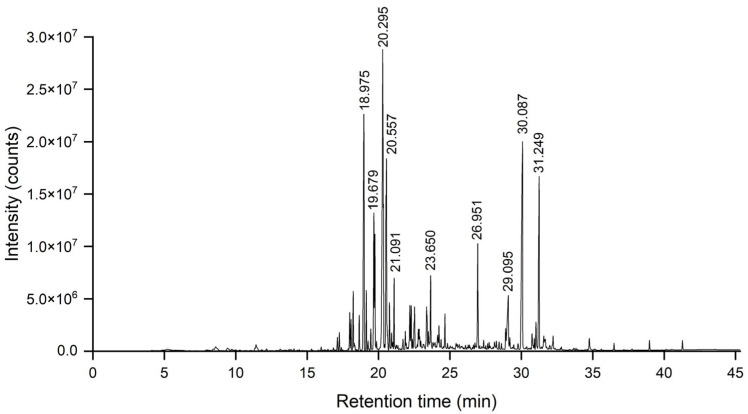 Figure 1
Figure 1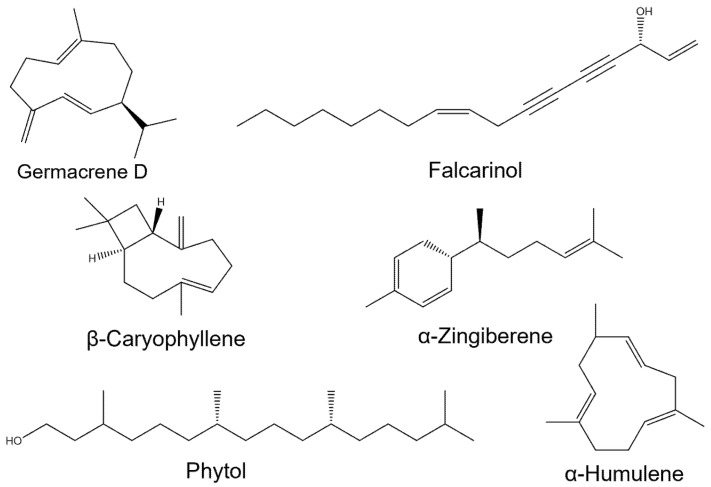 Figure 2
Figure 2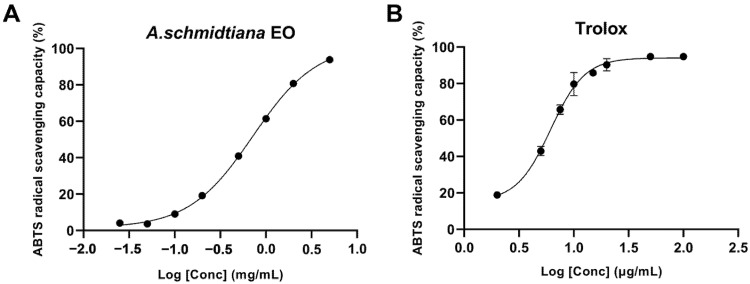 Figure 3
Figure 3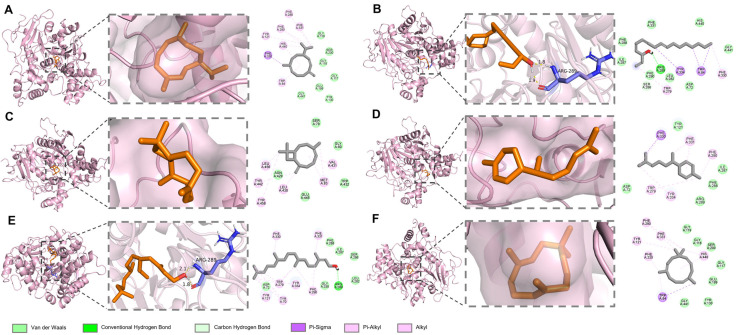 Figure 4
Figure 4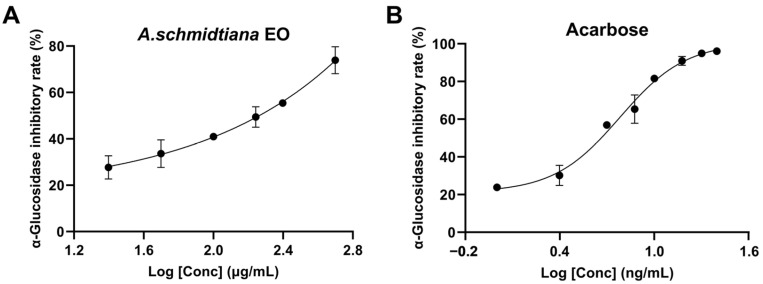 Figure 5
Figure 5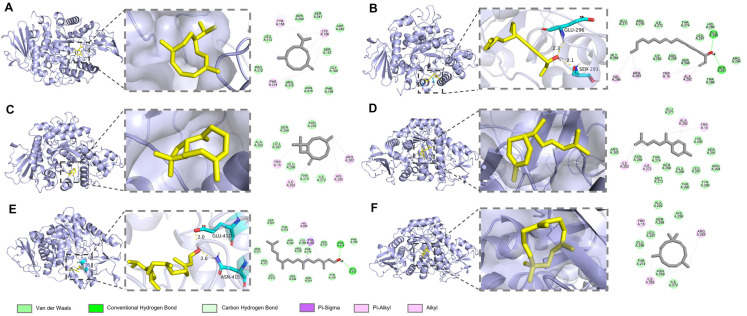 Figure 6
Figure 6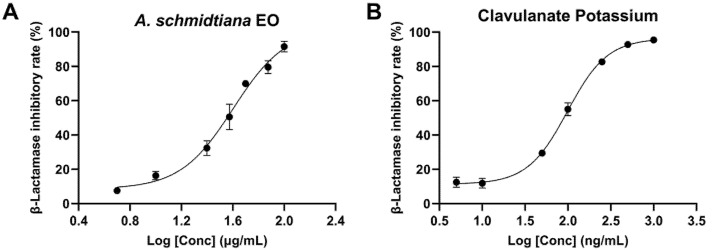 Figure 7
Figure 7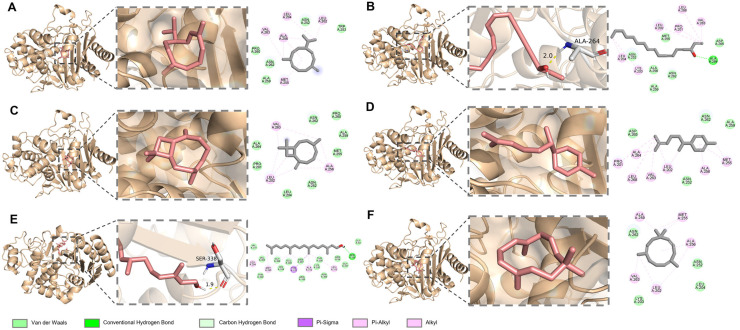 Figure 8
Figure 8Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsEssential Oils and Antimicrobial Activity · Cholinesterase and Neurodegenerative Diseases · Insect Pest Control Strategies
1. Introduction
Plants represent one of the primary sources of raw materials for the food, pharmaceutical, and cosmetic industries, with many nutraceuticals and cosmeceuticals deriving their medicinal value from plant-based constituents [1]. Among various plant-derived products, essential oil (EO), a complex mixture of hydrophobic volatile compounds originating from plant secondary metabolism, is a promising molecular library for drug discovery [2]. This oil exhibits specific bioactivities, including antimicrobial, anti-inflammatory, and antioxidant properties [3,4]. Furthermore, the emergence of green consumerism has increased consumer demand for natural-extracted pharmaceutical agents with minimal environmental footprints. As natural products, EO has been widely utilized in traditional medicine to promote human health and treat diseases, owing to its intriguing biochemical profiles and high added value in environmental sustainability [5].
In exploring the bioactivity of EO, its antioxidant capacity is often regarded as paramount, given the central role of oxidative stress in various pathological processes [6]. Oxidative stress arises from either excessive production of free radicals or compromised antioxidant defense