Isolation, Identification, Biological Characterization, and Pathogenicity of Entomopathogenic Fungus from the Larvae of the Evergestis extimalis (Scopoli) (Lepidoptera: Pyralidae)
Youhua Ma, Minggang Qin, Yuanfang Zeng, Yinyin Shen, Youpeng Lai, Guangxin Lu

TL;DR
A pathogenic fungus, Mucor hiemalis, was isolated from Evergestis extimalis larvae and shown to effectively control the pest, especially in younger larvae.
Contribution
Identification and characterization of Mucor hiemalis QH01 as a novel biocontrol agent for Evergestis extimalis.
Findings
Mucor hiemalis QH01 was identified as a pathogenic fungus infecting Evergestis extimalis larvae.
The fungus showed higher pathogenicity to younger larvae, with significant survival reduction in second instar larvae.
Optimal growth conditions for M. hiemalis QH01 were determined as SDAY medium, fructose, peptone, 25°C, pH 6.0–7.0, and a 0:24 light:dark cycle.
Abstract
Evergestis extimalis is a significant pest of rapeseed crops. During our study on the biology of E. extimalis, we discovered a strain of pathogenic fungus that infected its larvae. Through morphological and molecular biological analyses, the strain was identified as Mucor hiemalis and named M. hiemalis QH01. Afterwards, the optimal growth conditions for strain QH01 were determined by evaluating mycelial growth rate and sporulation yield as indicators. We evaluated the pathogenicity of M. hiemalis against the larvae of the E. extimalis under laboratory conditions. The results indicated that the strain was highly pathogenic to the younger larvae, followed by the older larvae. Subsequently, the pathogenicity of the strain against the second instar larvae of the E. extimalis was further evaluated under greenhouse conditions. The results showed that the survival rate of the second instar…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
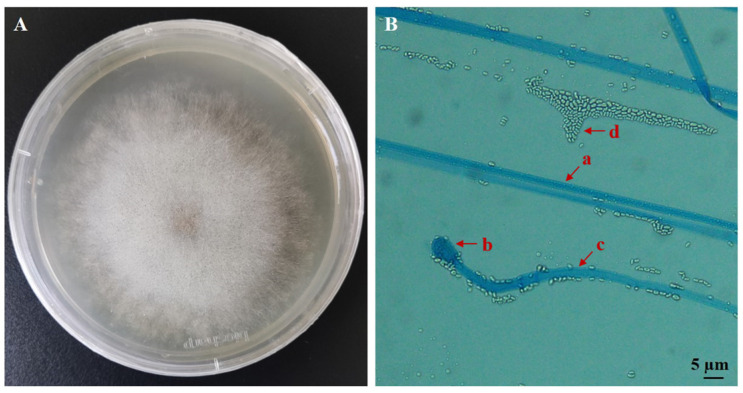 Figure 1
Figure 1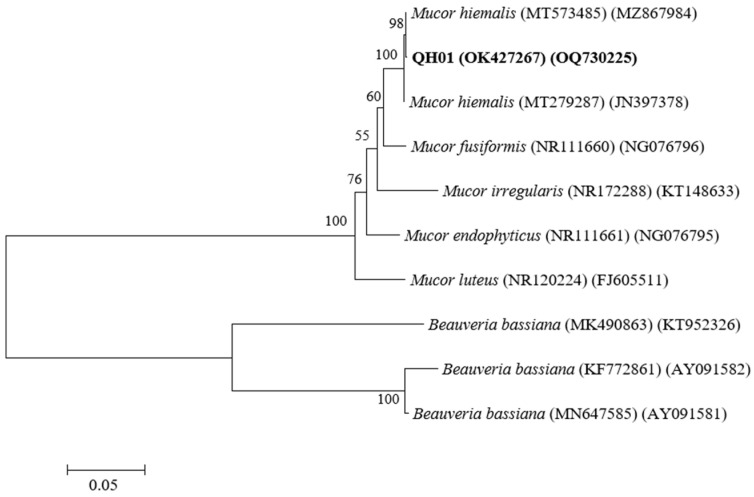 Figure 2
Figure 2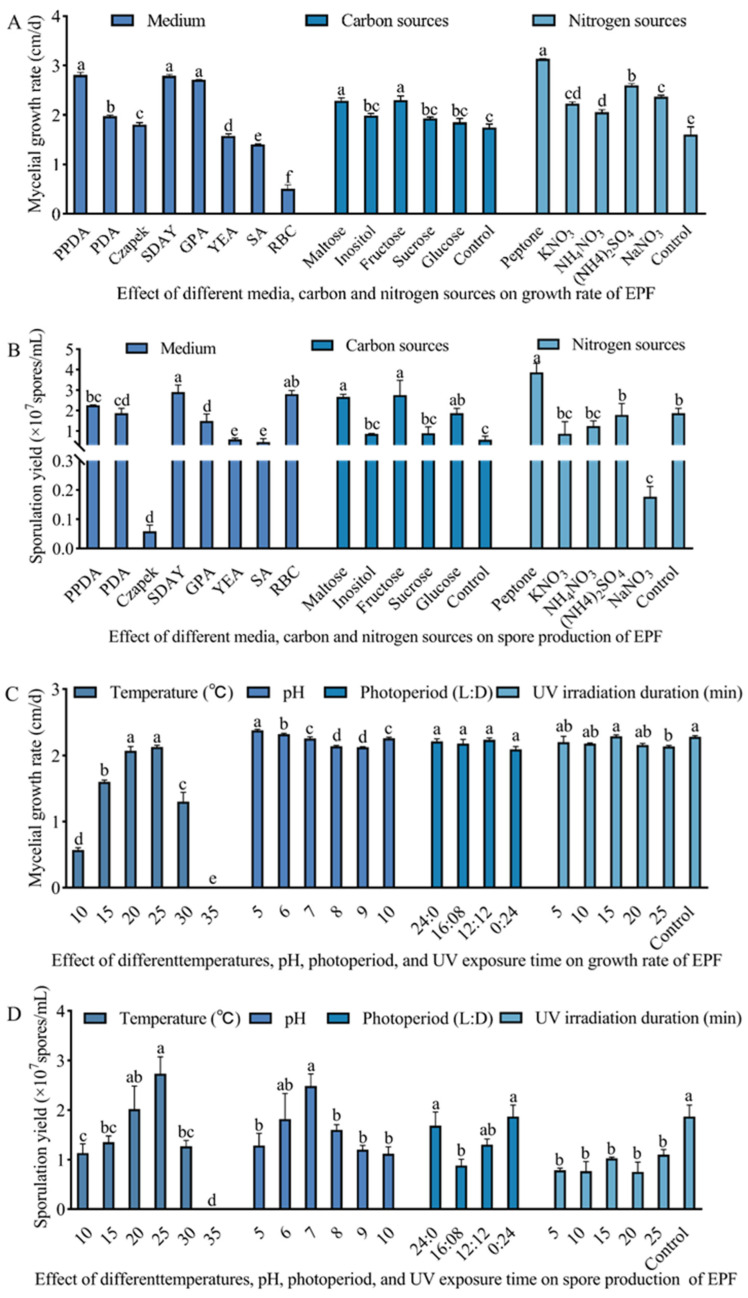 Figure 3
Figure 3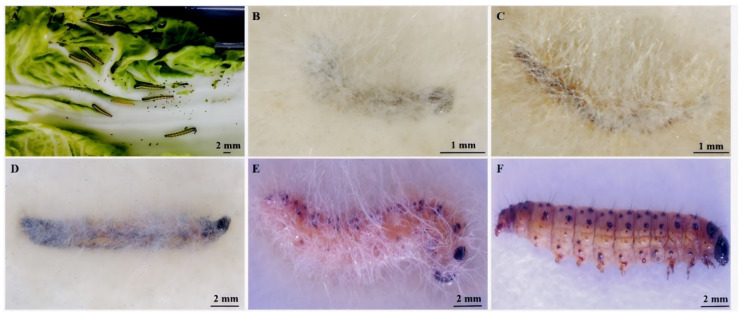 Figure 4
Figure 4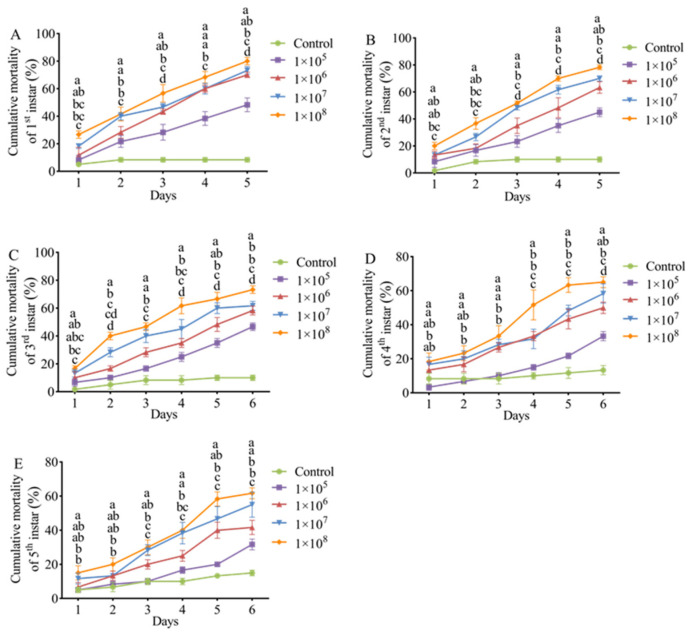 Figure 5
Figure 5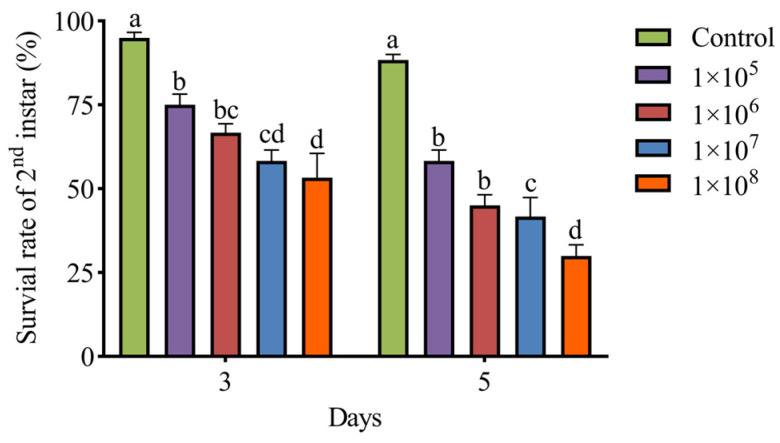 Figure 6
