A Highly Potent Apomorphine Derivative Enhancing Neurite Outgrowth via Nrf2 Activation
Tamaki Ishima, Hitoshi Osaka, Ryozo Nagai, Kenichi Aizawa

TL;DR
A new apomorphine derivative, D55, was found to strongly promote nerve growth through Nrf2 activation, offering potential for treating neurodegenerative diseases.
Contribution
The study introduces D55, a novel apomorphine derivative with high potency for Nrf2 activation and neurite outgrowth.
Findings
D55 showed the highest potency (EC50 = 0.5661 nM) in promoting neurite outgrowth.
Neuroplasticity effects of APO, D55, and Eda were Nrf2-dependent, as shown by siRNA knockdown.
D30, with a methoxy substitution, had no effect on neurite outgrowth.
Abstract
Apomorphine (APO), a dopamine agonist, activates nuclear factor erythroid 2-related factor 2 (Nrf2) and exerts antioxidant effects, making it a promising candidate for neuroprotection against oxidative stress. This study evaluated neuroplasticity-enhancing properties of newly synthesized APO derivatives, focusing on their ability to promote neurite outgrowth in PC12 cells under nerve growth factor (NGF) stimulation. D55, an APO derivative, retains the hydroxyl group at APO’s 11th position while substituting the 10th with an ethoxy group. D55 exhibited the highest potency (EC50 = 0.5661 nM), significantly enhancing neurite outgrowth. APO demonstrated the highest efficacy (Emax ~10-fold increase), while edaravone (Eda) required higher concentrations (EC50 = 22.5 nM) for moderate effects (Emax ~4-fold increase). D30, in which the 11th hydroxyl was replaced with a methoxy group, had no…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
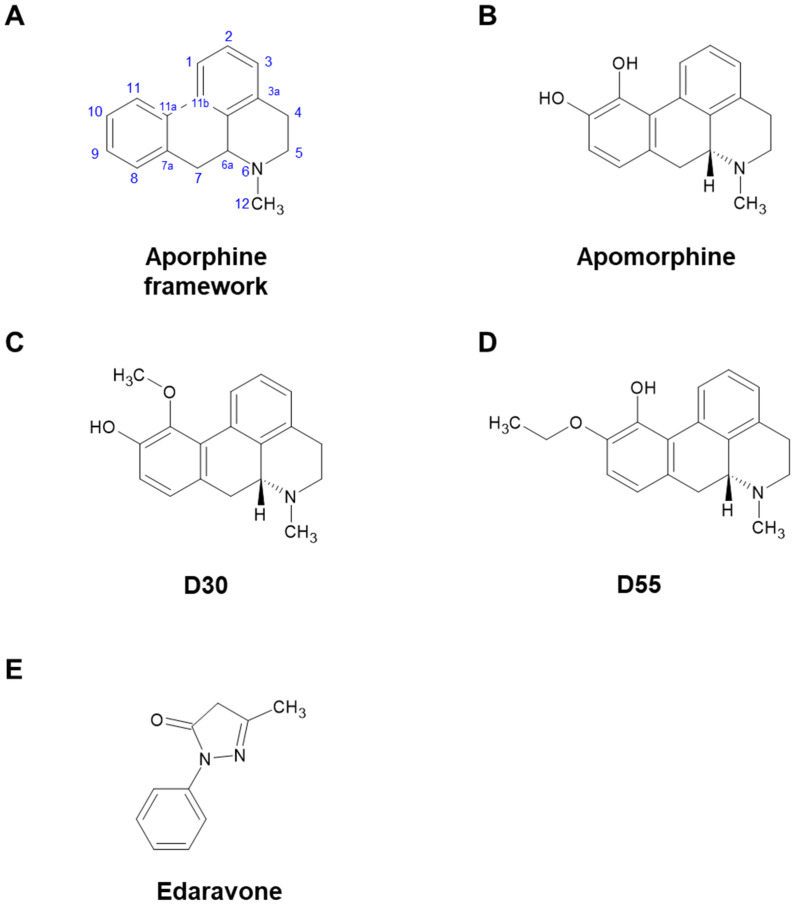 Figure 1
