Association Analysis of SLC11A1 Polymorphisms with Somatic Cell Score in Chinese Holstein Cows
Kai Liu, Yufang Liu, Tuo Li, Qiuling Li, Jinyu Wang, Yongfu An, Yuze Yang, Kaiyang Li, Mingxing Chu

TL;DR
This study found genetic variations in the SLC11A1 gene that are linked to lower somatic cell scores in Chinese Holstein cows, which could help improve mastitis resistance and milk quality.
Contribution
The study identifies specific SNPs in the SLC11A1 gene associated with somatic cell scores, suggesting their potential use as molecular markers for improving mastitis resistance in dairy cows.
Findings
The SNP c.723C>T in SLC11A1 was significantly associated with lower somatic cell scores in Chinese Holstein cows.
The A allele of c.723C>T is a potential marker for improving mastitis resistance in dairy cows.
Bioinformatic analysis suggests the SNPs may affect SLC11A1 mRNA structure and function.
Abstract
Milk production traits, including milk yield, fat content, and protein content, are critical economic indicators in the dairy industry. Somatic cell score (SCS) is another important metric, as it reflects the health status of dairy cows and impacts milk quality. The SLC11A1 gene, which is involved in the transport of divalent metals such as iron and manganese, has been implicated in influencing immune responses and potentially milk production traits. Single nucleotide polymorphisms (SNPs) are variations at a single nucleotide position in the DNA sequence and play a vital role in genetic studies of milk production traits. In this research, we identified two SNPs within the SLC11A1 gene and analyzed their associations with SCS. The results indicated that specific SNPs in the SLC11A1 gene were significantly associated with SCS, suggesting that these genetic variations could influence the…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
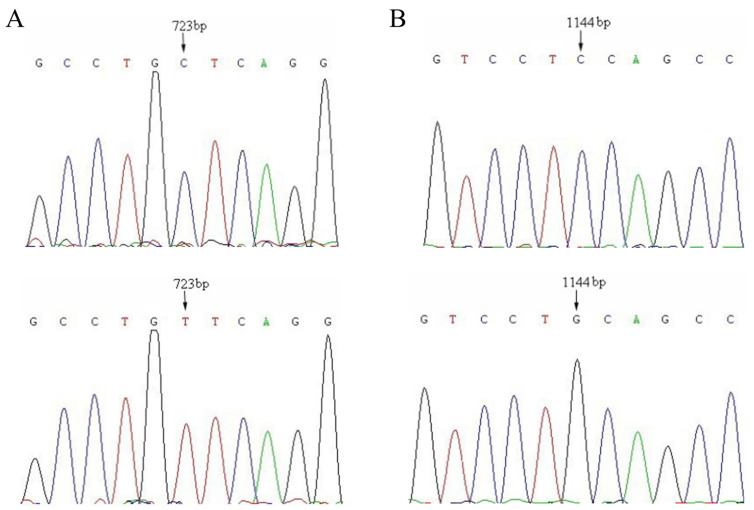 Figure 1
Figure 1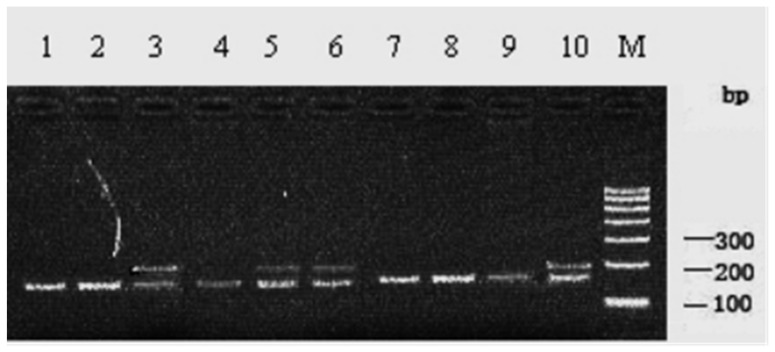 Figure 2
Figure 2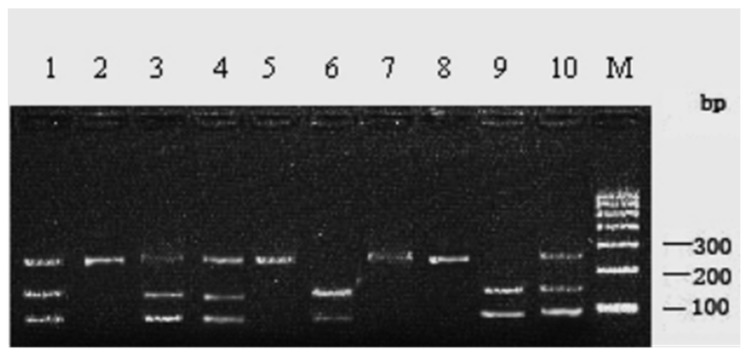 Figure 3
Figure 3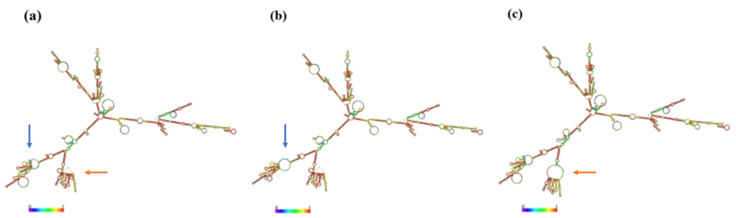 Figure 4
Figure 4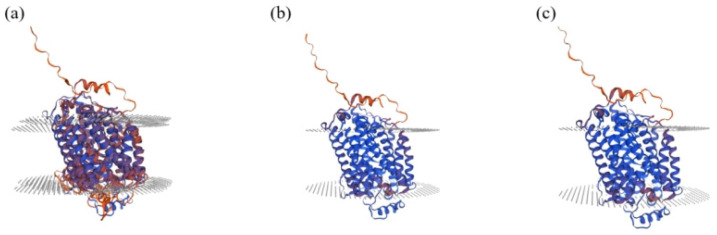 Figure 5
Figure 5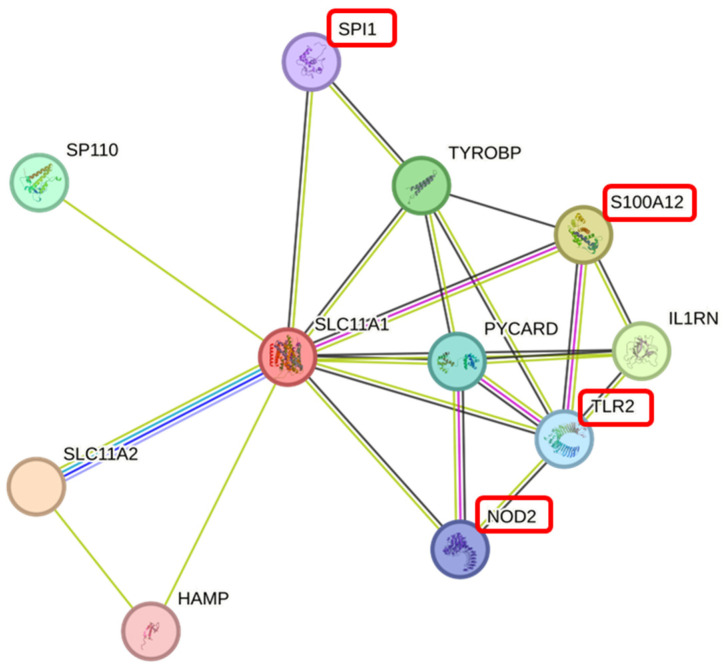 Figure 6
Figure 6- —China Agriculture Research System of MOF and MARA
- —Agricultural Science and Technology Innovation Program of China
- —National Key Technology Research and Development Program of China
- —Beijing Natural Science Foundation
- —Science and Technology Project of Hebei Education Department