Figure 6- —Applied Fundamental Research Fund of Qinghai Provincial Department of Science and Technology
- —Major Project of Qinghai Academy of Agriculture and Forestry
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsEntomopathogenic Microorganisms in Pest Control · Insect Resistance and Genetics · Insect Pest Control Strategies
1. Introduction
Evergestis extimalis Scopoli (Lepidoptera: Pyralidae), a destructive pest, is mainly distributed in China, Korea, Japan, and the United States. It primarily infests rapeseed, fennel, cabbage, kale, and sugar beet crops. Its larvae burring into the rapeseed kernels and eating seeds cause serious harm to spring rapeseed, while the insect harms other host plants by leaf-rolling and eating core leaves and seed buds [1,2]. The spring rapeseed crop has the largest planting area in Qinghai Province, China, which was the primary income resource for local famers. However, the occurrence of E. extimalis has seriously threatened rapeseed production in Qinghai, causing great economic losses in recent years [1]. At present, E. extimalis is mainly controlled by chemical pesticides, which is limited by the emergence of strong resistance, environmental pollution, and concerns for human and animal health. Therefore, it is urgent to explore new alternatives for the management of E. extimalis, including a novel high-efficiency, low-toxicity, and low-residue microbial insecticide.
Entomopathogenic fungi (EPF) widely exist in nature and can infest insects, causing disease or even death [3]. They are considered highly effective in regulating the population of many Lepidoptera insect pests [4,5,6]. Shehzad et al. [7] reported strong pathogenicity of Beauveria bassiana and Metarhizium anisopliae against the second and third instar of Plutella xylostella under both direct spraying of inoculum and leaf dipping treatments. M. anisoplariae CQMa128 was shown to have a good control effect against the fourth instar larvae of Pyrausta nubilalis and Pieris rapae, with up to 95% and 76.7% mortality rates, respectively [8]. EPF of E. extimalis has rarely been reported in the literature. EPF may rapidly germinate and grow as a biological control agent [9]. However, the germination, growth, pathogenicity, and persistence of EPF can vary in different hosts and may be influenced by the strain types and environmental conditions [10]. For example, the growth and sporulation of most EPF usually reach their maximum at about 25 °C [11,12,13], and the maximum virulence is generally achieved at 20–25 °C [14].
