Antibody-guided identification of Achromobacter xylosoxidans protein antigens in cystic fibrosis
Cecilia Sahl, Sounak Chowdhury, Johan Malmström, Lisa I. Påhlman

TL;DR
This study identifies specific Achromobacter xylosoxidans proteins that trigger antibody responses in cystic fibrosis patients, which could help in diagnosing infections.
Contribution
The study introduces a novel method to identify A. xylosoxidans antigens using patient antibodies, revealing potential biomarkers for infection.
Findings
Four out of seven Achromobacter-infected patients had antibodies against the bacteria.
Eight antigens were identified, with three (DLD, type I secretion domain, DUF336) showing specific IgG responses.
DLD and DUF336 showed minimal binding to sera from non-infected cystic fibrosis patients.
Abstract
Persistent bacterial airway infection is a hallmark feature of cystic fibrosis (CF). Achromobacter spp. are gram-negative rods that can cause persistent airway infection in people with CF (pwCF), but the knowledge of host immune responses to these bacteria is limited. The aim of this study was to investigate if patients develop antibodies against Achromobacter xylosoxidans, the most common Achromobacter species, and to identify the bacterial antigens that induce specific IgG responses. Seven serum samples from pwCF with Achromobacter infection were screened for antibodies against bacteria in an ELISA coated with A. xylosoxidans, A. insuavis, or Pseudomonas aeruginosa. Sera from pwCF with or without P. aeruginosa infection (n = 22 and 20, respectively) and healthy donors (n = 4) were included for comparison. Serum with high titers to A. xylosoxidans was selected for affinity purification…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
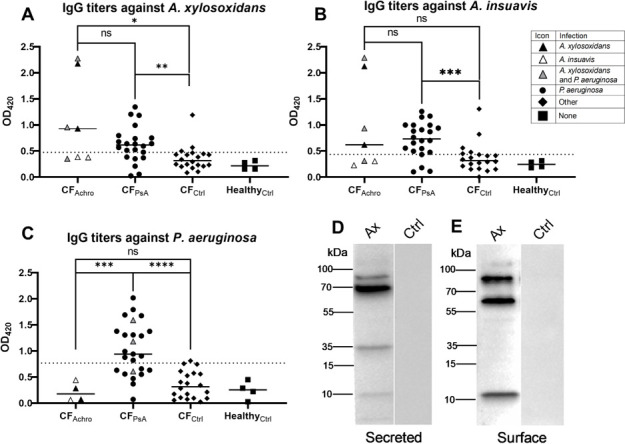 Fig 1
Fig 1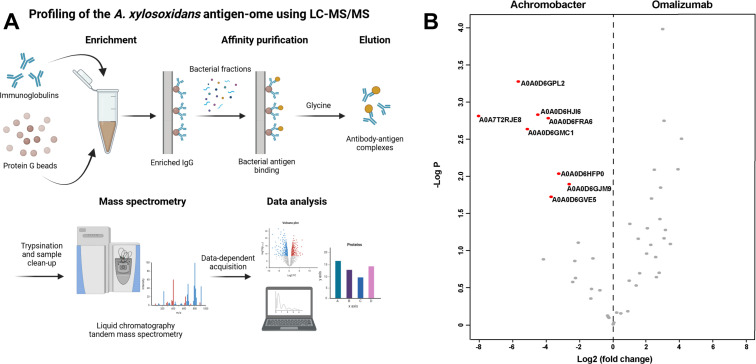 Fig 2
Fig 2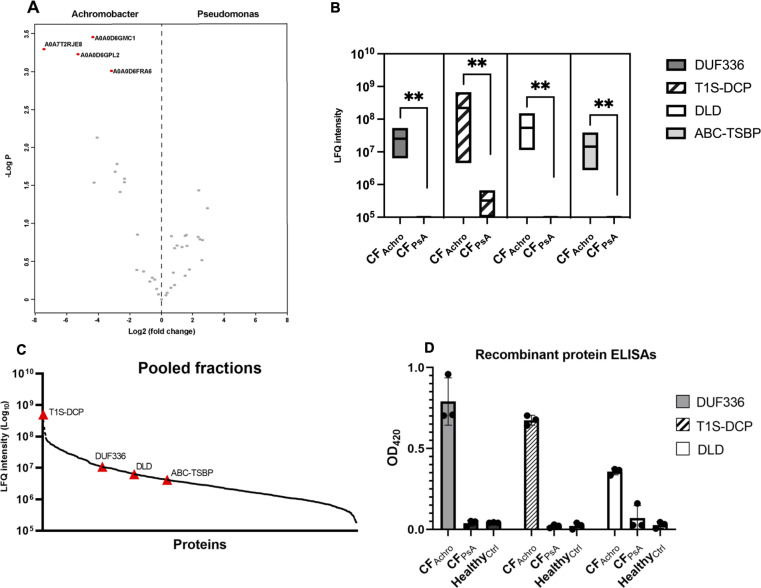 Fig 3
Fig 3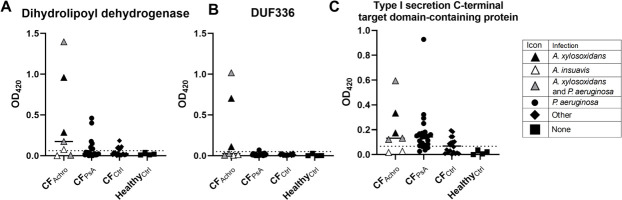 Fig 4
Fig 4| UniProt ID | Name | Fraction | Function |
|---|---|---|---|
|
| Type I secretion C-terminal target domain-containing protein | Secreted, surface | Membrane transport |
|
| Dihydrolipoyl dehydrogenase | Secreted, surface | Dehydrogenase activity |
|
| Domain of uncharacterized function (DUF336) | Secreted, surface | Heme binding |
|
| DNA protection during starvation protein 2 | Surface | Iron binding |
|
| Amino acid ABC transporter substrate-binding protein | Secreted, surface | Membrane transport |
|
| Inorganic pyrophosphatase | Secreted, surface | Phosphatase activity |
|
| Glutamate dehydrogenase | Secreted, surface | Dehydrogenase activity |
|
| Alkyl hydroperoxide reductase AhpD | Secreted, surface | Hydroperoxidase activity |
| Positive samples | CFAchro ( | CFPsA ( | CFCtrl ( | ||
|---|---|---|---|---|---|
| DLD | 5 (71%) | 7 (32%) | 0.06 | 6 (30%) | 0.05 |
| DUF336 | 3 (43%) | 1 (5%) | 0.01 | 0 (0%) | 0.001 |
| T1S-DCP | 5 (71%) | 18 (82%) | 0.55 | 10 (50%) | 0.33 |
- —Hjärt-Lungfondenhttp://dx.doi.org/10.13039/501100003793
- —Alfred Österlunds Stiftelsehttp://dx.doi.org/10.13039/501100005390
- —Stiftelsen Riksförbundet Cystisk Fibros Forskningsfondhttp://dx.doi.org/10.13039/501100009235
- —Swedish government funds for clinical research
- —Vetenskapsrådethttp://dx.doi.org/10.13039/501100004359
- —Knut och Alice Wallenbergs Stiftelsehttp://dx.doi.org/10.13039/501100004063