Figure 1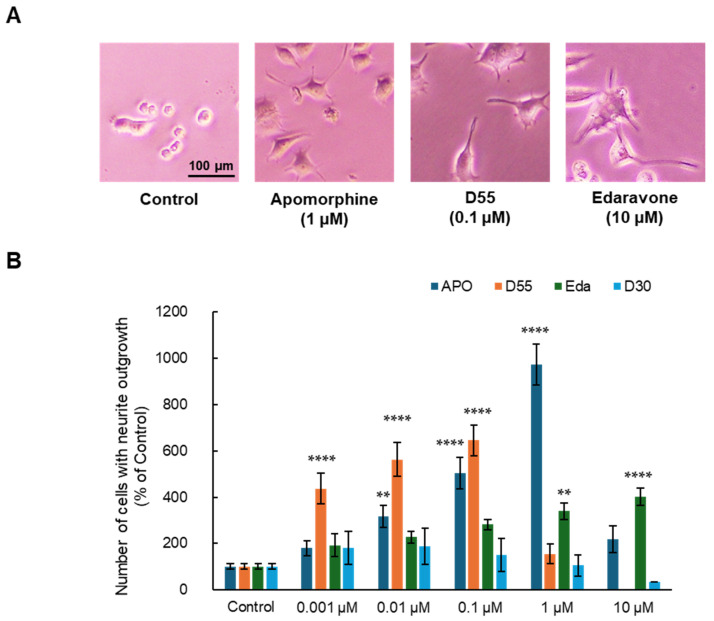 Figure 2
Figure 2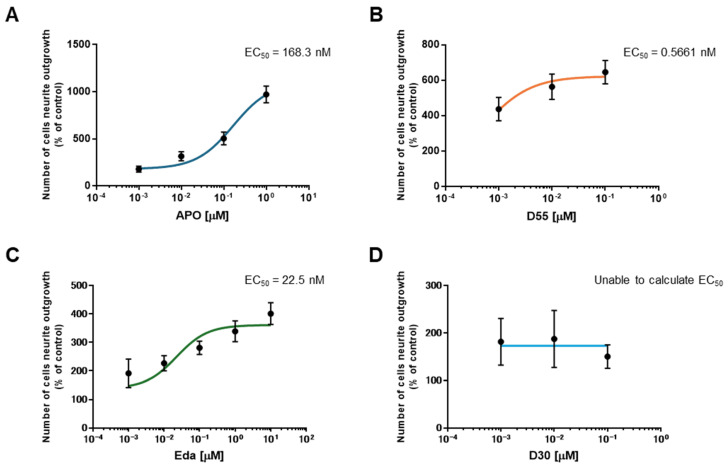 Figure 3
Figure 3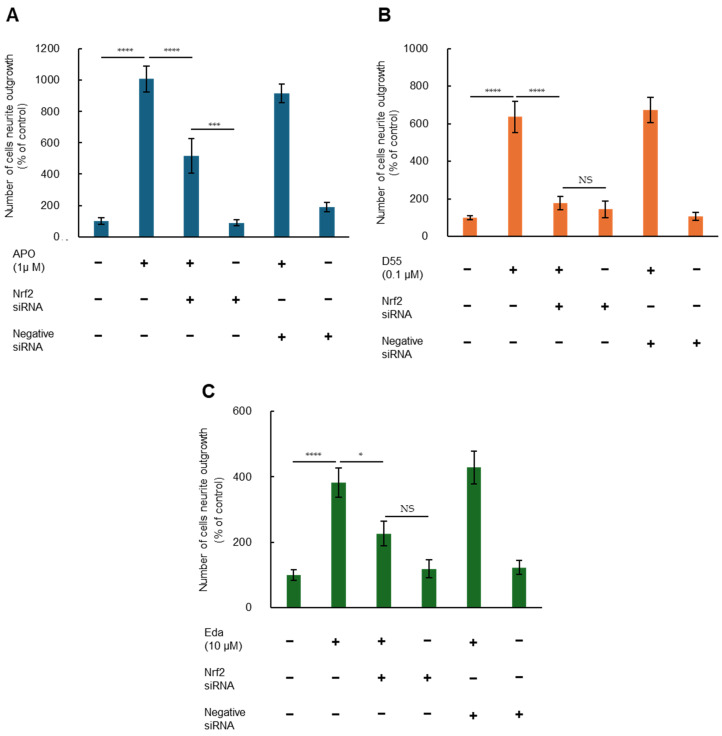 Figure 4
Figure 4- —Japan Agency for Medical Research and Development (AMED)
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsCholinesterase and Neurodegenerative Diseases · Genomics, phytochemicals, and oxidative stress · Piperaceae Chemical and Biological Studies
1. Introduction
Apomorphine (APO), a drug used to treat Parkinson’s disease and other disorders, is primarily known as a dopaminergic (DA) agonist. However, it also possesses many other potentially useful properties [1,2]. Miyauchi et al. identified APO as an agent that rescues apoptosis of fibroblasts induced by reactive oxygen species in pediatric patients with mitochondrial encephalomyopathies [3]. However, in order to reposition APO as a therapeutic agent for mitochondrial diseases, the strong DA effect leading to emesis, an undesirable side effect, became a challenge. Therefore, Kobayashi et al. synthesized 20 derivatives with an aporphine framework (Figure 1A) and screened their DA activity and anti-cell death effect under oxidative stress. As a result, from the parent compound, APO (Figure 1B, IUPAC: 6-Methyl-5,6,6a,7-tetrahydro-4H dibenzo[de,g]quinoline-10,11-diol), we obtained D30 (Figure 1C, IUPAC: 11-Methoxy-6-methyl-5,6,6a,7-tetrahydro-4H dibenzo[de,g]quinolin-10-ol) and D55 (Figure 1D, IUPAC: 10-Ethoxy-6-methyl-5,6,6a,7-tetrahydro-4H dibenzo[de,g]quinolin-11-ol). D30 lost its anti-cell death effect, but D55 showed better anti-cell death effect (EC_50_ = 4 nM) than APO [4]. It has been suggested that the anti-cell death effect of