Mucor spp. are widely distributed in nature, parasitizing plants and animals or exhibiting saprophytism in soil, plant, and animal residues, as well as in other substrates [15,16]. Some mucor species can infect certain Lepidoptera, Coleoptera, and Diptera insects [17]. Heitor et al. [18] isolated a virulent strain of M. hiemalis from Mamestra brassicae larvae. Reiss et al. [19] found M. hiemalis in Artemia salina. Konstantopoulou and Mazomenos [20] isolated four EPF from diseased pupae of Bactrocera oleae and larvae of Sesamia nonagrioides, and two strains of M. hiemalis were reported to have a good controlling effect on the adult of Ceratitis capitata. Konstantopoulou et al. [21] reported that the metabolic extract of M. hiemalis SMU-21 has insecticidal activity.
In this study, we identified a fungus infecting the larvae of E. extimalis and evaluated its pathogenicity. Our results show that it could be a potential biological control agent for the control of E. extimalis. Meanwhile, to protect the ecological environment of the Qinghai–Tibet Plateau, it is essential to research and develop biological control technologies for pest management.
2. Materials and Methods
2.1. Isolation of Entomopathogenic Fungus
The EPF strain was isolated from the carcasses of E. extimalis larvae, which were collected from Datong County (36°43′ N/100°51′ E), Qinghai Province, China. Briefly, the carcasses were soaked in 0.3% sodium hypochlorite for 1 min, washed with distilled water thrice, and then placed in a disposable Petri dish lined with sterile filter paper for moisturizing the culture [22]. When mycelia or spores were grown, a few were isolated in a sterile environment to inoculate on PDA (Potato Dextrose Agar) medium and cultured in an incubator at 23 ± 1 °C in the dark for 3 days. The next generation of mycelium was transferred into a new PDA plate and cultured five times to obtain the final isolate.