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsInfections and bacterial resistance · Cystic Fibrosis Research Advances · Glycosylation and Glycoproteins Research
INTRODUCTION
Cystic fibrosis (CF) is a monogenic disease caused by mutations in the CF transmembrane regulator (CFTR) gene. These mutations result in impaired chloride transport across cell membranes, leading to the accumulation of abnormally thick mucus in the airways and pancreatic ducts. The viscous CF mucus provides a favorable environment for bacterial infections. Persistent respiratory tract infections and the resulting airway inflammation cause progressive lung injury, ultimately leading to respiratory failure, which is the primary cause of mortality in this patient population (1).
Achromobacter spp. is a genus of environmental gram-negative bacteria capable of causing opportunistic airway infections in people with CF (pwCF) (2, 3). The prevalence of Achromobacter spp. infection in pwCF is approximately 5%, although the rate varies between countries (4). Achromobacter xylosoxidans is the most frequently identified Achromobacter species in pwCF, but other species, including A. insuavis, are also reported (5, 6). Persistent Achromobacter spp. infection is associated with decreased lung function (7), an increased need for intravenous antibiotics (8), more frequent pulmonary exacerbations, and a higher risk of requiring lung transplantation (9). However, it remains unclear whether Achromobacter actively contributes to these outcomes or if its presence is merely indicative of low lung function caused by other factors (3, 10).
While common CF pathogens such as Pseudomonas aeruginosa are well-characterized, the virulence mechanisms and host responses to Achromobacter are less understood. According to a previous study, pwCF infected with Achromobacter develop antibody responses against the bacteria (11). However, the specific antigenic targets are not yet identified. The aim of this study was to identify proteins expressed by Achromobacter that are recognised by host antibodies. To characterize Achromobacter antigens targeted by circulating IgGs in pwCF, we took advantage of a novel mass spectrometry-based systems antigenomics approach (12) where serum IgG from pwCF infected with Achromobacter spp. were used to affinity-purify bacterial antigens from the most common species of Achromobacter, that is, A. xylosoxidans.
MATERIALS AND METHODS
Human sera and bacterial isolates
Serum samples were collected from adult people with CF attending the CF center at Skåne University Hospital, Lund, Sweden. Individuals with repeated growth of Achromobacter spp. in airway cultures were regarded to have Achromobacter airway infection. Chronic P. aeruginosa infection was defined as growth in ≥50% of airway cultures according to Leeds criteria (13). Sera from healthy individuals were included as controls.
A. xylosoxidans type strain 56,438T and A. insuavis type strain 62,426T were purchased from the Culture Collection University of Gothenburg, Sweden. The P. aeruginosa reference strain PAO1 was a gift from Professor Arne Egesten, Lund University. Clinical Achromobacter spp. isolates were obtained from the Department of Clinical Microbiology at Skåne University Hospital, Lund, Sweden. Species identification of clinical Achromobacter isolates was done using standard laboratory methods at the Department of Clinical Microbiology at Skåne University Hospital, Lund, Sweden, followed by nrdA sequencing as described previously (14).