- —Scientific Research and Innovation Team of Basic Scientific Research Project of Provincial Universities in Hebei Province
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsMilk Quality and Mastitis in Dairy Cows · Animal Genetics and Reproduction · Genetic and phenotypic traits in livestock
1. Introduction
Mastitis is the most common and costly disease on dairy farms, with adverse effects including reduced milk production, impaired fertility and premature culling [1,2]. Traditional breeding strategies for mastitis resistance based on phenotypic selection have not been very successful due to the low heritability of mastitis resistance traits [3]. An indirect selection strategy to reduce mastitis is based on milk somatic cell score (SCS), which is strongly and positively correlated with clinical mastitis [4,5]. Single nucleotide polymorphism (SNPs) is the third generation of genetic markers, which arealterations in the DNA sequence where one nucleotide is replaced by another at a specific genomic position, deviating from the reference genome. In livestock breeding, SNPs are important in determining an animal’s genotype and its relationship to production, reproductive and economic traits [6,7,8]. SNPs and genomic linkage technology have been used to identify genes for milk production traits in dairy cows [9]. The impact of mastitis can be reduced by genetic testing and indirect selection for cows with lower SCS [10,11]. Natural resistance-associated macrophage protein 1 (NRAMP1), also known as solute carrier 11A1 (SLC11A1), is a member of the solute carrier family 11 [12,13,14]. SLC11A1 is a highly conserved gene and was first identified in mouse IC3 [15]. It is located on bovine chromosome 2q43-44, contains 16 introns and 15 exons, and SLC11A1 encodes a multichannel membrane protein that functions by translocating divalent cations (iron and manganese) involved in iron metabolism and host resistance to specific pathogens [13]. Its role in resistance or susceptibility to bacterial infection makes it a promising candidate gene for breeding disease-resistant animals [16]. This gene plays an important role in innate immunity by preventing bacterial growth in macrophages during the early stages of infection [17]. Mutations in SLC11A1 have been found to confer susceptibility to a variety of intracellular pathogens, including Mycobacterium, Salmonella and Leishmania [18,19,20]. In addition, the SLC11A1 gene has been found to be associated with natural resistance to brucellosis in cattle [21,22] and