2.2. Morphological Characterization and Molecular Identification of the Isolated Fungus
The fungal isolate was inoculated on PDA medium, with a sterile and obliquely inserted coverslip, at 25 °C for 3 days. Subsequently, the grown mycelium was stained with lactic acid phenol cotton blue staining solution for morphological observation under an optical microscope. The pathogen was imaged and morphologically identified according to the Fungal Identification Manual [23].
A fungal genomic DNA extraction kit (Sangon Biotech, Shanghai, China) was used to extract the fungal DNA. Universal primers ITS1 (5′-TCCGTAGGTGAACCTGCGG-3′) and ITS4 (5′-TCCTCCGCTTATTGATATGC-3′) were used to amplify the ribosomal DNA internal transcriptional spacer (ITS) of the isolate. The small subunit 18S ribosomal DNA was amplified with NS1 (5′-GTAGTCATATGCTTGTCTC-3′) and NS6 (5′-GCATCACAGACCTGTTATTGCCTC-3′) primers. The primers were obtained from Shanghai Sangon Biological Co., Ltd., Shanghai, China. The PCR amplification reaction system consisted of 1 μL template DNA, 1 μL primers, 12.5 μL PCR MasterMix, and 9.5 μL dd H_2_O. The reaction conditions were as follows: pre-denaturation at 95 °C for 5 min, 30 cycles of denaturation at 94 °C for 30 s, annealing at 57 °C for 30 s, extension at 72 °C for 90 s, and a final extension at 72 °C for 10 min. The amplified products were detected and recovered by gel electrophoresis and sent to Shanghai Sangon Biological Co., Ltd., Shanghai, China, for sequencing. The sequencing results of the isolate were submitted to the GenBank nucleic acid sequence database for BLAST (https://ftp.ncbi.nlm.nih.gov/blast/) comparison, and sequences with higher homology were selected to construct a Neighbor-joining (NJ) phylogenetic tree with 1000 bootstrap calculations using MEGA 7.0 [24].
2.3. Optimization of the Culture Conditions of the Isolated Fungus
2.3.1. Influence of Medium on Fungal Growth and Sporulation
Fungal growth rate and spore production were evaluated in various media (n = 8) as described in Table S1. The 8 mm diameter agar plug containing mycelia was inoculated in the plate center of the different test media, and the culture was performed in the dark at 25 °C for 3 days. The colony diameter was measured by the cross-crossing method every day to estimate the fungal growth rate. Each test was repeated three times. After 6 d of culture, the spores were eluted in 10 mL of sterile water, and the solution was filtered through four layers of gauze to make a spore suspension. The spores were estimated by counting using a blood cell counting plate under an optical microscope.
2.3.2. Optimizing the Carbon and Nitrogen Sources for Fungal Growth and Sporulation
Using PDA as the base medium, glucose was replaced with fructose, sucrose, inositol, and maltose in equal amounts. The control group had no carbon source. Organic peptone was used as a nitrogen source, which was then replaced with equal amounts of sodium nitrate, ammonium sulfate, ammonium nitrate, and potassium nitrate. The control group contained no nitrogen source. Agar plugs containing mycelia (d = 8 mm) were cultured on media with different carbon and nitrogen sources at 25 °C in the dark for 3 days. All tests were repeated three times. The fungal growth rate and sporulation were calculated as described in Section 2.3.1.
2.3.3. Optimization of Temperature for Fungal Growth and Sporulation
Agar plugs containing mycelia (d = 8 mm) were inoculated on optimized PDA plates at 15, 20, 25, 30, and 35 °C, as described in Section 2.3.1, and the fungal growth rate and sporulation were calculated. All tests had three repeats.
2.3.4. Effect of pH on Fungal Growth and Sporulation
The pH values were adjusted to 5.0, 6.0, 7.0, 8.0, 9.0, and 10.0 with 1 mol/L HCl or 1 mol/L NaOH. Agar plugs containing mycelia (d = 8 mm) were inoculated on PDA media with different pH values, as described in Section 2.3.1. Average fungal growth rate and sporulation were calculated from three repeats at different pH values.
2.3.5. Optimization of Photoperiod for Fungal Growth and Sporulation
The incubator was programmed with four light/dark (L:D) cycles: 24:0, 16:8, 12:12, and 0:24, while the rest of the fungal culture conditions were the same as in Section 2.3.1. Average fungal growth rate and sporulation were estimated from three repeats at different photoperiods.