APO involves the nuclear factor erythroid 2-related like 2 (Nrf2) Antioxidant Response Element (ARE) pathway [5]. Recently, in a study using a model of steatohepatitis, Maeda et al. reported that APO activates Nrf2 and inhibits ferroptosis [6]. In studies of ischemic stroke and ALS models, edaravone (Eda) (Figure 1E), a cerebroprotective agent, was also approved for treatment of amyotrophic lateral sclerosis (ALS) and has been reported as an Nrf2 activator [7,8,9].
It has been reported that Nrf2 is involved in neuroprotection, including neuroplasticity [10]. The PC12 cell line, derived from rat adrenal medullary pheochromocytoma, has been used to study neuronal plasticity because it differentiates into neuron-like cells and promotes neurite outgrowth in response to drugs such as nerve growth factor (NGF) [11,12,13]. However, in studies involving APO, there are only reports of its use to measure cytotoxicity in Parkinsonian models [14], and to our knowledge, there are no reports of its effects on neuroplasticity. In this study, we first determined whether these compounds induce NGF-mediated neurite outgrowth in PC12 cells and then calculated their potency. Furthermore, the role of Nrf2 in neuroplasticity was verified through RNAi experiments.
2. Materials and Methods
2.1. Compounds
Compounds were obtained from the following sources: (R)-(-)-Apomorphine Hydrochloride Hemihydrate was sourced from Fujifilm Wako Pure Chemicals Corporation (#013-18323, Osaka, Japan), and 3-Methyl-1-phenyl-5-pyrazolone (Eda) from Tokyo Chemical Industry Co., Ltd. (#M0687, Tokyo, Japan). D30 (CAS No. 36507-69-4) and D55 (CAS No. 2926569-76-6) were derived from apomorphine as the parent compound [4].