buffalo [23,24]. It is noteworthy that SNP in the SLC11A1 gene was identified as a significant genetic marker and susceptibility factor for mastitis tolerance/susceptibility in both Holstein and Swiss Brown cows [25]. Furthermore, a mutation was identified at 1139bp in exon 11 of the SLC11A1 gene, which was significantly associated with the development of clinical mastitis [26]. Furthermore, a comparison of the transcriptomes of circulating leukocytes from healthy cows with those from cows with naturally occurring subclinical or clinical mastitis revealed some differentially expressed genes, including SLC11A1 [27]. These results suggest that SLC11A1 is a candidate gene for disease resistance in cattle. Since SLC11A1 is involved in innate immunity by killing bacteria, we hypothesized that it is a candidate gene for mastitis resistance.
In the present study, we analyzed the relationship between polymorphisms in SLC11A1 and SCS in Chinese Holstein cows, and the potential biological functions of SNPs in this gene. The results may provide a theoretical basis for marker-assisted selection for mastitis resistance in Chinese Holstein cows.
2. Materials and Methods
2.1. Blood Sample Collection and DNA Preparation
The 210 Holstein cows involved in this study came from four dairy farms in Hebei Province, China (70 from Farm 1, 70 from Farm 2, 35 from Farm 3, and 35 from Farm 4). These cows are the progeny of five bulls with 39, 41, 42, 43 and 45 offspring respectively. The number of cows at three parities was 65 (first birth), 71 (second birth) and 74 (third birth), respectively. This study collected data on various factors related to dairy cows. These included recording of body condition scores on a scale of 1 to 5 in increments of 0.25 with 3 being the ideal score. In addition, milk yield data were collected daily from each cow throughout the lactation period. Stage of lactation: Days in milk (DIM) categorized as early (1–100 DIM), mid (101–200 DIM), and late (>200 DIM) per dairy management standards. Other relevant data such as health status and management practices were also recorded. The average SCC (somatic cell count) of the cows on the four farms was 700,000/mL, 550,000/mL, 400,000/mL and 450,000/mL. Blood was collected from the jugular vein of each cow in 10 mL samples, anticoagulated with ACD and stored at −20 °C. Genomic DNA was extracted from blood samples using the phenol-chloroform method, then dissolved in TE buffer (10 mmol/L Tris-HCl, 1 mmol/L EDTA) and stored at −20 °C.