2.3.6. Effect of Ultraviolet (UV) Irradiation on Fungal Growth and Sporulation
The 8 mm diameter agar plugs containing mycelium were inoculated on PDA plates, which were placed at a vertical distance of 40 cm from a 30 W UV-C lamp in an ultra-clean bench. The lids of the Petri dishes were removed, and the plates were exposed to UV-C radiation for 5, 10, 15, 20, and 25 min. Each treatment was repeated 3 times. After the UV-C treatment, plates were cultured as described in Section 2.3.1, and the fungal growth and spore production rates were calculated.
2.4. Pathogenicity of EPF
2.4.1. Rearing of Insects and EPF Treatment
The insect eggs and larvae were collected from Datong County, Xining City, Qinghai Province, China, and brought back to the laboratory in a self-made incubator (8 cm × 14 cm × 5 cm). The larvae were fed with fresh cabbage (Brassica rapa) in an artificial climate incubator (temperature 22 ± 1 °C, relative humidity 70 ± 5%). Fungal isolates were cultured on SDAY medium for 8 days at 25 °C, after which they were used to prepare 1 × 10^5^, 1 × 10^6^, 1 × 10^7^, and 1 × 10^8^ spores/mL aqueous spore solution (containing 0.05% Tween 80) under aseptic conditions. An aqueous solution containing 0.05% Tween 80 was used as a blank control.
2.4.2. Bioassay Test
The dipping method was utilized for bioassay treated with the 1st to 5th instar larvae. The larvae (n = 15) were immersed in different spore suspensions for 20 s and then picked out to dry naturally. Subsequently, the larvae were transferred to an incubator (25 ± 1 °C, RH 70 ± 5%) and fed with fresh cabbage. Each treatment was repeated four times, and the mortality of larvae was observed every day.
2.4.3. Greenhouse Pot Experiment
Several pots of rapeseed (Qingza No. 5) were planted in the greenhouse of the Qinghai Academy of Agriculture and Forestry, China. Finally, 16 healthy well-grown rapeseed pots were selected at the seedling stage. Healthy 2nd instar larvae of E. extimalis were carefully transferred to the plants using a soft brush, 15 larvae per pot. Spore suspensions were sprayed on the potted plants in low to high concentrations (1 × 10^5^, 1 × 10^6^, 1 × 10^7^, and 1 × 10^8^ spores/mL), ensuring coverage of larval cuticle. There were four replicates for each spore concentration, and the sterile aqueous solution containing 0.05% Tween 80 was used as a blank control. Survival rate was calculated on the third and fifth days of treatment.
2.5. Data Analysis
Equations (1)–(3) were used to calculate E. extimalis larval cumulative mortality, survival, and cumulative corrected mortality. SPSS 26.0 and GraphPad Prism 8.0 were used for statistical analysis. Analysis of variance (ANOVA) and least significant difference (LSD) tests were performed for fungal growth, spore production, and insect mortality rates. LC_50_, LT_50_, and their 95% confidence limits (CI) were calculated by probit probability analysis.
3. Results
3.1. Identification of the EPF Strain
The fungus was cultured on a PDA medium for 3 days. The grown mycelium was white and feathery, with a dense center and sparse edges. Black sporangia could be observed at the top of the mycelium (Figure 1A). The fungus has 3.2–8.0 µm wide aseptate hyphae, and upon maturation of the sporangia releases ellipsoidal sporangiospores measuring 1.26–3.0 × 1.1–2.2 µm (Figure 1B).
The 535 bp ITS sequence of the fungal isolate shares 100% homology with M. hiemalis (such as MT573485 and MT279287). Additionally, the 1353 bp 18S sequence shares more than 99% homology with M. hiemalis. We named the strain M. hiemalis QH01 (GenBank accession numbers OK427267 and OQ730225) (Figure 2).
3.2. Culture Condition Optimization of M. hiemalis
We found marked differences in the growth and sporulation quantities of the M. hiemalis on different media (Figure 3A,B). The fastest growth rate of 2.32 cm/d was observed in the Peptone Potato Dextrose Agar (PPDA) medium, followed by the SDAY and Glucose Peptone Agar (GPA) media, which were not significantly different from each other. The slowest mycelial growth was 0.02 cm/d in the Bengal red medium (F_7,16_ = 25.85; p < 0.05). The highest sporulation quantity of 2.9 × 10^7^ spores/mL was in the SDAY medium, followed by Bengal red and PPDA media after 6 d of incubation. The lowest spore production of 0.06 × 10^7^ spores/mL was observed in the Czapek–Dox medium, which was significantly less than in other media (F_7,16_ = 358.15; p < 0.05). Thus, the SDAY medium was screened as the most suitable medium for the growth and spore production of M. hiemalis QH01.