systems [7,8]. Free radicals, characterized by unpaired electrons, are highly reactive and readily interact with cellular components [9]. While they participate in critical cellular processes, an overabundance of free radicals can inflict widespread damage on cellular structures [1]. This damage is implicated in the pathogenesis of numerous chronic and degenerative diseases, including Alzheimer’s disease (AD), diabetes mellitus (DM), and cardiovascular disorders [10]. Nevertheless, this detrimental process can be modulated through exogenous antioxidants derived from natural sources, such as plant EO and extracts, which offer a promising therapeutic avenue [11].
AD is an age-related neurodegenerative disorder characterized by three hallmark features: the deposition of amyloid-β (Aβ) plaques, hyperphosphorylation of the cytoskeletal protein tau, and reduced levels of acetylcholine (ACh), leading to neuronal dysfunction [12]. Among these, Aβ accumulation plays a pivotal role in driving oxidative stress [13]. Concurrently, AD patients exhibit decreased levels of antioxidants, such as uric acid, vitamin C, and vitamin E, and reduced activity of antioxidant enzymes like superoxide dismutase and catalase [14]. These findings underscore oxidative stress as a key contributor to AD pathogenesis, suggesting that antioxidant supplementation may offer therapeutic benefits [15]. Furthermore, acetylcholinesterase (AChE), a serine protease that hydrolyzes ACh in the brain, contributes to AD progression by diminishing ACh levels and disrupting neuronal signaling [16]. Notably, EO can cross the blood–brain barrier and modulate the central nervous system, potentially alleviating AD symptoms due to its small molecular size and lipophilic properties [17]. Therefore, EO is a promising candidate for developing novel therapeutics targeting neurodegenerative diseases.
