Deletion of RAP1 affects iron homeostasis, azole resistance, and virulence in Candida albicans
Min-Chi Yang, Wei-Luen Huang, Hsuan-Yu Chen, Shin-Huey Lin, Yu-Shan Chang, Kuo-Yun Tseng, Hsiu-Jung Lo, I-Ching Wang, Chi-Jan Lin, Chung-Yu Lan

TL;DR
Deleting the RAP1 gene in Candida albicans affects iron regulation, antifungal resistance, and the fungus's ability to cause disease.
Contribution
This study reveals new roles of RAP1 in iron homeostasis, azole resistance, and virulence in Candida albicans.
Findings
RAP1 deletion reduces Candida albicans virulence in a mouse model.
RAP1 is involved in iron acquisition and regulation of iron-related genes.
RAP1 contributes to fluconazole resistance under low-iron conditions.
Abstract
Rap1 is a DNA-binding protein conserved from yeast to mammals for its role in telomeric maintenance. Here, to explore additional functions of Candida albicans Rap1, we performed RNA sequencing analysis. Experimental validations further showed that Rap1 plays a role in iron regulation, especially under low-iron conditions. Moreover, Rap1 was involved in iron acquisition and modulation of iron-related genes. Rap1 was found to be associated with fluconazole resistance in a low-iron condition. Finally, we demonstrated that the deletion of RAP1 leads to reduced C. albicans virulence in a mouse model of infection. Together, this study reveals new functions of C. albicans Rap1, particularly