Danish electronic counter (Series 300 Fossomatic Type 75200) was used to determine SCC of 210 Holstein cows’ milk once per month, for a maximum of 10 times each cow. The average of these counts was the SCC of the parity. Hypothesis testing requires that errors are normally distributed and subgroup variances are homogeneous in analysis of variance. Therefore, the SCC of abnormal distribution characteristic must be converted to SCC of normal distribution before the analysis. Ali and Shook [28] reported that the converted SCC data have widely normal distribution and homogeneity characteristic, the logarithm conversion of SCC is the best way to obtain normal distribution. Somatic cell score (SCS = log_2_^(SCC/100)^ + 3, the unit of SCC is 1000 cells/mL) has the normal distribution characteristic, has been the criterion measure of SCC of the National Cooperative Dairy Herd Improvement Program of USA. The number, father’s number, farm, parity and SCS of each cow were recorded.
2.2. Primer Sequences and PCR Amplification
Primers were designed using the bovine SLC11A1 mRNA sequence. Primer sequences, target fragment lengths and annealing temperatures are listed in Table 1. The polymerase chain reaction (PCR) was performed in a 25 μL volume containing approximately 2.5 μL 10× PCR buffer (50 mmol/L KCl, 10 mmol/L Tris-HCl (pH 8.0), 0.1% Triton X-100), 1.5 μL MgCl_2_ (1.5 mM), 2.0 μL dNTPs (2.5 mM each), 0.5 μL each forward and reverse primers (20 μM), 50 ng genomic DNA and 1 U Taq DNA polymerase (SABC, Beijing, China). PCR conditions were as follows: pre-denaturation at 94 °C for 6 min; followed by 32 cycles of denaturation at 94 °C for 30 s, annealing for 30 s, extension at 72 °C for 30 s; with a final extension at 72 °C for 10 min, all operations were performed on a Mastercycler^®^ 5333 (Eppendorf AG, Hamburg, Germany).
2.3. Single Strand Conformation Polymorphism (SSCP) Detection
A volume of 3 μL PCR product was transferred to an Eppendorf tube and mixed with 7 μL gel loading solution containing 98% formamide, 0.025% bromophenol blue, 0.025% xylene cyanol, 20 mmol/L EDTA (pH 8.0), 10% glycerol. The mixture was agitated and denatured at 98 °C for 10 min, rapidly placed on ice for 7 min and loaded onto 10–12% neutral polyacrylamide gels (acrylamide:bisacrylamide = 29:1). Electrophoresis was performed in 1× Tris borate (pH 8.3)-EDTA buffer at 120 V for 15–20 h at 4 °C. After electrophoresis, the DNA fragments in the gels were visualized by silver nitrate staining, photographed and analyzed using an AlphaImager^TM^ 2200 and 1220 Documentation and Analysis System (Alpha Innotech Corporation, San Leandro, CA, USA).
2.4. Cloning and Sequencing
Amplification products from P1-P16 were analyzed by SSCP. Each DNA fragment was inserted into the pGEM-T Easy vector (Promega) according to the manufacturer’s instructions at 16 °C overnight. Ligation reactions were performed in 10 μL volumes containing PCR product 1 μL, pGEM-T Easy vector (50 ng/μL) 1 μL, T4 ligase (3 U/μL) 1 μL, 2× ligation buffer 5 μL, ddH_2_O 2 μL. The recombinant plasmid was transformed into competent Escherichia coli DH5α. Positive clones were identified by restriction enzyme digestion. Five clones of each genotype were selected for sequencing. Each clone was sequenced twice on an ABI3730 automated sequencer (Perkin Elmer Applied Biosystems, Foster City, CA, USA) by Shanghai Invitrogen Biotechnology Ltd. Co. (Shanghai, China).