ELISA of serum IgG binding to whole bacteria
Three to four colonies of A. xylosoxidans, A. insuavis, or P. aeruginosa type strains were suspended in 20 mL coating buffer (1.69 g Na_2_CO_3_ and 2.94 g NaHCO_3_ in 1 L H_2_O, pH 9.6) and dispensed at a volume of 200 µL into a 96-well microtiter plate (Nunc Maxisorp). After incubation overnight at 37°C, 5% CO_2_, the coating buffer was removed, and bacteria were heat-killed at 80°C for 15 min. Unspecific binding was blocked by incubating with 5% BSA in PBST (blocking buffer) for one hour. Sera were diluted 1:100 in blocking buffer, added at a volume of 200 µL to the coated wells, and incubated for 1 h. The plates were then washed three times with PBST. Protein G-HRP conjugate (Bio-Rad #170-6425) was diluted 1:3,000 in blocking buffer and added at a volume of 200 µL. The plates were incubated for 1 h, after which the wash step was repeated. A total of 200 µL of developing solution (600 µL of 2% [wt/vol] ABTS and 248 µL 0.6% H_2_O_2_ in 12 mL of substrate buffer (0.1 M C_6_H_8_O_2_ × H_2_O) and 0.1 M Na_2_HPO_4_ × 2H_2_O, pH 4.5) was added and incubated in the dark for 5 min. The absorbance was read at 420 nm, and the background OD of blank wells was subtracted. The average OD value of the healthy control group + 3 standard deviations was considered the cut-off for positivity.
Preparation of bacterial secreted and surface proteins
A. xylosoxidans was grown overnight in ABT medium (15) supplemented with 0.5% (wt/vol) of casamino acids and 0.5% (wt/vol) glucose (Merck). The following day, a subculture with OD_620_ = 0.1 was started in the same medium and grown to the mid-logarithmic phase (OD_620_ ≈ 0.4) with shaking incubation at 37°C and 5% CO_2_. The resulting culture was transferred to 50 mL tubes and centrifuged at 10,000 × g, 30 min, at 4°C.
Secreted proteins were prepared from the supernatant. After sterilization using a 0.22 µm filter (Merck-Millipore #SLGP033RB), the supernatant was purified through 10 kDa ultrafiltration columns (Merck-Millipore #UFC901008) by centrifuging at 4,000 × g for 12 min at 4°C. The fraction collected in the filter was washed three times with ice-cold PBS and saved.
Surface proteins were prepared from the bacterial pellet. After washing two times in HEPES buffer, bacteria were resuspended in HEPES buffer at a concentration of approx. 1 × 10^8^ CFU. A total of 0.2 µg trypsin (Promega V5113) per mL of original bacterial culture was added and incubated for 60 min at 37°C, 500 rpm. The reaction was then stopped by incubating on ice for 2 min. The samples were centrifuged at 1,000 × g for 15 min at 4°C, and the supernatant was transferred into new tubes. The resulting surface protein fraction was incubated for 15 min at 85°C, 500 rpm to heat-kill any remaining bacteria.
Protein concentrations in both fractions were measured using the Pierce BCA Protein Assay Kit (Thermo Scientific, 23225) according to the manufacturer’s instructions, after which the fractions were saved at −20°C.
Western blot
Bacterial protein fractions were mixed 4:1 with 5× loading buffer (GenScript #MB01015) and denatured at 85°C for 10 min before loading onto an SDS-PAGE gel (Mini-PROTEAN TGX Precast, Bio-Rad) and run at 170 V for 1 h. The gel was blotted onto the membrane (Bio-Rad Trans-Blot Turbo #1704156) and blocked with 5% BSA in PBST for 1 h. The membrane was washed three times with PBST and incubated overnight at 4°C with 1:10,000 human serum in blocking buffer. After washing, the membrane was incubated with protein G-HRP conjugate diluted 1:3,000 in blocking buffer for 1 h at 37°C. The membrane was washed as described and then developed using Clarity Western ECL Substrate (Bio-Rad #1705061) according to the manufacturer’s instructions. Images were taken using the ChemiDoc (Bio-Rad).
Affinity purification of bacterial antigens
The affinity purification was performed essentially as described by Chowdhury et al. 12. Bacterial proteins were affinity purified using IgG from the following sources: one serum from pwCF infected with A. xylosoxidans (CF_Achro_), one serum from pwCF infected with P. aeruginosa (CF_PsA_), one healthy control serum, omalizumab (Xolair, Novartis), and pharmaceutical grade intravenous immunoglobulin G (IVIG, Octagam Octapharma). Omalizumab and IVIG were used at a concentration of 1 mg/mL, and the serum samples were diluted 1:10 to approximately match this concentration.
A total of 50 µL Pierce Protein G magnetic beads (ThermoFisher #13424229) was added to 1.5 mL microcentrifuge tubes. The beads were washed three times with 200 µL TBST using a magnetic rack. In total, 100 µL of each immunoglobulin sample and 400 µL TBST were added to the vials in triplicates and incubated with shaking at room temperature for 1 h. The beads were then washed with 500 µL TBST three times. The secreted and surface fractions of Achromobacter proteins were pooled so that 500 µL added to the beads contained 25 µg of each fraction, and the samples were then incubated at room temperature overnight. The beads were then washed three times with TBST before eluting the IgG-antigen complexes with 100 µL 0.1 M glycine. To neutralize the pH, 15 µL of 1 M Tris was added. Next, 15 µL of the eluted samples were run on SDS-PAGE as described above, and the gel was stained overnight with PageBlue solution (ThermoFisher #24620) to confirm the presence of antibody complexes. All affinity purifications were performed in triplicates.