in iron homeostasis, azole resistance, and virulence. Candida albicans is an important pathogenic fungus that can cause superficial to life-threatening infections. Iron is essential for almost all…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
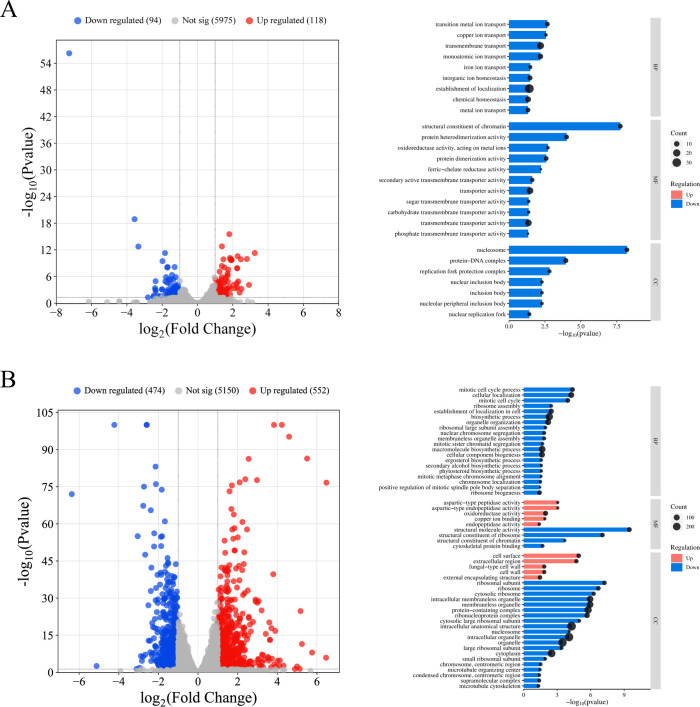 Fig 1
Fig 1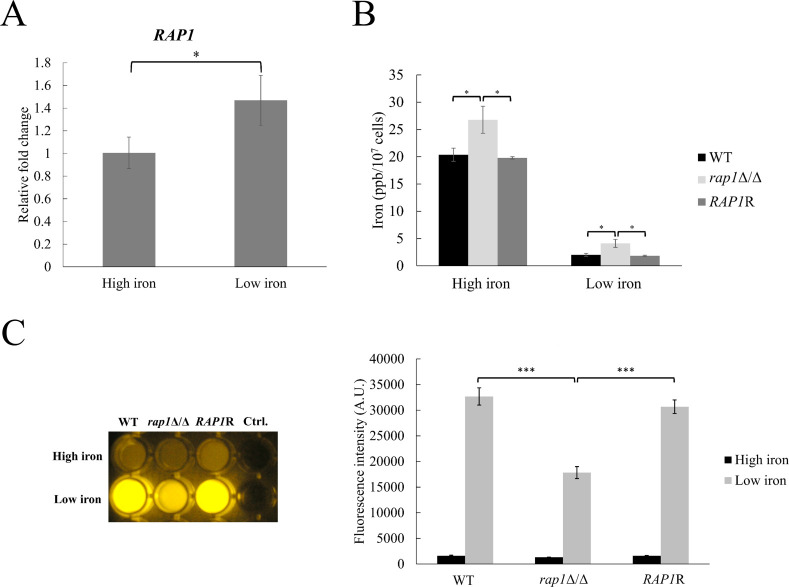 Fig 2
Fig 2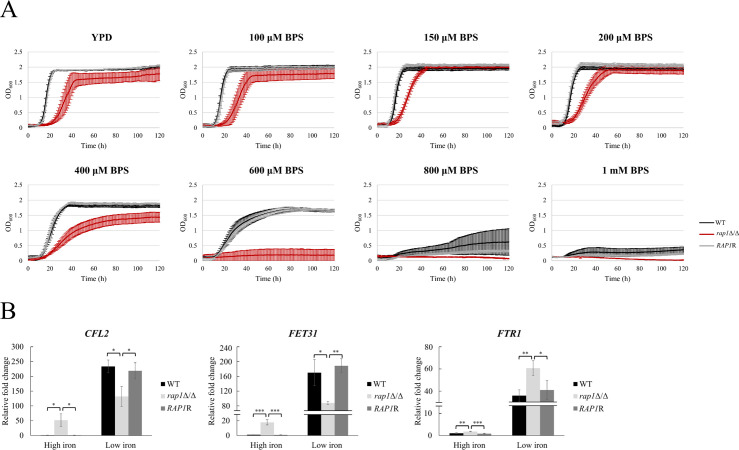 Fig 3
Fig 3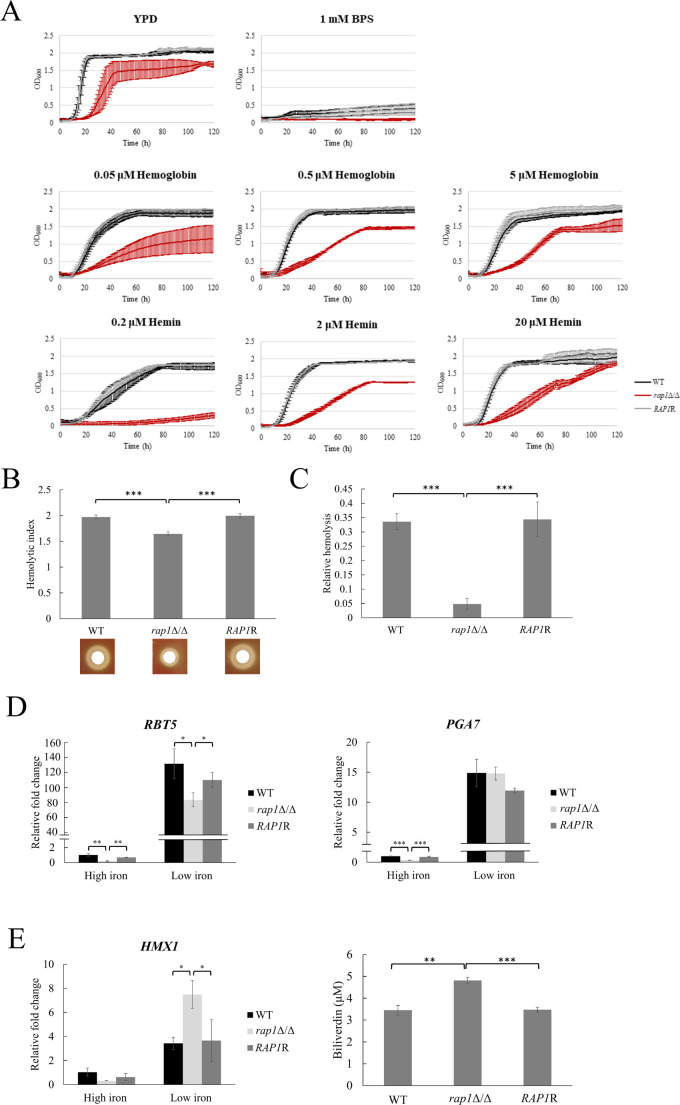 Fig 4
Fig 4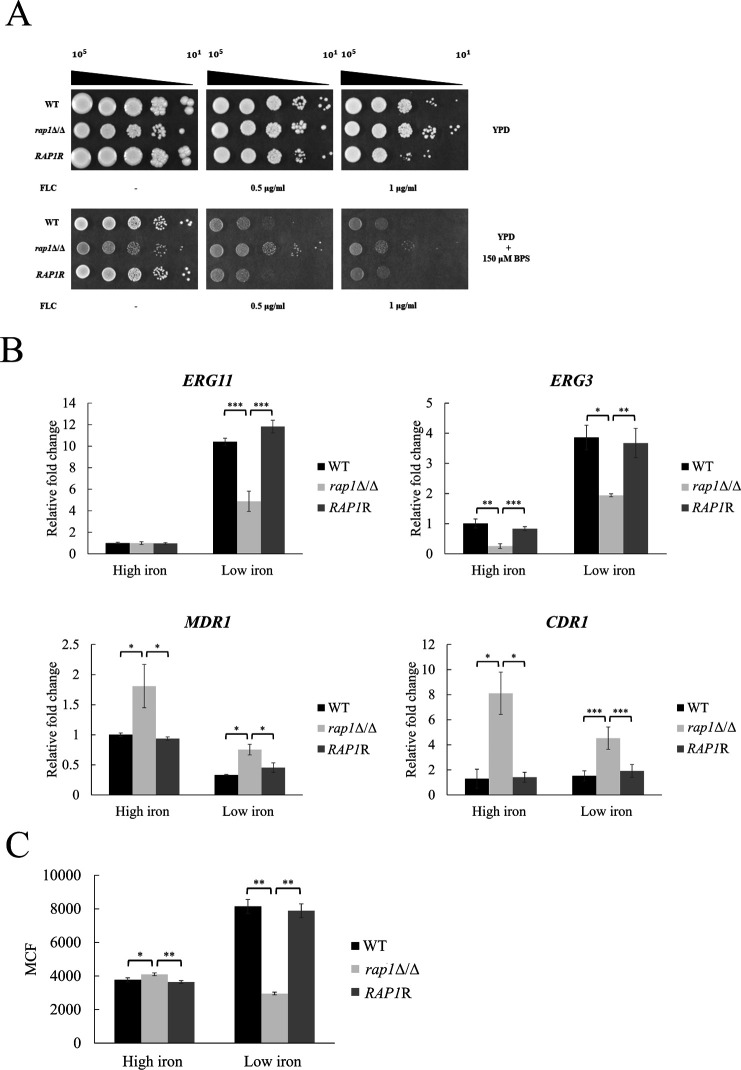 Fig 5
Fig 5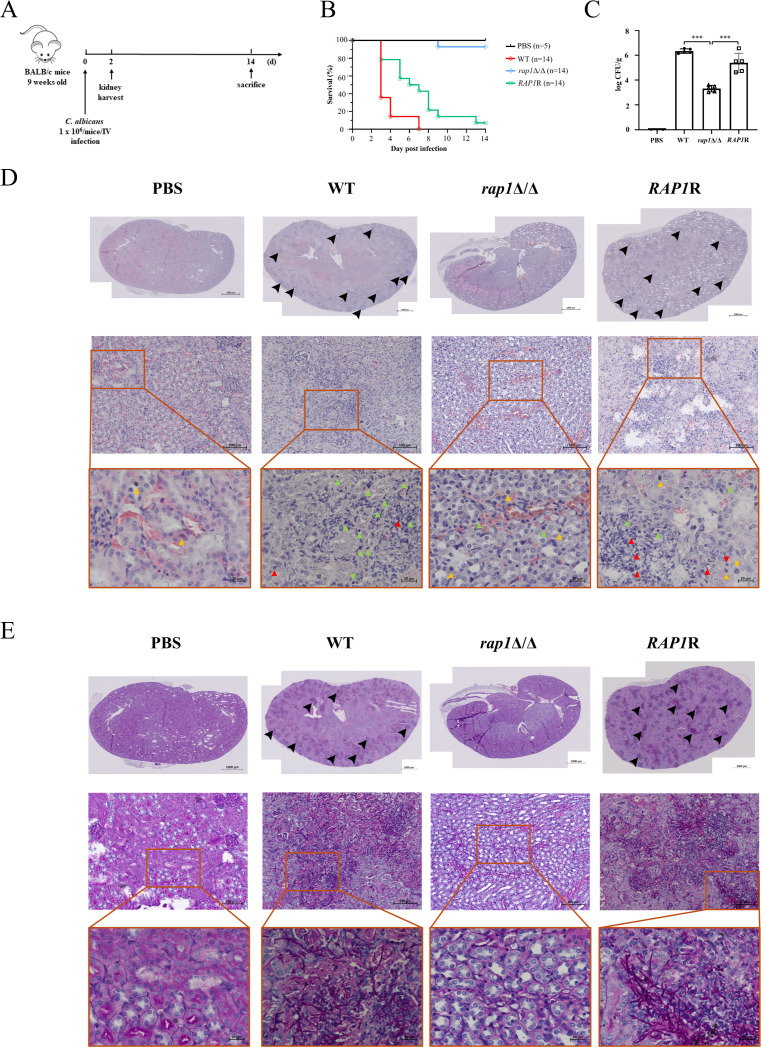 Fig 6
Fig 6| Condition | Strains | 24 h MIC50 (μg/mL) | 48 h MIC50 (μg/mL) |
|---|---|---|---|
| YPD | WT | 0.5 | 0.5 |
| 1 | 1 | ||
| 0.5 | 0.5 | ||
| YPD + 150 µM BPS | WT | 0.25 | 0.25 |
| 1 | 0.5 | ||
| 0.25 | 0.25 |
- —National Science and Technology Councilhttp://dx.doi.org/10.13039/501100020950
- —National Science and Technology Councilhttp://dx.doi.org/10.13039/501100020950
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsAntifungal resistance and susceptibility · Fungal Infections and Studies · Parasitic Infections and Diagnostics
INTRODUCTION
Candida albicans is a commensal fungus that normally inhabits the skin and mucosal surfaces of humans (1). However, C. albicans is also an opportunistic pathogen and can cause superficial to life-threatening invasive infections, especially in immunocompromised patients (2). Moreover, antifungal resistance