2.5. Restriction Fragment Length Polymorphism (RFLP) Analysis
Two mutations (c.723C>T and c. 1144C>G) were detected in SLC11A1 by sequencing. Based on the sequences, we found two restriction enzymes to carry out RFLP analysis. Restriction enzyme reaction was carried out in 15 μL volume by mixing 5 μL of PCR products, 5 U Bpu10I (New England Biolabs, Beverly, MA, USA) or 5 U PstI (New England Biolabs, Beverly, MA, USA) and 1 μL corresponding 10× reaction buffer and incubating at 37 °C overnight. The resulting fragments were separated by electrophoresis on 2% agarose gel (Promega) in parallel with a 100 bp DNA marker.
2.6. Bioinformatics Analysis
RNAFOLD 2.6.3 (http://rna.tbi.univie.ac.at//cgi-bin/RNAWebSuite/RNAfold.cgi (accessed on 22 February 2024)) was used to predict the mRNA secondary structure of wild-type and mutated SLC11A1. The tertiary structure of the target proteins was analyzed using SWISS-MODEL software [29] (https://swissmodel.expasy.org/ (accessed on 22 February 2024)). STRING 11.0 software (https://string-db.org/cgi/input.pl (accessed on 23 February 2024)) was used to analyze SLC11A1 proteins interactions.
2.7. Statistical Analysis
The following statistical model was fitted to compare difference of SCS among SLC11A1 genotypes.
where y_ijklm_ is SCS, μ is population mean, S_i_ is the fixed effect of the ith bull (i = 1, 2, 3, 4, 5), H_j_ is the fixed effect of the jth dairy farm, P_k_ is the fixed effect of the kth birth (k = 1, 2, 3), G_l_ is the fixed effect of the lth genotype, e_ijklm_ is the random error effect of each observation. Bull Effect (S_i_): The inclusion of the bull effect accounts for the genetic relatedness among the cows, as they are progeny of five different bulls. This ensures that the genetic background of the cows is considered in the analysis. Farm Effect (H_j_): The farm effect adjusts for environmental and management differences across the four dairy farms, ensuring that farm-specific factors do not confound the results. Parity Effect (P_k_): The parity effect accounts for differences in SCS due to the lactation stage of the cows, as SCS can vary with parity. Genotype Effect (G_l_): This is the primary factor of interest, representing the association between SLC11A1 genotypes and SCS. eijklm: The random error term captures unexplained variability in SCS values, ensuring that the model accounts for individual differences not explained by the fixed effects. Calculations were achieved using Proc GLM (General Linear Model) of SAS (V.8.12). Mean separation procedures were performed using a least significant difference test.
3. Results
3.1. SSCP Detection and Genotyping
SSCP genotypes were identified by mobility shift due to conformational difference of the single-stranded DNAs of the amplified fragments by each primer, which is caused by nucleotide variation. The results showed polymorphisms only in P8 and P11 in the CDS in SLC11A1 gene. Sequence alignment between different genotypes of primer P8 revealed a C to T mutation at the 723 locus (exon 8) according to the mRNA sequence of the bovine SLC11A1 (GenBank accession number NM_174652) of exon 8 (Figure 1), and this mutation caused an amino acid change (Ala to Val) at residue 217 (A217V). Sequence alignment between different genotypes of primer P11 revealed a C to G mutation at position 1144 according to the mRNA sequence of bovine SLC11A1 (GenBank accession number NM_174652) (Figure 1), and c.1144C>G caused an amino acid change (Pro to Ala) at residue 356 (P356A). The C to T transition at site 723 and the C to G transversion at site 1144 changed the recognition site of the restriction endonuclease Bpu10I and PstI, respectively, which could be detected by PCR-RFLP. Both mutation sites are missense mutations.