As for the carbon source, strain QH01 grew fastest on medium with fructose, with a growth rate of up to 2.30 cm/d (Figure 3A,B). Mycelial growth was also enhanced with maltose and glucose, with a significant increase compared to the control group (F_5,12_ = 13.23; p < 0.05). The fungus produced the most spores on a fructose-containing medium, followed by media containing glucose or maltose, both of which were significantly better than the control (F_5,12_ = 7.73; p < 0.05). The growth and sporulation quantity of the fungus on the sucrose- or inositol-supplemented medium were not significantly different from the control medium.
Regarding the nitrogen sources, the fungal mycelial growth rate on the peptone-supplemented medium was 3.13 cm/d, which was significantly higher than for the other nitrogen sources (such as KNO_3_, NaNO_3_, (NH_4_)2_SO_4, and NH_4_NO_3_, F_5,12_ = 51.37; p < 0.05). The fungus produced the most spores on a peptone-containing medium, while the medium containing KNO_3_, (NH_4_)2_SO_4, or NH_4_NO_3_ was not significantly different from the control group. Notably, the spore production was severely inhibited and lower than the control when the fungus was grown with NaNO_3_ as the nitrogen source (F_5,11_ = 11.53; p < 0.05). Thus, fructose and peptone were selected as the optimum carbon and nitrogen sources for the growth and spore production of strain QH01 (Figure 3A,B).
Strain QH01 was able to grow from 10 to 30 °C, but the difference was significant (Figure 3C,D). At 25 °C, the fastest growth rate was 2.13 cm/d (F_5,12_ = 156.02; p < 0.05). The slowest fungal growth was at 10 °C. Under optimal temperature of 25 °C, the fungus produced 2.73 × 10^7^ spores/mL (F_5,12_ = 12.72; p < 0.05). No mycelial growth or spore production was observed at 35 °C.
Medium pH exerted a significant influence on both fungal mycelial growth and sporulation. The maximum mycelial growth rate of 2.38 cm/d was recorded at pH 5.0 (F_5,12_ = 35.18; p < 0.05). The optimum pH for fungal spore production was 6–7, but a too-acidic or alkaline medium reduced spore production (F_5,12_ = 3.70; p < 0.05). No significant differences in fungal mycelial growth rates were observed across the tested photoperiods (F_3,8_ = 2.02; p > 0.05). In contrast, sporulation quantity was affected under different treatments (F_3,8_ = 4.76; p < 0.05). The highest spore production (0.87 × 10^7^ spores/mL) was in the dark (24 h dark), and the lowest (0.88 × 10^7^ spores/mL) was at 16L:8D. The mycelial growth rate remained unaffected, showing no significant difference from the control treatment after the fungus was irradiated with UV-C light for 5–20 min (F_5,9_ = 1.91; p > 0.05). Compared to the control treatment, however, the sporulation quantity was obviously reduced, and there was little difference between different time treatments (F_5,10_ = 7.47; p < 0.05) (Figure 3C,D).
3.3. E. extimalis Larvae Infestation with M. hiemalis QH01
Since E. extimalis larvae were infected by M. hiemalis QH01, their feeding rate gradually decreased. The dead larvae turned brown or black-red, and the body softened and finally dried up. The larvae body surface became covered with white mycelium and black sporangia after 2–5 days of moisturized incubation at 25 °C (Figure 4).
3.4. Pathogenicity of M. hiemalis QH01 Against E. extimalis Larvae
E. extimalis larvae infected with M. hiemalis spore suspensions exhibited significant dose- and time-dependent mortality compared to the control group (Figure 5). The spores induced mortality in all ages of insect larvae.
The LC_50_ values of M. hiemalis QH01 spores against E. extimalis larvae of various ages are listed in Table 1. The LC_50_ exhibited a time-dependent decrease, with larval susceptibility showing an inverse relationship to developmental maturity (first instar > second instar > third instar > fourth instar > fifth instar). Basically, the younger larvae were more sensitive to M. hiemalis QH01.
The LT_50_ values of M. hiemalis QH01 against E. extimalis larvae progressively decreased with increasing inoculum concentration. The fungus exhibited the fastest lethal effect against the 1st–5th instar larvae treated with 1 × 10^8^ spores/mL suspensions, and LT_50_ values were 2.99, 3.01, 3.37, 4.04, and 4.40 days, respectively. In contrast, under 1 × 10^5^ spores/mL concentration, the fungus had the slowest lethal effect, and LT_50_ values were 5.15, 5.38, and 5.29 days for 1st–3rd instar larvae. With this spore concentration, the mortality rate of 4th-5th instar larvae was lower than 50% even after 6 days of exposure, and it was not possible to estimate the LT_50_ (Table 2). These results indicated that M. hiemalis QH01 is highly virulent to young larvae.