of C. albicans holds another emergent threat to public health (3). Therefore, C. albicans has recently been included in the World Health Organization’s list of fungal priority pathogens, highlighting the urgent need for research and drug development for this important pathogen (4).
Iron is an essential micronutrient for C. albicans and its host, and it plays a crucial role in host-pathogen interactions (5). The human host employs various strategies to limit iron availability to C. albicans, known as nutritional immunity (6, 7). For example, host proteins such as transferrin and ferritin sequester iron, making it less available to C. albicans. To adapt to the iron-restricted host environment, C. albicans has developed three elaborate mechanisms for iron acquisition. First, C. albicans utilizes a reductive iron uptake system to acquire free iron, as well as sequestering iron from the host’s iron-binding proteins (8, 9). Ferric iron (Fe^3+^) is initially reduced to ferrous iron (Fe^2+^) by ferric reductases (5). The generated Fe^2+^ is oxidized again by multicopper ferro-oxidases, and Fe^3+^ is transported into the cell by the high-affinity iron transporter Ftr1 (5). Second, C. albicans also possesses a system that acquires iron from the host’s heme-binding proteins such as hemoglobin and serum albumin. This system is mediated by distinct hemophore proteins including Csa2, Rbt5, and Pga7 (10–12). Through a cascade of these proteins, heme is endocytosed into C. albicans cell and degraded by the vacuolar heme oxygenase Hmx1, releasing iron into the cytosol (10–12). Third, a siderophore-mediated iron acquisition has been identified in C. albicans. Although C. albicans does not produce its own siderophores, this pathogen uptakes xenosiderophores via a siderophore transporter, Sit1 (13).
Importantly, iron acquisition is also closely associated with pathogenicity and virulence of C. albicans. For example, the ferric reductase CFL1-deletion mutant shows attenuated virulence and reduced fungal burden in a mouse systemic infection model (14), and the mutant lacking FTR1 is unable to establish systemic infection in mice (15). Moreover, Sit1 is required for epithelial invasion, while the HMX1 null mutant exhibits decreased virulence and alters chemokine and cytokine expression in infected mice (13, 16). In addition, the reductive iron uptake system also affects biofilm formation, and iron deprivation induces hyphal development in C. albicans (17, 18). Finally, there is a link between iron and antifungal susceptibility (19). Iron depletion leads to increased susceptibility to antifungal agents in C. albicans due to alterations in membrane fluidity and permeability (20). The iron chelator deferasirox can reduce C. albicans invasion of oral epithelial cells and infection in murine oropharyngeal candidiasis (21).