3.2. Bpu10I Analysis of the c.723C>T of SLC11A1
The 199 bp PCR product with primer P8 was digested completely with the restriction endonuclease Bpu10I and genetic polymorphisms were detected by PCR-RFLP technique. Two genotypes were identified in 210 Chinese Holstein cows, which are AA (199 bp) and AB (42 bp/157 bp/199 bp) (Figure 2).
3.3. PstI Analysis of the c.1144C>G of SLC11A1
The 243 bp PCR product with primer P11 was digested completely with the restriction endonuclease PstI and genetic polymorphisms were investigated by PCR-RFLP. Three genotypes (CC, 243 bp; CD, 95 bp/148 bp/243 bp; DD, 95 bp/148 bp) were detected in 210 Chinese Holstein cows (Figure 3).
3.4. Genetic Characteristics of Polymorphic Sites of SLC11A1 Gene in Chinese Holstein Cows
It was shown that AA and CC were the dominant genes at the c.723C>T and c.1144C>G loci, with genotype frequencies of 0.790 and 0.629, respectively. Both loci are in Hardy-Weinberg equilibrium, indicating that allele and genotype frequencies are stable in the population. The c.723C>T locus has a PIC of less than 0.25, which is a low polymorphism; the c.1144C>G locus has a PIC between 0.25 and 0.5, which is a moderate polymorphism (Table 2).
3.5. Association of Two Polymorphic Loci of the SLC11A1 with SCS in Chinese Holstein Cows
As shown in Table 3, for the c.723C>T polymorphic locus, the mean value of SCS of the AA type was significantly lower than that of the AB type (p < 0.05), and AA was the favorable genotype and AB was the unfavorable genotype for mastitis resistance. For the c.1144C>G polymorphic locus, CC, CD, and DD genotypes, the SCS means were not significantly different (p > 0.05).
3.6. Bioinformatics Analysis of Chinese Holstein Cows SLC11A1
Prediction of the secondary structure of SLC11A1 mRNA from Chinese Holstein cows showed that the c.723C>T mutation and the c.1144C>G mutation resulted in changes in the secondary structure of the mRNA, and the minimum structural free energy increased from −770.17 kJ/mol to −768.67 kJ/mol and −768.57 kJ/mol, respectively, which decreased the stability of the mRNA secondary structure (Figure 4). It was predicted that the secondary structure of the proteins before and after the mutation would change slightly. 52.92% of the secondary structure of the proteins before the mutation was alpha helix, 12.04% was extended strand, 3.65% was beta turn and 31.39% was random coil. After the c.723C>T mutation, 53.47% was alpha helix, 12.59% was extended strands, 3.83% was beta turn and 30.11% was random coil. After the c.1144C>G mutation, 52.92% was alpha helix, 10.95% was extended strands, 3.65% was beta turn and 32.48% was random coil. The tertiary structure of the SLC11A1 protein also changed before and after the mutation (Figure 5).
3.7. The PPI Network of the SLC11A1 Protein
To further analyze the function of the SLC11A1 protein in Chinese Holstein cows, the network of target and related proteins was predicted using the STRING online tool. SLC11A1 was closely associated with the expression of genes involved in the regulation of the immune response and inflammation, with S100A12, NOD2, TLR2 and SPI1 being closely associated with mastitis resistance (Figure 6).
4. Discussion
Mastitis poses a significant challenge to the dairy industry, leading to considerable economic losses due to decreased milk yield and quality. It also has a substantial impact on the health and reproductive performance of dairy cows. Furthermore, mastitis is critical to maintaining public health [30,31,32]. The SLC11A1 gene, previously defined as the Natural Resistance Associated Macrophage Protein 1 gene (NRAMP1), encodes an iron/divalent cation transmembrane protein located in phagocytosed lysosomal late membranes, particularly in macrophages [33]. SLC11A1 plays a critical role in innate immunity [34] and is also important for macrophage recycling of metals (including iron) and digestion of apoptotic cells and aged erythrocytes [35,36]. In addition, microsatellite polymorphisms in the 3′ untranslated region (UTR) of the SLC11A1 have been reported to confer protection against Brucella abortus [23,24,37], Mycobacterium Bovis [20,38], and Mycobacterium avium ssp. Paratuberculosis [39]. Moreover, the gene has been reported to be associated with natural resistance to mastitis in cattle [26].