DM is a chronic disease characterized by elevated blood glucose levels due to insulin deficiency or impaired insulin function [18]. Oxidative stress is widely recognized as a pivotal contributor to the pathogenesis of DM, primarily through mechanisms such as mitochondrial H_2_O_2_ production and NADPH oxidase activation, which disrupt insulin signaling and promote insulin resistance [19]. Consequently, antioxidants demonstrate beneficial effects in DM treatment. In addition, the inhibition of α-glucosidase, a carbohydrate-hydrolyzing enzyme located on the intestinal brush border, represents a critical therapeutic strategy. By delaying the digestion of oligosaccharides into absorbable monosaccharides, α-glucosidase inhibitors effectively modulate postprandial glucose absorption, thereby mitigating glycemic fluctuations and slowing disease progression [20]. Recent studies have identified EO derived from 20 plant species with significant potential for DM management, attributed to their dual antioxidant and α-glucosidase inhibitory activities [21]. EO has already been utilized in therapeutic formulations and has demonstrated a favorable safety profile with no significant adverse effects reported [22]. Consequently, EO may offer a promising avenue for discovering and developing novel antidiabetic agents.
β-Lactam antibiotics are widely regarded as one of the most effective antibacterial agents due to their broad spectrum of activity, favorable pharmacokinetics, and safety profile [23]. However, the extensive use of β-lactams has led to the emergence and dissemination of resistance, mediated through diverse mechanisms, including target modification, downregulation of porins required for β-lactam entry, overexpression of efflux systems, and enzymatic modification or degradation [24]. Among these, enzyme-mediated resistance, driven by β-lactamase activity, poses a significant threat. β-Lactamases, produced by both Gram-positive and Gram-negative bacteria, hydrolyze β-lactam antibiotics, rendering one of the most significant threats to antibacterial efficacy [24,25]. In our previous work, Spermacoce alata EO exhibited potent β-lactamase inhibitory activity [26]. Therefore, developing EO-based adjunctive therapies may represent a promising strategy to counteract β-lactam resistance and enhance the efficacy of existing antibiotics [27].
Artemisia, one of the largest and most widely distributed genera within the Asteraceae family, comprises over 500 species predominantly found in temperate regions of Europe, Asia, and North America [28]. Artemisia species are renowned for their diverse bioactivities, including antimicrobial, anticancer, anti-inflammatory, antioxidant, and antipyretic properties, making them highly valued in ethnopharmacology [29,30]. The 2015 Nobel Prize in Medicine, awarded for the discovery of artemisinin, a potent antimalarial sesquiterpene lactone isolated from Artemisia annua, renewed scientific focus on these plants, including Artemisia schmidtiana Maxim. [30]. A. schmidtiana, a perennial herbaceous plant native to alpine or rocky environments, is characterized by its slender, silver–white hairy stems and leaves, reaching heights of approximately 10 cm, and produces small white flowers in July and August. It has historically been prized for its medicinal properties and distinctive aromatic qualities, and it is commonly cultivated as an ornamental plant in gardens and pots, favored by horticultural enthusiasts for its unique foliage. Beyond its traditional uses, thiophene acetylenes were isolated from A. schmidtiana in 1986 [31]. Thiophenes, a class of secondary metabolites, exhibit a broad spectrum of biological activities, including antimicrobial, antiviral, HIV-1 protease inhibitory, and anticancer effects, highlighting the plant’s potential for further pharmaceutical development [32].
The previous literature has documented the broad bioactivities and high medicinal value of A. schmidtiana. However, the chemical composition and biological activities of its EO remain unexplored. To address this gap, we investigated the chemical profile, antioxidant activity, and inhibitory effects of A. schmidtiana EO on AChE, α-glucosidase, and β-lactamase. Additionally, molecular docking was employed to elucidate the potential mechanisms underlying its enzyme-inhibitory properties.
2. Materials and Methods
2.1. Plant Material
Plant samples of aerial parts at the vegetative stage were collected from a commercial nursery in Shanghai City, China (31.2304° N, 121.4737° E) in September 2023. The species was identified as A. schmidtiana by Pro. Zhao Hong, through morphological characteristics. The voucher specimen is stored at the Center for Bioscience Analysis and Testing, Shandong University, Weihai, China, with registration number EO2325. The plant samples were refrigerated at −18 °C until EO extraction.
2.2. EO Extraction
Fresh leaves and stems (140 g) of the plant material were finely crushed and placed into a 5 L round-bottom flask, and then 3 L of ultrapure water. Hydrodistillation was carried out for approximately 4 h using a Clevenger-type apparatus to isolate the EO from the plant material. The EO was then separated from the aqueous layer using diethyl ether. Subsequently, the extracted EO was dried using sodium sulfate and concentrated using a Termovap sample concentrator (MD200-1, Shanghai Huyi Technology Co., Ltd., Shanghai, China). The obtained EO was stored at a low temperature (4 °C) for further analysis.