3.5. Greenhouse Experiments
The results of greenhouse pot experiments of M. hiemalis QH01 on second instar larvae of E. extimalis are shown in Figure 6. By spraying M. hiemalis suspensions of 1 × 10^5^, 1 × 10^6^, 1 × 10^7^, and 1 × 10^8^ spores/mL, the survival rates of second instar larvae at 5 days were 58.33%, 45%, 41.67%, and 30%, respectively, which were significantly lower than control group (F_4,15_ = 37.47; p < 0.05). The suspension containing 1 × 10^8^ spores/mL resulted in the lowest survival rate, indicating a concentration-dependent survival.
4. Discussion
The composition of the culture medium can affect the growth characteristics of EPF [25]. We found that the SDAY medium was the most suitable for the growth and spore production of M. hiemalis QH01, which was consistent with previous studies [26]. The spore yield of other tested media, such as PDA, GPA, Yeast extract agar (YEA), etc., was relatively lower than that of SDAY, which might have been due to the difference in selective absorption of nutrients by the same strain because of the different types, contents, and solubility states of nutrients in different media. In addition, carbon and nitrogen sources can also affect the growth and spore production of EPF [27]. This study demonstrated that fructose favored the growth and spore production of M. hiemalis QH01, while the absence of a carbon source inhibited it. The best growth and spore production of strain QH01 were observed with peptone as a nitrogen source, which was consistent with Huang et al. [28].
The optimal growth and reproduction of EPF cannot be achieved without appropriate environmental conditions [29]. Temperature affects the metabolic activity of entomopathogenic fungi through key physiological processes including enzyme biosynthesis, toxin production, spore germination, and germ tube penetration [30]. We found that M. hiemalis QH01 exhibited a growth and sporulation temperature range of 10–30 °C, with an optimum at 25 °C. In contrast, M. hiemalis BO-1 grew the fastest at 23 °C and produced the highest sporulation at 18 °C [31]. This is likely attributable to QH01’s specific adaptations to the extreme climatic conditions of its native Tibetan Plateau environment. The medium pH can significantly impact the growth and pathogenicity of EPF [32]. For example, pH 5.5–6.5 is best suited for the sporulation of B. bassiana, and the same is also good for its pathogenicity [33,34]. M. hiemalis QH01 can grow and produce spores in a wide range of pH (5–10), which is consistent with the results of Wang et al. [35]. In addition to temperature and pH, external conditions such as natural light have a certain impact on EPF. In this study, M. hiemalis had the highest sporulation in total darkness. This may be attributed to Mucor spp.’s natural adaptations to light-limited environments, given their predominant colonization of soil and decaying organic substrates [16]. Their spore production mechanism may have evolved a dependence on darkness. UV light usually inhibits the growth, sporulation, and pathogenicity of EPF. We found that UV irradiation decreased spore production in M. hiemalis. Yao et al. [36] reported that 312 and 365 nm UV light exposure decreased the pathogenicity of M. anisopliae against tobacco aphids. The influence of ultraviolet light on the pathogenicity of M. hiemalis had to be considered when applying it to pest control in the field.
So far, there have been few reports on the biological control of E. extimalis. Wang et al. [37] conducted a field investigation and reported that the Cloth beetle and B. bassiana were the natural enemies of the insect in the larval stage, while the pupal stage could be parasitized by mites. Here, we show the pathogenicity of M. hiemalis against E. extimalis. Particularly, the young larvae of E. extimalis were more susceptible to M. hiemalis QH01, while the older larvae exhibited relatively a lower mortality rate. This difference can be attributed to the different body wall structures and immune capacities of young and old larvae. A thinner body wall in younger larvae makes them more susceptible to EPF infection. Older larvae are more resistant to disease due to hardening and thickening of the cuticle, enhanced immunity and metabolism [38]. Moreover, with the increase in the developmental stage of E. extimalis, the corresponding LT_50_ and LC_50_ values increased for the same concentration of M. hiemalis. This indicated that M. hiemalis QH01 is more virulent to lower-stage larvae than the developed larvae. This result is in agreement with the findings of Tozlu et al. [39]. Zhu et al. [40] reported 46.25% mortality in Bradysia impatiens second instar larvae following 5 days of exposure to M. hiemalis BO-1 at 1 × 10^6^ spores/mL, whereas our experimental data demonstrated 63.33% mortality in E. extimalis larvae under identical spore concentration with M. hiemalis QH01. This may be the result of host specificity, environmental adaptation, etc., which led to significant differences in the pathogenicity of strains of the same species. Notably, surviving larvae exhibited significantly reduced feeding activity, progressed to the prepupal stage with sporadic pupation, but ultimately failed to complete adult eclosion. It is possible that infection by M. hiemalis QH01 induces a sublethal effect on life history traits including developmental processes and reproduction in E. extimalis [41]. EPF can target all developmental stages of insects, such as eggs, larvae, and adults. Wang et al. [42] isolated a strain of M. hiemalis from diseased Bradysia odoriphaga larvae and found its pathogenicity varied against different stages of insect; strong pathogenicity was against larvae and relatively weak pathogenicity was against eggs and pupae. In the study, the pathogenicity of M. hiemalis QH01 was determined only against the larvae of E. extimalis, and further studies are needed to determine the pathogenicity against other insect forms.