2.2. Quantification of Neurite Outgrowth in PC12 Cells
PC12 cells (RIKEN Cell Bank, Tsukuba, Japan) were cultured at 37 °C and 5% CO_2_ in Dulbecco’s Modified Eagle’s Medium (DMEM), supplemented with 10% heat-inactivated fetal bovine serum (FBS), 5% heat-inactivated horse serum, and 1% Gibco™ penicillin–streptomycin (10,000 U/mL: Thermo Fisher Scientific, Waltham, MA, USA). The medium was changed two to three times a week. PC12 cells were plated onto 24-well tissue culture plates coated with poly-D-lysine/laminin. Cells were plated at a relatively low density (0.25 × 10^4^ cells/cm^2^) in DMEM medium containing 0.5% FBS and 1% penicillin–streptomycin. Medium containing a minimal level of serum (0.5% FBS) was used as previously reported [15,16,17]. Twenty-four hours after plating, the medium was replaced with DMEM medium containing 0.5% FBS and 1% penicillin–streptomycin with NGF (2.5 ng/mL), with or without 0.001, 0.01, 0.1, 1, or 10 μM of each compound.
Four days after incubation with NGF (2.5 ng/mL) with or without the specified compound, morphometric analysis was performed on digitized images of live cells taken under phase contrast illumination using an inverted microscope linked to a camera. An average of 100 cells per field was analyzed. Differentiated cells were counted by visual examination of the field. Only cells with at least one neurite were counted and expressed as a percentage of all cells in the field. Counting was performed in a blinded manner. Data were expressed as a percentage of the control group (NGF alone).
RNAi gene expression knockdown studies were performed according to previously reported methods [18,19], using the following sequence: Rattus norvegicus Nrf2 (Nfe2l2), mRNA GenBank Accession No. NM_031789: sense, 5′-GCCAAAGCUAGUGUAGAAAAUAUAT-3′; antisense, 5′-AUAUUAUUCUACACUAGCUUGGCUA-3′. For all relative control experiments, cells were exposed to scrambled non-specific RNAi duplex with the following sequence: sense, 5′-CUCCUUCUCUCCCUUGUGA-3′; antisense, 5′-UCACAAGGGAGAAAGAGAGGAAGGA-3′. Each RNAi duplex was synthesized as chimeras, incorporating two DNA nucleotides in the 25-mer sense strand to enhance RNAi efficacy. These were then annealed with the 27-mer antisense strand (FASMAC Co., Ltd., Atsugi, Japan) and introduced into cells using Lipofectamine 2000 reagent (Invitrogen, Carlsbad, CA, USA) following the manufacturer’s guidelines.
2.3. Statistical Analysis
Statistical analysis was performed using GraphPad Prism (version 7.00, GraphPad Software, Boston, MA, USA). Data were analyzed by one-way analysis of variance (ANOVA) or two-way ANOVA. As appropriate, post hoc comparisons were performed using the Bonferroni test or Tukey’s Honestly Significant Difference test. Data are presented as means ± standard errors of the mean (S.E.M.), with statistical significance set at p < 0.05.
3. Results
3.1. Effects of APO, Its Derivatives, and Eda on NGF-Induced Neurite Outgrowth in PC12 Cells
Effects of each compound on NGF-induced neurite outgrowth in PC12 cells were examined. DMEM medium containing the minimum amount of FBS (0.5%) in which cells could survive and 2.5 ng/mL NGF, the minimum requirement for neuronal differentiation, was used as a control.
As a result, APO, D55, and Eda exhibited neurite outgrowth effects at concentrations of 1 µM, 0.1 µM, and 10 µM, respectively (Figure 2A). APO, D55, and Eda exhibited concentration-dependent neurite outgrowth effects at concentrations up to 1 µM, 0.1 µM, and 10 µM, respectively, while D30 had no effect (Figure 2B). APO exhibited an Emax of approximately 10-fold neurite outgrowth compared to the control at 1 µM, whereas D55 exhibited an Emax of about 4-fold neurite outgrowth compared to the control at concentrations as low as 1 nM. Eda showed no toxicity at concentrations as high as 10 µM.