2.3. GC-MS and GC-FID Analyses
The composition and relative content of the EO were analyzed using gas chromatography–mass spectrometry (GC-MS) and gas chromatography with flame ionization detection (GC-FID). GC-MS analysis was conducted on an Agilent 7890–5975C system (Santa Clara, CA, USA) with an HP-5MS fused silica capillary column (30 m × 0.25 mm, 0.25 μm film thickness, Agilent, Santa Clara, CA, USA). The injector and interface temperatures were maintained at 260 °C and 280 °C, respectively. The oven temperature program began at 50 °C for 4 min, followed by a gradient increase to 280 °C at 6 °C/min, and held for 3 min. Ultra-pure helium (99.999%) served as the carrier gas at a 1.0 mL/min flow rate. The mass spectrometer operated in electron ionization (EI) mode at 70 eV, with a scan range of 25–500 amu and a quadrupole temperature of 150 °C. Samples were prepared as a 1% (w/v) solution in dichloromethane, and 0.3 µL was injected in splitless mode. A PerkinElmer Clarus 500 system (Shelton, CT, USA) with an HP-5 column (30 m × 0.25 mm, 0.25 μm film thickness, Agilent, Santa Clara, CA, USA) was used for GC-FID analysis. The injector and detector temperatures were set at 260 °C and 305 °C, respectively. The oven temperature followed the same program as GC-MS, with nitrogen as the carrier gas at 1.1 mL/min.
Compound identification was achieved by comparing mass spectra and retention indices (RI) with the NIST20 library and published data. RIs were calculated using GC-MS data from a series of n-alkanes (C_8_–C_30_) analyzed under identical conditions.
2.4. Antioxidant Activities Evaluation
2.4.1. DPPH Method
The experimental protocol was slightly modified from previously established methods [33]. 6-Hydroxy-2,5,7,8-tetramethylchroman-2-carboxylic acid (Trolox) was used as the positive control. Trolox and the EO stock solution concentrations were prepared in ethanol at 0.5 mg/mL and 50 mg/mL, respectively. Aliquots of 50 μL from either the Trolox or gradient-diluted EO solutions were transferred into a 96-well microplate, followed by 200 μL of 0.17 mmol/L 2,2-diphenyl-1-picrylhydrazyl (DPPH) solution. The microplate was shielded from light and incubated at 25 °C for 30 min. Absorbance was subsequently measured at 516 nm using an Epoch microplate spectrophotometer (BioTek Instruments, Minneapolis, MN, USA). The DPPH radical scavenging capacity (RSC%) was determined using Equation (1):
where A_Sample_ is the absorbance of the tested sample at different concentrations, A_Control_ is the absorbance of the control (ethanolic DPPH solution), and A_Sample Blank_ is the absorbance of the ethanol sample without DPPH.
2.4.2. ABTS Method
The experimental protocol was modified slightly based on established methodologies from prior studies [26]. The ABTS^•+^ radical solution was prepared by combining 7.4 mmol/L 2,2-azino-bis(3-ethylbenzothiazoline-6-sulfonic acid) (ABTS) with 2.6 mmol/L potassium persulfate. The mixture was incubated in the dark at room temperature for 12 h to ensure complete radical generation. Subsequently, 200 μL of the diluted ABTS^•+^ solution was mixed with 50 μL of gradient-diluted ethanolic EO solutions in a 96-well plate. Absorbance was measured at 734 nm six minutes after initiating the reaction. Ethanol served as the blank and dilution solvent, while 0.5 mg/mL Trolox was used as the positive control. The ABTS radical scavenging capacity (RSC%) was calculated to evaluate antioxidant activity using Equation (2):
where A_0_ and A are the absorbance of 200 μL diluted ABTS^•+^ solution mixed with 50 μL ethanol and 50 μL sample solution, respectively, at 734 nm. IC_50_ was then calculated.
2.4.3. Ferric-Reducing Antioxidant Power (FRAP) Method
The assay was performed following established protocols with minor adaptations. Three stock solutions—(i) acetate buffer (pH 3.6), (ii) 10 mM 2,4,6-tripyridyl-s-triazine (TPTZ), and (iii) 20 mM Fe^3+^ solution—were mixed in a 10:1:1 ratio and diluted 50-fold with ethanol to prepare the FRAP working solution. A standard curve was constructed using Trolox. EO samples were serially diluted to 5000, 2500, 1000, 500, 250, 100, 50, and 25 μg/mL concentrations. The EO samples were combined with the FRAP working solution and incubated at 37 °C in the dark. Absorbance was measured at 593 nm, and the antioxidant capacity was expressed as Trolox equivalent antioxidant concentration (TEAC) by referencing the standard curve. All experiments were conducted in triplicate, and results were reported as mean values.