Sample preparation for mass spectrometry and liquid chromatography-tandem mass spectrometry (LC-MS/MS) analysis
Samples obtained from affinity purifications as well as samples from both bacterial protein fractions were homogenized at 4,350 rpm for 90 s (BeadBug 6, Benchmark Scientific). Fifty microliters of each sample was denatured in 50 µL of ABC-urea buffer (8 M urea, 100 mM ammonium bicarbonate in HPLC-grade H_2_O). To reduce disulfide bonds, 0.5 µL of 500 mM Tris(2-carboxyethyl)phosphine was added, and the samples were incubated at 37°C for 1 h. Subsequently, 1 µL of 500 mM 2-iodoacetamide was added to alkylate the disulfide bonds, followed by a 30-min incubation at room temperature in the dark. Samples were incubated at 37°C overnight with 2 µL of 0.5 µg/µL trypsin. The reaction was then stopped by adding formic acid until pH 3.
Harvard apparatus Macro SpinColumns Silica C18 were used for sample purification according to the manufacturer’s instructions. The eluate was concentrated by SpeedVac (ThermoFisher) until complete dryness and resuspended in 20 µL 0.1% formic acid. Resuspended samples were sonicated for 5 min and transferred to MS vials, which were stored at −20°C until analysis.
The peptides were analyzed using a Q Exactive HFX instrument (Thermo Scientific) coupled to an Easy-nLC 1200 system (Thermo Scientific), in data-dependent acquisition (DDA) mode LC-MS/MS as described in Chowdhury et al. (12). Samples were picked up by the Easy-Spray LC column (50 cm, 45°C, Thermo Scientific) at a volume of 2 µL with 20 µL flow, followed by loading 6 µL with 5 µL/min flow. The maximum pressure was set to 800 bar. A linear gradient of 4–45% acetonitrile in 0.1% formic acid was run for 65 min. One full MS scan (resolution of 60,000 for a mass range of 390–1,210, automatic gain control = 3e6) was followed by MS/MS scans (resolution of 15,000, automatic gain control = 1e5) of the 15 most abundant signals. Isolation width was set to 2 m/z for precursor ions, and fragmentation was performed with higher-energy collisional-induced dissociation at a normalized collision energy [(N)CE] of 30.
Data analysis and statistics
The DDA data were analyzed in MaxQuant (16) (version 2.2.0.0) using the A. xylosoxidans reference proteome (Uniprot proteome ID: UP000595052) with the addition of common contaminants and RT peptides to the reference proteome file. Oxidation (M) and acetyl (protein N-term) were used as variable modifications. Carbamidomethyl (C) was used as a fixed modification. The maximum number of modifications per peptide was 5. Fast LFQ was used with a minimum ratio count of 2, a minimum of 2, and an average of 6 neighbours. Identification was performed with a false discovery rate (FDR) of 0.01 and matching between runs.
The data set obtained was further filtered in Perseus (17) (Cox Lab, version 2.0.7.0). Data were filtered to remove those identified by site, reverse, and potential contaminants. Hits with <2 unique peptides or present in <2 of the triplicates were excluded. After filtering and before statistical analysis, the LFQ intensities were log2 transformed and missing values replaced using imputed values from normal distribution. Proteins were considered antigenic hits if they were significantly enriched in the CF_Achro_ serum compared to the negative control omalizumab (double-sided t-test, FDR 0.05).
Recombinant protein expression and ELISA of serum IgG binding to recombinant proteins
The amino acid sequences of three putative antigens (dihydrolipoyl dehydrogenase [DLD; Uniprot identifier A0A0D6GPL2], type I secretion C-terminal target domain-containing protein [T1S-DCP; A0A7T2RJE8], and domain of uncharacterized function 336 [DUF336; A0A0D6GMC1]) were submitted to Protein Production Sweden for recombinant expression of proteins in Escherichia coli. Protein suspensions were diluted to a concentration of 4 µg/mL (T1s_DCP and DUF336) or 2 µg/mL (DLD) in coating buffer as described earlier. The coating buffer for the T1S protein was supplemented with 5 mM calcium chloride dihydrate. The ELISA was performed in triplicates as described above, except for the serum concentration being 1:50 and the development time for the colorimetric readout being 30 min.
PCR
Bacterial colonies were resuspended in PBS and placed in a heat block at 100°C for 5 min. Samples were spun down at 12,000 × g for 5 min and the resulting supernatants were used as PCR template in 1:10 dilution. Conventional PCR was performed with 1 U Taq-Polymerase “Phusion HS-II” (Thermo Scientific) and 0.5 µM primers (T1S-DCP forward CGAGGCCTACCTGAAGTTCTTT, T1S-DCP reverse CCATCGGTGTTGAGGGTGTC, DUF336 forward ACAAGTGGGCTGTCACCATC, DUF336 reverse CCGACAGGAAGGAATAGCGG, DLD forward TTCACCAAGCAAGGCCTGAA, DLD reverse CACGGCTTCCGAGATCAGTT) per 2 µL DNA sample. PCR products were visualized on 1% agarose (Thermo Scientific) using gel electrophoresis.