In our study, we identified two mutation sites in the SLC11A1 gene: c.723C>T and c.1144C>G. The c.723C>T locus contains two genotypes, of which type AA is in the majority and type AB is in the minority. The c.1144C>G locus contains three genotypes, of which type CC is in the majority and type CD and DD are in the minority. This may be due to the elimination of another pure type, possibly a recessive disease-causing gene, over a long period of time during the selection process. In addition, the PIC value of the c.1144C>G locus showed moderate polymorphism, suggesting that it could be used for breeding selection. Furthermore, both loci were in Hardy-Weinberg equilibrium, indicating that allele and genotype frequencies are stable in the population. This study also determined the association of c.723C>T locus and c.1144C>G locus with SCS in Chinese Holstein cows. Notably, a significant correlation between the c.723C>T locus and the mean value of SCS. The mean value of SCS of type AA was significantly lower than that of type AB at the c.723C>T polymorphic locus. These findings suggest that the c.723C>T locus may be a candidate gene associated with mastitis resistance. At the c.1144C>G locus, there was a difference between the SCS of the different genotypes, but it was not significant. Our findings are consistent with those of previous findings that have demonstrated the presence of a novel mutation at 1139 bp in exon 11 of the SLC11A1 gene and a significant association of this SNP with clinical mastitis [26]. In addition, the SNP in the SLC11A1 gene was identified as an important genetic marker and susceptibility factor for mastitis tolerance/susceptibility in Holstein and Swiss Brown cows [25]. It is suggested that SLC11A1 may be a candidate gene associated with mastitis resistance.
Meanwhile, in the SNP site analysis, we found that the c.723C>T site and the c.1144C>G site had alanine replaced by valine and proline replaced by alanine, respectively. These mutations are closely related to the function and stability of the SLC11A1 protein. It has been shown that mutations in SNPs lead to changes in the secondary structure of mRNAs and the tertiary structure of proteins, which in turn affects animal phenotypic traits [40,41]. To elucidate the effects of these sense mutations on the function of the SLC11A1 protein, we performed a series of bioinformatic analyses. The results of secondary structure prediction showed that the minimum free energy of the c.723C>T and c.1144C>G sites was increased compared to that before the mutation, leading to a decrease in the stability of the secondary structure of the mRNA, which may affect the efficiency of gene expression. The c.723C>T and c.1144C>G sites alter the proportion of proteins with secondary structure to varying degrees, which may affect the stability of the protein’s secondary structure, thereby affecting the protein’s tertiary structure and causing changes in protein function. Therefore, changes in the structure of SLC11A1 may influence mastitis resistance traits in Holstein cows.
However, the indirect role of SLC11A1 in mastitis needs to be further investigated. Most physiological processes are supported by multiple proteins, so building protein interaction networks can improve our understanding of complex traits such as mastitis. We predicted a protein interaction network involving SLC11A1 interacting with 10 proteins, of which SPI1, NOD2, TLR2 and S100A12 have been reported as candidate genes associated with mastitis resistance. SPI1 is an important member of the ETS transcription factor family and plays a key role in several steps of the inflammatory pathway. The SPI1 gene was found to be a candidate for immunity and mammary gland inflammation in dairy cows [6]. NOD2 is a key mediator of the innate and adaptive immune response to microbial infection. Association analysis of polymorphic loci of the NOD2 with mastitis in dairy cows showed significant differences in the allele frequencies of the polymorphic loci [42,43]. TLR2 plays a critical role in bacterial recognition and the host immune response during infection. TLR2 is not only associated with inflammation and immunity in dairy mammary tissue [44], but also plays a key role in apoptosis and angiogenesis in mammary tissue [45]. S100A12 protein triggers a signal transduction cascade response leading to activation of the transcriptional co-regulator NF-KB and subsequent expression of pro-inflammatory cytokines. S100A12 has been shown to enhance the antimicrobial capacity of dairy cows, helping to clear mammary tissue of infection [46], and can be used as a diagnostic indicator of subclinical mastitis in dairy cows [47]. These studies suggested that the SLC11A1 was at the core of candidate regulatory genes for mastitis and that the expression of this gene potentially influences mastitis production. Therefore, it was important to analyze the association of SLC11A1 polymorphisms with somatic cell scores.