2.5. Anti-AChE Activity Test
This assay was conducted using the previously described method with minor modifications [34]. In total, 145 μL of 0.1 mM phosphate buffered saline (PBS) (pH 8.0), 20 μL of ethanolic EO solution, and 15 μL of AChE solution (0.28 U/mL) were combined in a microplate, with galantamine as the positive control. The mixture was incubated at 4.0 °C for 20 min. Subsequently, 10 μL of 15 mM acetylthiocholine iodide (ATCI) and 10 μL of 2 mM 5,5′-dithiobis-(2-nitrobenzoic acid) (DTNB) were added, followed by homogenization for 60 s. Absorbance was measured at 412 nm at 60 s intervals for 6 min using a microplate reader (Epoch, BioTek Instruments, Minneapolis, MN, USA). The acetylcholinesterase inhibition rate was calculated based on the absorbance data. The AChE inhibition rate is calculated according to Formula (3):
where K_E_ is the initial reaction rate of the enzyme without the inhibition, while K_S_ is the initial reaction rate of the inhibited enzyme.
2.6. Anti-α-Glucosidase Capacity Test
The test was conducted according to previous research with minor modifications [7]. In total, 20 μL of ethanolic EO solution, 80 μL of 100 mM PBS (pH 6.8), and 40 μL of α-glucosidase solution (0.25 U/mL) were combined in a microplate, with acarbose serving as the control. The mixture was incubated at 30 °C for 10 min. Subsequently, 20 μL of 3.0 mg/mL 4-nitrophenyl-β-D-glucopyranoside (pNPG) was added, followed by homogenization for 60 s and further incubation at 30 °C for 4 min. Absorbance was measured at 410 nm at 60 s intervals for 6 min. The α-glucosidase inhibition rate is expressed using Equation (3). The IC_50_ value was calculated using nonlinear regression.
2.7. Test for β-Lactamase Inhibitory Effect
The β-lactamase inhibitory effect was assessed using a previously described method with some modifications [35]. In total, 20 μL of ethanolic EO solution, 30 μL of 50 mM PBS (pH 7.0), and 100 μL of β-lactamase solution (1000 U/mL) were combined in a microplate, with clavulanate potassium serving as the positive control. The mixture was incubated at 30 °C for 10 min. Subsequently, 50 μL of 0.1 mg/mL nitrocefin was added, followed by an additional 10 min incubation at 30 °C. Absorbance was measured at 489 nm. The β-lactamase inhibition rate was calculated using the following Formula (4):
where A_S_ is the absorbance of the EO-containing sample, A_sb_ is the absorbance of the sample blank reaction, A_E_ is the absorbance of the reaction in which the enzyme was not inhibited, and A_B_ is the absorbance of the blank response. IC_50_ was assessed using nonlinear regression.
2.8. Molecular Docking
In the molecular docking experiments, complexes of AChE (Tetronarce californica, PDB code: 1EA5), α-glucosidase (Saccharomyces cerevisiae, PDB code: 3AJ7), and β-lactamase (Enterobacter cloacae, PDB code: 7TI1) were collected from the PDB database. The receptor structures were prepared by removing bound water molecules and ligands using PyMol v2.2.0, and hydrogen atoms were subsequently added to ensure proper protonation states. The 3D structures of the major components of A. schmidtiana EO, used as ligands for interaction with the enzyme receptors, were retrieved from the CAS SciFinder Discovery Platform (https://scifinder-n.cas.org/, accessed on 10 March 2025) with energy minimized using Chem3D.
Autodock v4.2.6 was employed to perform semi-flexible molecular docking. Each ligand was subjected to at least 50 runs. Results were analyzed using the Lamarckian genetic algorithm. Binding energies and interactions between the ligands and proteins were evaluated during the docking process. Discovery Studio visualizer and PyMol v2.2.0 were used to visualize the docking results.
3. Results and Discussion
3.1. EO Yield and Chemical Composition Analysis
The A. schmidtiana EO, obtained through hydrodistillation of 0.14 kg of plant material with 3.0 L ultrapure water, yielded 0.20 mL of red, oily liquid, corresponding to an EO yield of 0.14% (v/w). The production of plant EO is influenced by numerous factors, including genetic variability, geographical distribution, climatic conditions, seasonal variations, and post-harvest processing, such as drying and storage [36]. The EO yield of A. schmidtiana (0.14%) was comparable to other Artemisia species (0.26–0.31%) [37,38,39], indicating normal production levels for this genus. The total ion chromatogram (TIC) of A. schmidtiana is shown in Figure 1.