Local alignment search (basic local alignment search tool [BLAST])
BLAST in protein-protein mode was used to identify sequence similarity of A. xylosoxidans DLD, T1S-DCP, and DUF336 in comparison to A. insuavis (taxid 1287735) and P. aeruginosa (taxid: 287). The search was performed with the standard database of non-redundant protein sequences and scored with the BLOSUM62 matrix. The protein with the highest similarity in each comparison was selected and presented in File S3. Multiple sequence alignment was performed using CLUSTALW alignment with the weight matrix BLOSUM for proteins, and the alignment score was calculated by CLUSTALW from the number of identities between the two sequences divided by the length of the alignment.
Statistics
Enrichment analysis for antigenic hits was performed using volcano plots in Perseus 2.0.7.0 (double-sided t-test, FDR 0.05). Statistical calculations for ELISA experiments were performed using GraphPad Prism 10.0.2 (GraphPad Software, San Diego, CA, USA). Comparisons between groups were made using the non-parametric Mann-Whitney U-test for continuous variables. A P value ≤ 0.05 was considered statistically significant.
RESULTS
Achromobacter infection evokes circulating antibodies in pwCF
To evaluate IgG responses against Achromobacter spp., serum samples from seven pwCF with Achromobacter spp. airway infection (CF_Achro_) were screened for antibody titers against whole Achromobacter bacteria. Five of the seven individuals were infected with A. xylosoxidans and two with A. insuavis. Three of the patients were also chronically co-infected with P. aeruginosa. To quantify serum-IgG against whole bacteria, microtiter plates were coated with either A. xylosoxidans or A. insuavis, and serum IgG titers were then analyzed by ELISA. For comparison, serum samples from pwCF with chronic P. aeruginosa infection (CF_PsA_, n = 22), sera from pwCF controls without Achromobacter or P. aeruginosa infection (CF_Ctrl_, n = 20), and sera from healthy donors (n = 4) were included in the analysis. Four of the seven individuals infected with Achromobacter spp. (57%) had positive serum titers against both A. xylosoxidans and A. insuavis (Fig. 1A and B). All four individuals were infected with A. xylosoxidans. In contrast, none of the two study participants infected with A. insuavis had serum IgG against either of the two Achromobacter species. As a group, the CF_Achro_ sera had higher IgG titers against A. xylosoxidans compared to CF_Ctrl_ (P = 0.02, Fig. 1A), whereas the difference in titers against A. insuavis was not significant between the CF_Achro_ and CF_Ctrl_ groups (P = 0.07, Fig. 1B). Surprisingly, CF_PsA_ sera had significantly higher median titers than CF_Ctrl_ sera against both A. xylosoxidans (P = 0.002, Fig. 1A) and A. insuavis (P = 0.0007 Fig. 1B). Therefore, to test whether Achromobacter infection triggers antibodies reactive against P. aeruginosa, all sera were also screened for IgG titers against P. aeruginosa. A majority of the CF_PsA_ group (64%) had IgG titers against P. aeruginosa (Fig. 1C). However, CF_Achro_ patients did not have serum IgG against P. aeruginosa, except for three patients who were infected with both Achromobacter and Pseudomonas.
Antibody responses of donor serum against whole bacteria and Achromobacter proteins. Serum samples from pwCF and healthy controls were analyzed for IgG titers against Achromobacter xylosoxidans (A), A. insuavis (B), and Pseudomonas aeruginosa (C) by direct ELISA using plates coated with whole bacteria. Bars represent the mean OD420 value per group. Each dot is the average value of three technical replicates. Serum from individuals infected with both Achromobacter and P. aeruginosa (gray triangles) was included in the Achromobacter group when screening for those antibodies (A, B), and vice versa for P. aeruginosa (C). The dotted lines represent the average of the healthy control group +3 SD, which was considered the cutoff value for positive titers. Groups were compared using the Mann-Whitney test. (D, E) Western blot assay of serum IgG binding to A. xylosoxidans proteins. Secreted proteins (D) and trypsinated surface fractions (E) were separated on SDS-PAGE followed by Western blotting using CFAchro and healthy control serum.
Since only patients infected with A. xylosoxidans, but not A. insuavis, raised IgG responses against Achromobacter spp., we wanted to identify A. xylosoxidans protein antigens responsible for inducing adaptive immunity in these individuals. Since the IgG responses observed in Fig. 1A were directed against whole bacterial cells, we prepared secreted and surface fractions of A. xylosoxidans for the identification of specific protein antigens (Fig. S1). These protein fractions were separated by SDS-PAGE, followed by Western blotting using serum from a patient in the CF_Achro_ group with high titers against A. xylosoxidans. The results demonstrated multiple bands both in the secreted and surface fractions, indicating IgG-binding to bacterial antigens (Fig. 1D and E). In contrast, no IgG binding was detected with serum from a healthy control (Fig. 1D and E). This qualitative screening suggests that A. xylosoxidans infection results in circulating antibodies against various A. xylosoxidans secreted and surface proteins.