Rap1 is a DNA-binding protein conserved in mammals, protozoa, and yeast (22). Saccharomyces cerevisiae Rap1 (ScRap1) regulates telomere homeostasis and length (23). Moreover, ScRap1 also possesses non-telomeric functions, including gene transcription and regulation of the ferric reductase FRE1, DNA repair, and ribosomal protein genes (24–26). In C. albicans, Rap1 is also crucial in controlling telomere length and structure (27, 28). Intriguingly, our recent study revealed that C. albicans Rap1 is also involved in cell wall integrity, biofilm formation, and virulence in a Galleria mellonella infection model (29). Nevertheless, our knowledge of other functions of C. albicans Rap1 remains limited.
In this study, we aimed to uncover previously unknown functions of Rap1 in C. albicans. Based on prior finding that ScRap1 is involved in regulating FRE1 gene expression (25), we hypothesized that Rap1 might play roles related to iron availability. Our results showed that RAP1 gene deletion affected cellular utilization of various iron sources and expression of iron acquisition-related genes. Additionally, we demonstrated that Rap1 contributes to fluconazole susceptibility, particularly under low-iron conditions. Notably, RAP1 deletion also leads to attenuation of C. albicans virulence in a mouse model of systemic infection. Together, our findings reveal unexplored functions of Rap1 in C. albicans.
RESULTS
Iron-responsive transcriptome profiling of the RAP1 deletion and wild-type strains
RNA-sequencing (RNA-seq) analysis was performed to identify differentially expressed genes (DEGs) in C. albicans. The RAP1-deleted (rap1Δ/Δ) and wild-type strains were pre-starved for iron by incubation with YPD medium containing 400 µM of the iron chelator bathophenanthrolinedisulfonate (BPS). Following this pre-treatment, cells were grown in YPD (high-iron condition) or YPD supplemented with 200 µM BPS (low-iron condition). These conditions were selected based on the differential activation of the iron transporters FTR2 and FTR1 under high- and low-iron environments, respectively, as previously described (15) and illustrated in Fig. S1.
DEGs were assessed using an adjusted P-value < 0.05 and an expression change of at least twofold (log2 fold change = 1). Under the low iron condition (YPD with 200 µM BPS), 118 genes were significantly upregulated and 94 genes were downregulated in the rap1Δ/Δ mutant. These changes are visualized in a volcano plot (Fig. 1A) and detailed in Table S1. Moreover, Gene Ontology (GO) analysis was performed. Although no GO terms were enriched among the upregulated genes, the downregulated genes were associated with multiple GO categories, including “iron ion transport,” “transition metal ion transport,” and “ferric-chelate reductase activity” (Fig. 1A; Table S2). For example, several ferric reductases and ferro-oxidase genes exhibited reduced expression in the rap1Δ/Δ mutant compared to the wild-type strain (Table S1). Interestingly, given the essential role of copper as a cofactor for ferro-oxidases (30, 31), a GO term associated with copper transport was also identified among the downregulated genes of the rap1Δ/Δ mutant (Tables S1 and S2).
Transcriptome profiling of the rap1Δ/Δ mutant compared to the wild-type (WT) strain. (A) Volcano plot of RNA-seq data and bar graph of gene ontology (GO) enrichment analysis comparing gene expression in the rap1Δ/Δ mutant relative to the WT strain in YPD supplemented with 200 µM BPS (low iron) medium. GO terms significantly overrepresented among differentially expressed genes (DEGs) with a fold change ≥ 2 and adjusted P-value < 0.05. No GO terms were enriched among the upregulated genes. (B) Volcano plot and GO enrichment analysis of RNA-seq data for the rap1Δ/Δ mutant relative to WT in YPD (high iron) medium. Significantly upregulated and downregulated genes are shown in red and blue, respectively. “Not sig” indicates genes with no significant changes. BP, biological processes; CC, cellular components; MF, molecular functions.
Additionally, under the high iron condition, 552 genes were significantly upregulated, while 474 genes were downregulated in the rap1Δ/Δ mutant (Fig. 1B; Table S1). GO analysis classified these DEGs into multiple categories (Fig. 1B; Table S2), including “ergosterol biosynthetic process.” Overall, RNA-seq analysis suggests that C. albicans Rap1 plays pleiotropic roles in cellular homeostasis, including iron-responsive activities.