EO is a complex mixture of volatile molecules, typically composed of multiple compounds in varying proportions, each exhibiting distinct chemical structures and functionalities. According to elution order, retention time (RT), and RI, each EO component was identified, as listed in Table 1. A total of 80 compounds, amounting to 98.47%, were identified. The major constituents included germacrene D (16.29%), falcarinol (11.02%), β-caryophyllene (9.43%), α-zingiberene (7.93%), phytol (6.06%), α-humulene (4.04%), n-hexadecanoic acid (3.38%), hexahydrofarnesyl acetone (3.31%), sesquisabinene (3.29%), and neointermedeol (2.08%). Further classification revealed a sesquiterpenoid-dominated profile (65.87%), followed by aliphatic compounds (18.97%), diterpenoids (6.21%), and monoterpenoids (0.73%) (Figure 2).
Germacrene D (16.29%), the most abundant compound in the EO, primarily exists as (+) and/or (−) enantiomers in EO and is recognized as a significant aromatic compound in the fragrance industry, serving as an antibiotic, repellent, attractant, or pheromone [40,41]. The second most abundant compound, falcarinol (11.02%), has been shown to reduce the expression of the apoptosis marker caspase-3, decrease basal DNA strand breaks, and promote CaCo-2 cell proliferation, thereby preventing and ameliorating colon cancer progression [42]. Falcarinol also exhibits cytotoxicity against human tumor cell lines and exerts kinetic effects on the proliferation of primary mammalian cells [43]. Another notable compound, β-caryophyllene (9.43%), has also garnered attention as a potential broad-spectrum pharmacological agent [44]. β-Caryophyllene extracted from Aquilaria crassna EO has been demonstrated to possess remarkable anticancer, antioxidant, and antimicrobial properties [45]. While these major constituents play pivotal roles in the biological activities of A. schmidtiana EO, minor components also contribute synergistically or antagonistically, collectively defining the bioactivity of the EO system. Given the chemical diversity of the EO system, we further evaluated its multifaceted biological activities, including antioxidant activity and inhibition of key therapeutic target enzymes associated with diseases.
3.2. Antioxidant Activity Analysis of A. schmidtiana EO
Due to the variability among antioxidant testing systems, results from a single assay cannot fully capture the antioxidant potential of samples. The chemical complexity of EO, comprising diverse constituents with unique behaviors, polarities, and functional groups, often leads to inconsistent outcomes depending on the assay used [46]. Therefore, a multi-assay approach is recommended for accurate evaluation.
The experimental results of DPPH radical scavenging activity are presented in Table 2. At the maximum tested concentration of 5.0 mg/mL, the inhibition rate was 44.9%, while the control Trolox achieved nearly 100% inhibition at the same concentration. Figure 3 shows the S-shaped curve of ABTS radical scavenging activity, which increases with concentration. From the graph, the IC_50_ of the EO was determined to be 0.72 ± 0.02 mg/mL. The EO generally demonstrated potent ABTS radical scavenging activity, significantly outperforming its DPPH radical scavenging capacity.
Previous studies have reported better coefficient of determination (R^2^) and coefficient of variation (CV) values for ABTS assays than DPPH assays, potentially offering a plausible explanation for our results. These differences may arise from the steric hindrance caused by the three phenyl groups surrounding the nitrogen atom in the DPPH radical, which impedes hydrogen atom transfer, or the higher sensitivity of DPPH radicals to reaction conditions, which contributes to the greater accuracy of ABTS assays [47]. Additionally, the stereoselectivity of active compounds and their solubility in various assay systems are considered plausible factors influencing the radical scavenging capacity of EO [48]. Typically, the major terpene compounds in A. schmidtiana EO lack polar groups, making it challenging to provide hydrogen atoms and limiting their solubility in DPPH assay media. In contrast, ABTS and FRAP assays rely on electron transfer, making them more effective in detecting the antioxidant capacity of both hydrophilic and lipophilic compounds [49]. Therefore, we further conducted FRAP assays to assess antioxidant properties comprehensively and corroborate the above findings.
Unlike free radical scavenging assays, the FRAP test measures the total reducing capacity of A. schmidtiana EO. Iron salts serve as the basis of the FRAP assay, acting as oxidants during electron transfer processes. The reducing capacity of the EO may serve as a reliable predictor of its potential antioxidant activity [50]. In this study, the EO exhibited a FRAP value of 126.61 ± 0.59 μmol/g.
By examining the structures of the major compounds in A. schmidtiana EO (Figure 2), it is evident that germacrene D (16.29%), falcarinol (11.02%), and α-zingiberene (7.93%) possess extensive conjugated systems, a common feature among most antioxidants. High conjugation allows electron-donating groups (EDGs) to delocalize additional electrons across overlapping π-bonds, enhancing the ability to quench free radicals [51]. Consequently, stronger conjugation correlates with greater antioxidant capacity. In addition to these major compounds, minor constituents like fokienol (1.46%) and isogermacrene D (0.51%) exhibit significant conjugation, contributing substantially to the EO’s antioxidant activity. Therefore, we hypothesize that the robust antioxidant activity of A. schmidtiana EO primarily stems from its abundance of conjugated terpenoids.