Identification of Achromobacter protein antigens
Having confirmed that A. xylosoxidans airway infection can give rise to circulating antibodies against Achromobacter, we next aimed to quantitatively identify these bacterial antigens. LC-MS/MS analysis of the secreted and surface fractions of Achromobacter identified a total of 357 bacterial proteins in the secreted fraction and 1,163 proteins in the surface fraction (File S1). Flagellin was the most abundant protein in both fractions, followed by T1S-DCP in the secreted fraction and chaperonin in the surface fraction. To quantitatively identify Achromobacter antigens triggering an antibody response, we took advantage of a newly established mass spectrometry-based systems antigenomics workflow (12), wherein secreted and surface fractions were pooled and passed over immobilized IgG from serum of different sources. The affinity purification resulted in antigen-antibody complexes, which were then eluted by glycine and analyzed using LC-MS/MS (Fig. 2A; File S2). Serum from an individual with high IgG titers against A. xylosoxidans (CF_Achro_ serum) was used in this experimental setup. To control for non-specific IgG binding, the monoclonal anti-IgE antibody omalizumab was included as a negative control. When comparing proteins enriched from IgG pulldowns using CF_Achro_ serum to those from the negative control omalizumab, eight proteins were found to be significantly associated with IgG from CF_Achro_ serum (Fig. 2B; File S2), suggesting that Achromobacter infection triggers an antibody response against a small subset of proteins. These antigens were predominantly present in both secreted and surface fractions and were involved in different cellular functions such as iron binding, enzymatic activity, and membrane transport (Table 1). To further determine that the antibody response against these antigens was a result of Achromobacter infection, we also performed the analysis on pooled IgGs from multiple individuals (IVIG) and serum from a healthy donor. IVIG and pooled human plasma were considered negative controls as Achromobacter is an opportunistic pathogen that rarely causes infection in healthy individuals. All eight A. xylosoxidans antigens identified with CF_Achro_ serum were absent in the IgG pulldowns using IVIG and healthy donor serum (Fig. S2), further supporting that the IgG response induced by Achromobacter infection is highly specific.
Identification of antigens in Achromobacter xylosoxidans. (A) Brief description of the systems antigenomics workflow used to identify antigenic proteins. (B) Enrichment analysis of bacterial proteins associated with serum from pwCF infected with Achromobacter compared to omalizumab, two-tailed t-test, FDR 0.05. Panel A created with BioRender.com.
Specificity of identified antigens to A. xylosoxidans
To rule out antigens which may also be targeted by P. aeruginosa antibodies, we also performed IgG affinity purification of Achromobacter proteins using IgG derived from an individual with P. aeruginosa infection who had high antibody titers against A. xylosoxidans (CF_PsA_ serum, Fig. 1A). This individual was not co-infected with A. xylosoxidans and had no known history of Achromobacter infection. When comparing intensities of A. xylosoxidans antigens purified with IgG derived from CF_Achro_ and CF_PsA_ serum respectively, four antigens were significantly associated with CF_Achro_ serum compared to CF_PsA_ serum (Fig. 3A). These proteins were DLD (A0A0D6GPL2), T1S-DCP (A0A7T2RJE8), DUF336 (A0A0D6GMC1), and amino acid ABC transporter substrate-binding protein (ABC-TSBP, A0A0D6FRA6). Additional repeats of the IgG affinity purification confirmed that all four antigens had significantly higher intensities in pulldowns using CF_Achro_ serum, and three of the antigens (DLD, DUF336, and ABC-TSBP) were not identified at all in pulldowns using CF_PsA_ serum (Fig. 3B). The four antigens were distributed across the intensity range of proteins identified from the pooled bacterial fractions (Fig. 3C), suggesting that the affinity purification was not biased towards highly abundant proteins.
Specificity of Achromobacter antigens compared to anti-P. aeruginosa IgG. (A) Enrichment analysis of bacterial proteins associated with CFAchro serum compared to CFPsA serum, two-tailed t-test, FDR 0.05. (B) LFQ intensities of the Achromobacter antigens DUF336, T1S-DCP, DLD, and ABC-TSBP in pulldowns using CFAchro and CFPsA serum are compared using the Mann-Whitney U-test. n = 6, ** = P < 0.01. (C) Abundance of significant Achromobacter antigens in the total bacterial protein fraction. LFQ intensities (log10) of all 935 identified Achromobacter proteins in the pooled bacterial protein fraction are plotted in descending order. The four CFAchro-associated bacterial antigens are marked with red triangles. (D) ELISA plates were coated with recombinantly expressed antigens, and IgG titers were tested in the same sera used in the IgG pulldown assay.
To further validate the IgG pulldown results, we selected three antigens, i.e., DLD, T1S-DCP, and DUF336, that were recombinantly expressed in E. coli. The recombinant proteins were used to coat ELISA plates, after which the patient and control sera used in the affinity purification protocol were added to quantify IgG binding. In agreement with the IgG pull-down results, we observed high IgG titers to all these three recombinant proteins in the CF_Achro_ serum, whereas the healthy donor serum and CF_PsA_ serum did not have IgG titers against any of the three antigens (Fig. 3D). Taken together, the ELISA results with recombinant antigens confirmed the identification of specific A. xylosoxidans antigens using the MS-based systems antigenomics workflow.