3.2. EC50 of APO, Its Derivatives, and Eda
To evaluate the potency of each drug, the half-maximal effective concentration (EC_50_) was calculated. EC_50_ values for APO, D55, and Eda were 168.3 nM, 0.5661 nM, and 22.5 nM, respectively (Figure 3A–C). For D30, the EC_50_ could not be calculated as it does not promote neurite outgrowth (Figure 3D).
3.3. Role of Nrf2 in Potentiating NGF-Induced Neurite Outgrowth by APO, D55, and Eda
To investigate whether Nrf2 is involved in potentiation of NGF-induced neurite outgrowth by APO, D55, and Eda, we examined effects of Nrf2 gene knockdown. Two-way ANOVA of neurite outgrowth data showed significant effects, indicating that potentiation effects of APO (1.0 µM), D55 (0.1 µM), and Eda (10 µM) on NGF-induced neurite outgrowth were significantly attenuated by treatment with Nrf2 siRNA, but not by the negative control (Figure 4). In contrast, treatment with either Nrf2 siRNA or the negative control alone did not alter NGF-induced neurite outgrowth in PC12 cells (Figure 4). These findings suggest that APO, D55, and Eda enhance NGF-induced neurite outgrowth through activation of Nrf2. However, in the case of APO, attenuation of the neurite outgrowth effect by Nrf2 siRNA treatment was partial (Figure 4A).
4. Discussion
4.1. Neuroprotective Effect of APO
A previous study by Miyauchi et al. screened apomorphine (APO) as a candidate drug for pediatric mitochondrial disease [3]. APO is a drug to improve off symptoms of Parkinson’s disease that reportedly has neuroprotective properties as well [20,21,22,23]. Recently, its neuroprotective effects due to its antioxidant properties [24], its anti-ferroptosis effects on fibroblasts [25], and its ability to reduce fatty liver and inflammation in a metabolic dysfunction-associated steatohepatitis (MASH) animal model [26] have been reported. It is thought to act as a radical scavenger [27] through activation of the Nrf2-ARE pathway [5].
4.2. Cell-Based In Vitro Model Using PC12 Cells
The PC12 pheochromocytoma cell line, derived from rat adrenal tumors, was established in 1976 [28]. Treatment of these cells with NGF halts their proliferation and promotes neurite outgrowth. Although not neurons, they have been used as in vitro neuronal-like models for studies of NGF signaling and neuronal differentiation [29,30,31], as well as models of cytotoxicity, including serum deprivation [32], toxic molecules [33], and neurodegenerative diseases [34,35]. Additionally, they are often used as in vitro models to identify neuroprotective agents. In this study, we investigated neurite outgrowth effects of APO and its derivatives, as well as Eda [36], which was used as a positive control for its radical scavenger properties.
As a result, concentration-dependent neurite outgrowth effects were observed with APO, its derivative D55, and Eda. APO has hydroxyl groups at positions 10 and 11 of the aporphine framework (Figure 1A,B). D30, which has a methoxy group at position 11 and lacks an anti-cell death effect, did not exhibit neurite outgrowth (Figure 1C). D55, which retains the hydroxyl group at position 11 (considered to have anti-cell death effects) and possesses an ethoxy group at position 10, showed similar effects (Figure 1D) [4]. Preliminary studies indicated that Eda did not induce cell death in PC12 cells even at high concentrations (100 µM). However, in this study, APO induced partial cell death at 10 µM, as did D55 at concentrations above 1 µM. At 1 µM, APO exhibited an Emax about ten-fold more effective than the control in promoting neurite outgrowth, whereas D55 exhibited an Emax four-fold more effective than the control, even at a concentration of 1 nM.