Most EPF performs remarkably well under controlled conditions, such as in the laboratory, exhibiting high mortality against the target insects. However, abiotic factors such as changes in temperature, humidity, and UV exposure can significantly affect the biocontrol effect of EPF in field applications [43]. In greenhouse environments, after 7 days of treatment with 2 × 10^6^ spores/mL of Bea111 (B. bassiana) and 9 × 10^6^ spores/mL of Isa340 (Isaria javanica), third instar larvae of Duponchelia fovealis exhibited 52% and 45% mortality rates, respectively [44]. The larval stage of E. extimalis causes the most damage and economic loss in rapeseed fields. First instar larvae exhibit higher mortality rates when attempting to bore into rapeseed pods for feeding. From the third instar onward, larvae exhibit migratory behavior, transitioning from pod-boring to stem-tunneling and inter-plant movement, while their food consumption surges dramatically [45]. Thus, we evaluated the biocontrol potential of M. hiemalis QH01 against the second instar larvae of E. extimalis under greenhouse conditions and found it significantly reduced their survival rates.
In this study, the effectiveness of strain QH01 against E. extimalis second instar larvae in greenhouse pots differed from the results of the indoor virulence assay, which may be due to different bioassay methods [46] and external environmental conditions. To sum up, when preparing fungal powder, ultraviolet protectants, thermal stabilizers, and ionic dispersants should be added to the spore powder to enhance environmental adaptability. Greenhouse and field applications should be carried out after sunset or on cloudy days when the temperature is around 20–25 °C.
5. Conclusions
In the present study, a fungal strain isolated from E. extimalis larvae was identified as M. hiemalis QH01 based on morphological characteristics, ITS, and 18S rDNA sequence analyses. To our knowledge, this is the first report assessing the biocontrol effects of an entomopathogenic fungus against E. extimalis. Our results lay the foundation for the subsequent research on designing a green prevention and control program for E. extimalis using EPF to benefit the spring rapeseed industry.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Shao H.N. Liu Y.X. Liu Y.J. Lai Y.P. The Effect of Ice-Nucleation-Active Bacteria on Metabolic Regulation in Evergestis extimalis (Scopoli) (Lepidoptera: Pyralidae) Overwintering Larvae on the Qinghai-Tibet Plateau Insects 20221390910.3390/insects 1310090936292857 PMC 9604135 · doi ↗ · pubmed ↗
- 2Lai Y.P. Tao K. Hou T.P. Preliminary analysis of geographical distribution based on cold hardiness for Evergestis extimalis (Scopoli) (Lepidoptera: Pyralidae) on Qinghai-Tibet Plateau Entomol. Res.201949132010.1111/1748-5967.12262 · doi ↗
- 3Shah P.A. Pell J.K. Entomopathogenic fungi as biological control agents Appl. Microbiol. Biotechnol.20036141342310.1007/s 00253-003-1240-812764556 · doi ↗ · pubmed ↗
- 4Samuels R.I. Coracini D. Martins dos Santos C.A. Gava C.A.T. Infection of Blissus antillus (Hemiptera: Lygaeidae) eggs by the entomopathogenic fungi Metarhizium anisopliae and Beauveria bassiana Biol. Control 20022326927310.1006/bcon.2001.1009 · doi ↗
- 5Yao H.Q. Xie Y.P. Wang E.H. Morphologic observation of Apocheima cinerarius Erschoff (Lepidoptera: Geometridae) pupae and comparison of virulence of Beauveria bassiana infected on them J. Environ. Entomol.201537348354
- 6Cruz-Avalos A.M. Bivián-Hernández M.L.Á. Ibarra J.E. Del Rincón-Castro M. High virulence of mexican entomopathogenic fungi against fall armyworm, (Lepidoptera: Noctuidae)J. Econ. Entomol.20191129910710.1093/jee/toy 34330383250 · doi ↗ · pubmed ↗
- 7Shehzad M. Tariq M. Mukhtar T. Gulzar A. On the virulence of the entomopathogenic fungi, Beauveria bassiana and Metarhizium anisopliae (Ascomycota: Hypocreales), against the diamondback moth, Plutella xylostella (L.) (Lepidoptera: Plutellidae)Egypt. J. Biol. Pest Control 202131869210.1186/s 41938-021-00428-z · doi ↗
- 8Xie N. Yin Y.P. Zhang J.W. Shen J.F. Wang Z.K. Characterization of the Metarhizium anisopliae CQ Ma 128 and its virulence measurements against Lepidoptera insects Proceedings of the 2009 Annual Meeting of the Mycological Society of China Beijing, China 23–25 August 2009