3.3. Anti-AChE Activity of A. schmidtiana EO
AD, the most prevalent form of dementia, is characterized by oxidative stress and progressive neuronal degeneration, often accompanied by visible Aβ deposits in the brain and reduced acetylcholine levels [52]. A typical therapeutic approach for AD management involves the use of AChE inhibitors, such as galantamine [53]. In this study, A. schmidtiana EO demonstrated an AChE inhibition rate of 16.9% at the highest tested concentration of 250 μg/mL. In contrast, the positive control galantamine achieved nearly 100% inhibition at the same concentration (Table 3).
Molecular docking has become indispensable in recent research for demonstrating the binding affinity of bioactive molecules to target enzymes and validating their potential inhibitory effects. It is reported that small molecules and proteins may exhibit bioactivity when their binding affinity is less than −5.0 kcal/mol, and stable binding occurs when the affinity is below −7.0 kcal/mol [54]. All major compounds docked with AChE exhibited binding energies below −5 kcal/mol, indicating favorable interactions (Table 4). Among these, β-caryophyllene (−7.24 kcal/mol) showed the lowest binding energy, followed by α-humulene (−7.23 kcal/mol) and germacrene D (−7.12 kcal/mol). Notably, these sesquiterpenes lack polar groups capable of forming hydrogen bonds with AChE, suggesting their superior binding may be attributed to significant hydrophobic interactions. Hydrogen bonds are crucial for the stability of protein–ligand complexes, enhancing the stability and longevity of molecules docked in the enzyme’s active site [55]. Alcohol compounds, such as falcarinol and phytol, formed two hydrogen bonds with ARG A: 289, respectively, underscoring their potential inhibitory effects (Figure 4).
Miyazawa et al. revealed that alkenes exhibit more vigorous AChE inhibitory activity than alcohols and ketones. Double bonds in alkenes significantly enhance their inhibitory effect, with allyl-methyl-containing compounds showing potent inhibition. In contrast, oxygen-containing functional groups tend to reduce AChE inhibition [56]. In vivo studies further demonstrated that germacrene D inhibits AChE activity by over 50% in rat brain structures [57]. Moreover, β-caryophyllene has been utilized in treating neurodegenerative diseases [57]. While these compounds demonstrate significant AChE inhibitory activity, the in vitro assay revealed that the EO exhibited only moderate AChE inhibition. This apparent discrepancy may arise from antagonistic interactions among the volatile constituents within the complex EO system, potentially attenuating the collective inhibitory effect [58].
3.4. Anti-α-Glucosidase Activity of A. schmidtiana EO
α-Glucosidase, a critical digestive enzyme, accelerates the breakdown of polysaccharides, mainly starch, into glucose by cleaving (1 → 4) glycosidic bonds, thereby elevating blood glucose levels. Inhibitors of α-glucosidase can effectively mitigate postprandial hyperglycemia, potentially relieving diabetes symptoms [59]. In this study, A. schmidtiana EO demonstrated an IC_50_ value of 178.80 ± 17.02 μg/mL for α-glucosidase inhibition, and acarbose exhibited an IC_50_ value of 6.04 ± 0.39 ng/mL (Figure 5).
To further elucidate the mechanism underlying the anti-α-glucosidase activity of A. schmidtiana EO, molecular docking was employed to evaluate the interactions between main compounds and the catalytic site of α-glucosidase. As shown in Table 5, the binding energies of all compounds ranged from −5.53 kcal/mol to −6.65 kcal/mol, with no unfavorable interactions observed (Table 5). Similar to the results with AChE, sesquiterpenes did not form hydrogen bonds with α-glucosidase. In contrast, falcarinol formed two hydrogen bonds with protein residues GLU A: 296 and SER A: 291, while phytol established hydrogen bonds with ASN A: 415 and GLU A: 411 (Figure 6). These compounds, characterized by multiple rotatable bonds and higher polarity, demonstrated potential as α-glucosidase inhibitors. β-Caryophyllene (−6.65 kcal/mol) exhibited the lowest binding energy among all compounds, followed closely by germacrene D (−6.59 kcal/mol), α-humulene (−6.46 kcal/mol), and α-zingiberene (−6.23 kcal/mol), likely due to their analogous cycloalkene structures.
Studies have shown that EO rich in terpenes can inhibit key enzymes associated with diabetes, particularly α-glucosidase. Majouli et al. reported that terpenes administered to alloxan-induced diabetic rats exhibited hypoglycemic effects and high antioxidant activity [60]. Currently, several Artemisia species, including A. anethifolia, A. desertorum, A. latifolia, A. umbrosa, A. tanacetifolia, A. palustris, A. leucophylla, and A. commutata, have been demonstrated to significantly inhibit α-glucosidase [61], highlighting Artemisia species’ potential in diabetes treatment.