The EC_50_ of each drug was calculated, with D55 having the lowest value at 0.5661 nM. APO causes neurotoxicity and neurodegeneration in animal models at high concentrations [37,38]. It also induces apoptosis in choriocarcinoma at concentrations of 10 µM or higher [39]. D55, which is potent at low concentrations, may have an advantage over APO and Eda in terms of reducing the risk of adverse effects.
4.3. Neurite Outgrowth and Nrf2
It has already been reported that the phytochemical, sulforaphane, in broccoli sprouts [18], Astragaloside IV in dried roots of Astragalus membranaceus (Radix Astragali) [40], and Nrf2 activators, such as TBE-31 and MCE-1 [19], enhance neurite outgrowth via Nrf2-mediated pathways. Although there are reports that both APO and Eda activate Nrf2 [6,41,42], there have been no reports on their effects on neurite outgrowth. In the present study, we sought to determine whether Nrf2 is associated with neurite outgrowth in PC12 cells and whether this is also the case for D55. Potentiating effects of APO, D55, or Eda on NGF-induced neurite outgrowth were significantly antagonized by Nrf2 siRNA treatment, but not by negative siRNA. Nrf2 siRNA and negative siRNA alone did not alter NGF-induced neurite outgrowth. Thus, these neurite outgrowth effects proved to be associated with Nrf2 expression.
Regarding APO, inhibition of neurite outgrowth effects by Nrf2 siRNA was partial (Figure 4A). Studies on atypical antipsychotic drugs, aripiprazole and brexpiprazole, have reported that the selective 5-hydroxytryptamine (5-HT)1A receptor antagonist, WAY100635, inhibits neurite outgrowth, whereas the agonist 8OH-DPAT has neurite outgrowth-promoting effects. Additionally, the selective 5-HT_2A_ receptor agonist, DOI, does not have neurite outgrowth-promoting effects, whereas the antagonist, M100907, promotes neurite outgrowth [16,17]. Since APO acts as a partial agonist of 5-HT_1A_ receptors and an antagonist of 5-HT_2A_ receptors [1,43], it is possible that some effects not inhibited by Nrf2 siRNA are related to serotonin receptor activity.
4.4. Neuroplasticity
There are many reports that Nrf2 is involved in neuroprotection [44]; however, in cellular models, the only reports on Nrf2 activation so far are related to neuroprotection under oxidative stress or against cytotoxicity, as shown in cell viability studies [45]. Even in studies limited to PC12 cells, research with APO has only reported neuroprotection against MPTP and 6-hydroxydopamine [46]. Neurite outgrowth observed with PC12 cells in the present study demonstrated the neurite outgrowth effect of the drug itself, independent of neuroprotection. These findings indicate that APO, D55, and Eda are associated with neuroplasticity. This is the first report on neuroplasticity of APO and its new derivatives, D55 and Eda.
Nrf2 is a modulator of proteostasis and has been implicated in Parkinson’s disease [47,48] and other neurodegenerative diseases [49] such as Alzheimer’s disease [50,51], Huntington’s disease [52,53], ALS [54,55], and ataxia [56,57]. It is also associated with psychiatric disorders such as depression [58,59] and schizophrenia [60,61], as well as stroke [62,63], traumatic brain injury [64,65], and multiple sclerosis [66,67]. D55, an Nrf2 activator that is unlikely to have DA adverse effects, appears to be a promising compound for treating neurological and psychiatric disorders, in addition to mitochondrial encephalomyopathies, due to its neuroprotective and neuroplasticity effects.
4.5. Limitations
The primary limitation of this study is that it was conducted solely using PC12 cells. Since Nrf2 is essential not only in oxidative stress, but also in proteostasis, detailed analyses need to be conducted using KO mice and other models. Additionally, further investigation is required to understand the effects of D55 beyond the D2 receptor. Detailed mechanisms of neuroplasticity remain unknown. According to Tebey et al., an excess of Nrf2 may disrupt the balance of homeostasis toward by causing reductive stress, which promotes protein misfolding [68]. It is believed that low levels of Nrf2 in the brain require only slight upregulation to be beneficial under pathological conditions [49]. Despite challenges in practical applications, we think that drugs like D55, which promote neurite outgrowth at low concentrations, may be advantageous in this regard.