Evaluation of A. xylosoxidans antigens in a wider CF cohort
To further investigate the specificity of A. xylosoxidans antigens, we performed a BLAST search and amino acid sequence alignment of the three antigens to identify similar proteins in A. insuavis and P. aeruginosa. Using a cutoff of 50% alignment score to define protein homology, DUF336 showed a high similarity to heme-binding proteins in A. insuavis (sequence homology 97%) and in P. aeruginosa (56.8%). DLD was found in both species and was considered homologous to A. xylosoxidans DLD in P. aeruginosa (68%) but not in A. insuavis (31.9%) (File S3). No homologous proteins to T1S-DCP were identified in either A. insuavis or P. aeruginosa.
We further aimed to confirm that DLD, T1S-DCP, and DUF336 are commonly present in A. xylosoxidans isolates and not specific to the type strain used in the LC-MS/MS experiment. PCR amplification of the genes corresponding to the three antigens was therefore performed in a collection of 14 A. xylosoxidans isolates. Among these isolates, five were obtained from the individuals in the CF_Achro_ serum group. All genes could be amplified in all isolates (Fig. S3), suggesting that the three genes are common across A. xylosoxidans and not unique to the type strain used in this study.
Having confirmed that all three antigens seem to be universally present in A. xylosoxidans isolates, we investigated the presence of specific IgG in a larger cohort of sera from pwCF. To this end, the recombinantly expressed antigens were used in an ELISA to quantify IgG titers in all 53 sera included in the study. As in the initial ELISA using whole bacteria, sera from two patients infected with A. xylosoxidans stood out as having high titers against all three antigens. Using the mean OD value +three standard deviations of the healthy control group as a cut-off for positivity, 71% of the CF_Achro_ sera had positive IgG titers against DLD and T1S-DCP, whereas 43% were positive against DUF336 (Fig. 4; Table 2). T1S-DCP was the least specific antigen, displaying a large proportion of positive serum titers in all groups except healthy controls (Fig. 4C; Table 2). Taken together, antibody responses to DUF336 and possibly also DLD appear to be specific to Achromobacter infection.
Serum IgG binding to recombinantly expressed A. xylosoxidans antigens. ELISA plates were coated with recombinantly expressed dihydrolipoyl dehydrogenase (A), domain of unknown function (DUF336) (B), and type I secretion C-terminal target domain-containing protein (C), and IgG titers in sera from pwCF and healthy controls were measured by direct ELISA. Each dot represents the average value of three technical replicates, and bars represent the mean value. The dotted lines represent the average of the healthy control group + 3 SD, which was considered the cutoff for positivity.
DISCUSSION
In the present study, we report that 57% (4/7) pwCF infected with Achromobacter spp. develop circulating IgG against whole bacteria, and we present an in-depth characterization of Achromobacter protein antigens that trigger an antibody host response. While some studies have attempted to discover lymphocyte epitopes using predictive tool approaches in silico (18, 19), our study is to our knowledge the first to investigate A. xylosoxidans antigens using patient serum IgG for affinity purification of bacterial proteins. Using his method, we identified eight antigens associated with CF_Achro_ serum-IgG, four of which (T1S-DCP, DLD, DUF336, and ABC-TSBP) did not show cross-reactivity with anti-Pseudomonas IgG. The role or function of these four antigens has not been previously characterized in A. xylosoxidans. However, T1S-DCP is associated with the type I secretion system, which mediates translocation of proteins across the cell membrane (20). The DUF336 was predicted by sequence homology to be part of a heme-binding protein (File S3), possibly as a hemophore (21). DLD exhibits dehydrogenase activity and has been implicated in complement evasion in P. aeruginosa (22, 23). Finally, ABC-TSBP is associated with ABC transporters, which have been implicated as therapeutic and vaccine targets as they are essential for the translocation of a wide variety of substrates across cell membranes (24).
Three of the antigens identified in the pull-down experiments (T1S-DCP, DLD, and DUF336) were expressed recombinantly, and antibody titers against recombinant proteins were confirmed in several CF_Achro_ sera. The corresponding genes were also confirmed to be present in all Achromobacter spp. isolates tested in this study. Taken together, these antigens could be potential candidates for serologic testing of Achromobacter serum antibodies. With the introduction of CFTR modulator therapies, the use of serological approaches in CF infection diagnostics may come to play a greater role (25). After the start of CFTR modulator treatment, many pwCF experience a marked reduction in sputum production and dramatically improved lung function and quality of life. However, pathogenic microorganisms that colonize the airways may remain or rebound despite modulator therapy (26, 27), and reduced sputum production can limit the possibility of monitoring chronic airway infections via surveillance cultures (28). Serological assays have been implicated for clinical routine use to monitor P. aeruginosa infection status (29), as serum antibodies to P. aeruginosa have the potential for early detection of chronic P. aeruginosa infection (30, 31).
However, the utility of serum antibodies against bacteria in pwCF may be limited by cross-reactivity between CF pathogens (32). A strength of the present study is the comparison of antigens retrieved from CF_Achro_ and CF_PsA_ serum-IgG, respectively, which allows the exclusion of cross-reactive antigens. Our data suggest that especially the A. xylosoxidans antigens DLD and DUF336 could be interesting candidates for the detection of specific Achromobacter serum IgG in serologic assays. However, the results need to be further validated against larger patient cohorts.