3.5. Anti-β-Lactamase Activity of A. schmidtiana EO
Since the introduction of β-lactam antibiotics, bacterial resistance to these drugs has become an increasingly critical issue. The production of β-lactamases by bacteria represents the most common resistance mechanism to β-lactam antibiotics. A key strategy to combat antibiotic resistance involves using β-lactamase inhibitors [62]. In this study, we evaluated the β-lactamase inhibitory activity of A. schmidtiana EO, which exhibited an IC_50_ value of 40.06 ± 8.22 μg/mL. At the same time, the positive control clavulanate potassium demonstrated an IC_50_ of 98.93 ± 3.93 ng/mL (Figure 7).
Molecular docking analysis revealed detailed interactions between six major components of A. schmidtiana EO and the β-lactamase target. The binding energies of these six small molecules with β-lactamase ranged from −5.15 kcal/mol to −6.71 kcal/mol, with no unfavorable interactions observed (Table 6). Falcarinol formed one hydrogen bond with ALA A:264, while phytol established a hydrogen bond with SER A:338 (Figure 8). Although these hydrogen bonds stabilized the binding between the small molecules and the receptor, their relatively high binding energies and inhibition constants suggested limited inhibitory efficacy. Notably, germacrene D (−6.71 kcal/mol) exhibited the lowest binding energy, followed by β-caryophyllene (−6.56 kcal/mol), α-humulene (−6.54 kcal/mol), and α-zingiberene (−6.51 kcal/mol). These compounds primarily interacted with β-lactamase through van der Waals forces, potentially contributing to their inhibitory effects. In addition, minor constituents present in EO may also play a critical role in inhibiting β-lactamase activity.
4. Conclusions
In this study, the chemical composition, antioxidant activity, anti-AChE, anti-α-glucosidase, and anti-β-lactamase activities of A. schmidtiana EO were investigated for the first time. A total of 80 compounds were identified, with the major compounds being germacrene D (16.29%), falcarinol (11.02%), β-caryophyllene (9.43%), α-zingiberene (7.93%), phytol (6.06%), and α-humulene (4.04%). The EO exhibited antioxidant activity and inhibitory effects on AChE, α-glucosidase, and β-lactamase in vitro, suggesting its potential as a source of enzyme inhibitors. Molecular docking studies further supported the in vitro findings. These bioactivities collectively indicate the pharmacological potential of A. schmidtiana EO. However, the in vitro results provide only preliminary data on its biological activities. Future in vivo studies are essential to evaluate potential side effects, therapeutic efficacy, and targeted delivery of the EO. Therefore, integrating in vivo research with pharmacodynamic and pharmacokinetic analyses is crucial for further developing and applying A. schmidtiana EO.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Michalak M. Plant-Derived Antioxidants: Significance in Skin Health and the Ageing Process Int. J. Mol. Sci.20222358510.3390/ijms 2302058535054770 PMC 8776015 · doi ↗ · pubmed ↗
- 2Salazar M.O. Osella M.I. Arcusin D.E.J. Lescano L.E. Furlan R.L.E. New α-glucosidase inhibitors from a chemically engineered essential oil of Origanum vulgare L.Ind. Crops Prod.202015611285510.1016/j.indcrop.2020.112855 · doi ↗
- 3Gupta I. Singh R. Muthusamy S. Sharma M. Grewal K. Singh H.P. Batish D.R. Plant Essential Oils as Biopesticides: Applications, Mechanisms, Innovations, and Constraints Plants 202312291610.3390/plants 1216291637631128 PMC 10458566 · doi ↗ · pubmed ↗
- 4Hou T. Sana S.S. Li H. Xing Y. Nanda A. Netala V.R. Zhang Z. Essential oils and its antibacterial, antifungal and anti-oxidant activity applications: A review Food Biosci.20224710171610.1016/j.fbio.2022.101716 · doi ↗
- 5Omonijo F.A. Ni L. Gong J. Wang Q. Lahaye L. Yang C. Essential oils as alternatives to antibiotics in swine production Anim. Nutr.2018412613610.1016/j.aninu.2017.09.00130140752 PMC 6104524 · doi ↗ · pubmed ↗
- 6Amorati R. Foti M.C. Valgimigli L. Antioxidant Activity of Essential Oils J. Agric. Food Chem.201361108351084710.1021/jf 403496 k 24156356 · doi ↗ · pubmed ↗
- 7Xu Z. Zhu J. Zhao J. Zhu X. Liu X. Essential oil of Hygrophila salicifolia (Vahl) Nees: Chemical composition, evaluation of antioxidant activity, anti-acetylcholinesterase and anti-α-glucosidase activities integrating molecular docking, and network pharmacology analysis Ind. Crops Prod.202422011927810.1016/j.indcrop.2024.119278 · doi ↗
- 8Ancuceanu R. Anghel A.I. HovanețM.V. Ciobanu A.-M. Lascu B.E. Dinu M. Antioxidant Activity of Essential Oils from Pinaceae Species Antioxidants 20241328610.3390/antiox 1303028638539820 PMC 10967544 · doi ↗ · pubmed ↗