Furthermore, in the future, we plan to (i) investigate other signaling pathways involved in neurite outgrowth, beyond Nrf2, (ii) perform long-term viability studies and LD_50_ assessments to evaluate safety margins of these compounds, (iii) assess additional neurite outgrowth markers such as β-III tubulin, GAP-43, and PSD-95, and (iv) test other neurotrophic factors and Nrf2 activators/inhibitors beyond NGF, to further validate neuroplasticity-enhancing effects of these compounds.
5. Conclusions
D55 exhibits NGF-induced neurite outgrowth effects in PC12 cells and is more effective at lower concentrations than the parent compound, APO, or the free radical scavenger, Eda. These results indicate that D55 not only prevents cell death but also has neuroplastic properties. We confirmed that this effect is mediated by Nrf2. Drugs that promote Nrf2-mediated neuroplasticity without the side effects of D55 are expected to be useful in the treatment of neurological and psychiatric disorders.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Auffret M. Drapier S. Vérin M. Pharmacological Insights into the Use of Apomorphine in Parkinson’s Disease: Clinical Relevance Clin. Drug Investig.20183828731210.1007/s 40261-018-0619-329327219 · doi ↗ · pubmed ↗
- 2RibaričS. The pharmacological properties and therapeutic use of apomorphine Molecules 2012175289530910.3390/molecules 1705528922565480 PMC 6268166 · doi ↗ · pubmed ↗
- 3Miyauchi A. Kouga T. Jimbo E.F. Matsuhashi T. Abe T. Yamagata T. Osaka H. Apomorphine rescues reactive oxygen species-induced apoptosis of fibroblasts with mitochondrial disease Mitochondrion 20194911112010.1016/j.mito.2019.07.00631356884 · doi ↗ · pubmed ↗
- 4Kobayashi M. Miyauchi A. Jimbo E.F. Oishi N. Aoki S. Watanabe M. Yoshikawa Y. Akiyama Y. Yamagata T. Osaka H. Synthetic aporphine alkaloids are potential therapeutics for Leigh syndrome Sci. Rep.2024141156110.1038/s 41598-024-62445-w 38773300 PMC 11109252 · doi ↗ · pubmed ↗
- 5Hara H. Ohta M. Adachi T. Apomorphine protects against 6-hydroxydopamine-induced neuronal cell death through activation of the Nrf 2-ARE pathway J. Neurosci. Res.20068486086610.1002/jnr.2097416802348 · doi ↗ · pubmed ↗
- 6Maeda H. Miura K. Aizawa K. Bat-Erdene O. Sashikawa-Kimura M. Noguchi E. Watanabe M. Yamada N. Osaka H. Morimoto N. Apomorphine Suppresses the Progression of Steatohepatitis by Inhibiting Ferroptosis Antioxidants 20241380510.3390/antiox 1307080539061874 PMC 11273851 · doi ↗ · pubmed ↗
- 7Barati A. Moghimi S. Taghavi Zanjani K. Rohani M. Sohrabi Hesar M. Arfaie A. Ghezelche Khamsiyan M. Mahmoudi J. Sadigh-Eteghad S. Acute Administration of Edaravone Improves Cognitive Impairment in a Mouse Model of m PFC Ischemia: Crosstalk Between Necroptosis, Neuroinflammation, and Antioxidant Defense Mol. Neurobiol.2025624420443410.1007/s 12035-024-04541-639448519 · doi ↗ · pubmed ↗
- 8Bajpai A. Bharathi V. Patel B.K. Therapeutic targeting of the oxidative stress generated by pathological molecular pathways in the neurodegenerative diseases, ALS and Huntington’s Eur. J. Pharmacol.202598717718710.1016/j.ejphar.2024.17718739645221 · doi ↗ · pubmed ↗