5. Conclusions
In this study, we identified two mutations c.723C>T and c.1144C>G in the coding region of SLC11A1 in Chinese Holstein cows. Association analysis showed that the c.723C>T locus of the SLC11A1 gene had a significant effect on SCS. Meanwhile, at the c.723C>T and the c.1144C>G sites, alanine is replaced by valine and proline by alanine, respectively, and both mutations cause changes in the secondary and tertiary structure of the protein. Thus, SLC11A1 may be a candidate genes associated with mastitis resistance. This study provided a theoretical basis for resolving the important role of SLC11A1 in mastitis in dairy cows. This study focused on only two specific mutation sites in the gene; there may be other polymorphisms that also affect mastitis resistance, and functional experiments are needed to directly confirm the effects of these genetic variants on the expression and function of the gene. Future studies need to increase the sample size and further analyze the function of the two identified SNPs.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Sumon S. Parvin M.S. Ehsan M.A. Islam M.T. Relationship between somatic cell counts and subclinical mastitis in lactating dairy cows Vet. World 2020131709171310.14202/vetworld.2020.1709-171333061248 PMC 7522932 · doi ↗ · pubmed ↗
- 2Touza-Otero L. Landin M. Diaz-Rodriguez P. Fighting antibiotic resistance in the local management of bovine mastitis Biomed. Pharmacother.202417011596710.1016/j.biopha.2023.11596738043445 · doi ↗ · pubmed ↗
- 3Jaglan K. Ravikumar D. Sukhija N. George L. Alex R. Vohra V. Verma A. Genomic clues of association between clinical mastitis and SN Ps identified by dd RAD sequencing in Murrah buffaloes Anim. Biotechnol.2023344538454610.1080/10495398.2023.216593736639144 · doi ↗ · pubmed ↗
- 4Rupp R. Boichard D. Genetic parameters for clinical mastitis, somatic cell score, production, udder type traits, and milking ease in first lactation Holsteins J. Dairy Sci.1999822198220410.3168/jds.S 0022-0302(99)75465-210531607 · doi ↗ · pubmed ↗
- 5Chen R. Wang Z. Yang Z. Zhu X. Ji D. Mao Y. Association of IL 8 -105G/A with mastitis somatic cell score in Chinese Holstein dairy cows Anim. Biotechnol.20152614314710.1080/10495398.2014.93965725380467 · doi ↗ · pubmed ↗
- 6Jiang Q. Zhao H. Li R. Zhang Y. Liu Y. Wang J. Wang X. Ju Z. Liu W. Hou M. In silico genome-wide mi RNA-QTL-SN Ps analyses identify a functional SNP associated with mastitis in Holsteins BMC Genet.2019204610.1186/s 12863-019-0749-531096910 PMC 6524300 · doi ↗ · pubmed ↗
- 7Xu Y. Zhang X. Hui T. Sun J. Cai W. Lin G. Wang L. Dou X. Wang Z. Han D. Association analysis for SN Ps of KRT 26 and TCHH genes with cashmere production performance, body measurement traits and milk production traits in Liaoning cashmere goats Anim. Biotechnol.20233469870810.1080/10495398.2021.199638634747683 · doi ↗ · pubmed ↗
- 8An D. Chen X. Li Z. Dai L. Huang J. Xiao M. Liu H. Xu J. Ruan Y. Genetic variation in the BLM gene and its expression in the ovaries is closely related to kidding number in goats Theriogenology 202421825426610.1016/j.theriogenology.2024.02.01338367334 · doi ↗ · pubmed ↗