Although IgA is the dominating immunoglobulin in airway secretions, IgG is present in the lower respiratory tract and plays an important role in airway immunity. IgG in the airways comes from local secretion from the airway mucosa and intraluminal lymphocytes, or via transudation from plasma (33). In CF, titers of circulating IgG against specific pathogens have been associated with low lung function and a worse prognosis. For example, the presence of precipitating serum antibodies against Achromobacter has previously been demonstrated by Rønne Hansen et al. 11, who also described great variability between individuals and a correlation between higher precipitin levels and a more rapid lung function decline. Similarly, rapidly increasing Pseudomonas precipitin levels are associated with poor prognosis (34), suggesting that antibody titers reflect the severity of infection.
Notably, neither of the two patients infected with A. insuavis in our study had circulating IgG against Achromobacter, whereas four out of five patients with A. xylosoxidans infection had positive serum titers. A. insuavis exoproducts are less potent in triggering inflammatory responses from airway epithelial cells in vitro compared to A. xylosoxidans (14), and it could be speculated that these findings suggest a difference in pathogenicity and immune evasion patterns between the two species. However, larger studies on clinical outcomes would be necessary to determine such differences.
The most important limitation of this study is the low availability of sera from pwCF colonized with Achromobacter. Due to the limited number of Achromobacter cases, we cannot investigate any associations between antibody titers and duration of infection, lung function, or effects of exacerbations and antibiotic treatment. The data are further confounded by the presence of both A. xylosoxidans and A. insuavis infections in the Achromobacter cohort. As the infection is uncommon in an already relatively small patient group, multi-center studies would be required to increase the sample size. Moreover, only one CF_Achro_ serum sample was used for the identification of A. xylosoxidans antigens. Although IgG against the identified antigens could be verified in serum from other pwCF, additional relevant antigens may have been captured in other serum samples due to individual variations. There is also a possibility of false-negative results arising from the limited statistical power of the analysis. Additionally, we were only able to identify protein antigens expressed by the specific bacterial isolate under the growth conditions used in this study. Other relevant antigens may be expressed during in vivo growth in the CF lung. Moreover, the antigens investigated in this study were obtained from the secretome and surface proteome fractions. There may be additional IgG targets, such as cytosolic proteins or polysaccharide antigens, which were not included in this experimental setup. However, the identified protein antigens provided a similar pattern of IgG titers in patient serum when compared to serum titers against whole bacteria, suggesting that they are representative of a host antibody response against Achromobacter infection.
In conclusion, most pwCF with A. xylosoxidans airway infection raise an antibody response against A. xylosoxidans. We identified four specific A. xylosoxidans antigens, out of which three were expressed recombinantly and validated by ELISA. In particular, DUF336 and DLD were highly specific to the sera from pwCF with A. xylosoxidans infection and could be candidates for the development of a serological assay.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Blanchard AC, Waters VJ. 2019. Microbiology of cystic fibrosis airway disease. Semin Respir Crit Care Med 40:727–736. doi:10.1055/s-0039-169846431887768 PMC 7117079 · doi ↗ · pubmed ↗
- 2Isler B. 2020. Achromobacter infections and treatment options. Antimicrob Agents Chemother 64:11. doi:10.1128/AAC.01025-20PMC 757712232816734 · doi ↗ · pubmed ↗
- 3Esposito S, Pisi G, Fainardi V, Principi N. 2021. What is the role of Achromobacter species in patients with cystic fibrosis? Front Biosci (Landmark Ed) 26:1613–1620. doi:10.52586/505434994175 · doi ↗ · pubmed ↗
- 4Kerem E, Orenti A, Zolin A, Annicchiarico L, Drevinek P, ECFSPR with the list of contributing authors. 2023. Clinical outcomes associated with Achromobacter species infection in people with cystic fibrosis. J Cyst Fibros 22:334–343. doi:10.1016/j.jcf.2022.11.00136418214 · doi ↗ · pubmed ↗
- 5Spilker T, Vandamme P, Lipuma JJ. 2013. Identification and distribution of Achromobacter species in cystic fibrosis. J Cyst Fibros 12:298–301. doi:10.1016/j.jcf.2012.10.00223141756 · doi ↗ · pubmed ↗
- 6Gade SS, Nørskov-Lauritsen N, Ridderberg W. 2017. Prevalence and species distribution of Achromobacter sp. cultured from cystic fibrosis patients attending the Aarhus centre in Denmark. J Med Microbiol 66:686–689. doi:10.1099/jmm.0.00049928481740 · doi ↗ · pubmed ↗
- 7Firmida MC, Pereira RHV, Silva EASR, Marques EA, Lopes AJ. 2016. Clinical impact of Achromobacter xylosoxidans colonization/infection in patients with cystic fibrosis. Braz J Med Biol Res 49:e 5097. doi:10.1590/1414-431X 2015509726909788 PMC 4792508 · doi ↗ · pubmed ↗
- 8De Baets F, Schelstraete P, Van Daele S, Haerynck F, Vaneechoutte M. 2007. Achromobacter xylosoxidans in cystic fibrosis: prevalence and clinical relevance. J Cyst Fibros 6:75–78. doi:10.1016/j.jcf.2006.05.01116793350 · doi ↗ · pubmed ↗