Rap1 seems to mainly function in cells that are grown under low-iron conditions
To correlate Rap1 and iron-responsive functions, the effects of RAP1 deletion on several iron-related traits were determined. In Fig. 2A, increased RAP1 expression was detected in the low-iron condition using real-time quantitative PCR (qPCR). Moreover, although the details about intracellular iron trafficking in C. albicans are still not clear, cellular iron homeostasis is associated with the availability of environmental iron (14, 19). To further examine the relationship between Rap1 and iron response, the intracellular iron levels were measured using inductively coupled plasma mass spectrometry (ICP-MS). The results indicated that cells had much more intracellular iron contents when they grew in the high-iron YPD medium than in the low-iron YPD with BPS (Fig. 2B). In addition, the rap1Δ/Δ mutant had a higher intracellular iron level than that in the wild-type and RAP1-reintegrated control strains under either low- or high-iron condition (Fig. 2B). Thus, these data suggest that Rap1 likely functions in response to iron availability and acts preferentially toward the iron-limited condition. To test this hypothesis, flavin production was also assessed, as it is known that flavin production is induced by iron limitation (32). For the convenience of observation and spectrometric measurement, transparent yeast nitrogen base (YNB) non-iron medium (NIM) and synthetic complete (SC) medium were here used as the low- and high-iron condition, respectively (32). In Fig. 2C, the low secreted flavin levels of all tested strains were observed in the high-iron SC medium. However, secreted flavin levels were largely induced in the low-iron condition (NIM). Importantly, the rap1Δ/Δ mutant showed approximately 50% less flavin production compared to the control strains. Together, our findings indicate that deletion of RAP1 affects iron-related traits, particularly in low-iron conditions.
*Rap1 functions in low iron conditions. (A) RAP1 transcript level was determined by real-time qPCR. The ACT1 transcripts were used as endogenous control. Data are collected from three independent experiments and presented as the mean ± standard deviation (SD). (B) Intracellular iron content was measured using ICP-MS. Data are collected from three independent experiments and presented as the mean ± SD. (C) Measurement of flavin production. Cells were cultured in high-iron (SC) or low-iron (NIM) media for 48 h. Left panel: culture supernatants were collected and photographed under UV excitation. The results from one of three replicates are displayed. Right panel: supernatants collected from cell culture were used to measure fluorescence upon excitation 450 nm and emission 530 nm. The values were normalized to cell numbers and presented as relative fluorescence intensity. WT, wild-type; rap1Δ/Δ, RAP1-deleted; RAP1R, RAP1-reintegrated strain. Data are collected from three independent experiments and presented as the mean ± SD. A.U., arbitrary units. *P < 0.05; **P < 0.001.
Rap1 is involved in iron utilization and iron acquisition-related gene expression
C. albicans has evolved to utilize diverse iron sources within the host environment via different iron acquisition systems (5). To further investigate iron-related functions of Rap1, iron utilization and expression of iron-acquisition genes were determined. For free iron utilization, iron-prestarved cells were grown in YPD broth with and without BPS (20, 32). As demonstrated in Fig. 3A, cell growth of all tested strains was inhibited under iron-limiting conditions, with increasing concentrations of BPS. However, the rap1Δ/Δ mutant was much more sensitive to iron limitation, particularly at 400 and 600 µM BPS, compared to control strains (Fig. 3A). These data suggest that the rap1Δ/Δ mutant is impaired in free iron acquisition, resulting in a more pronounced growth defect under low-iron conditions. To test this possibility, the expression of three representative genes of the reductive iron uptake system was assessed: the ferric reductase (CFL2), multicopper oxidase (FET31), and iron transporter (FTR1). In the low-iron condition, all three genes were largely induced (Fig. 3B). However, in the rap1Δ/Δ mutant, CFL2 and FET31 were downregulated, while FTR1 expression was enhanced compared to control strains under the same condition (Fig. 3B).
*Deletion of RAP1 affects utilization of different iron sources and expression of iron acquisition systems. (A) Iron pre-starved cells were inoculated into YPD broth supplemented with different concentrations of BPS. Data are collected from three independent experiments and presented as the mean ± SD. (B) Differential expression of genes in the reductive iron uptake system. Cells were harvested for RNA extraction, and gene expression was assessed by real-time qPCR. The ACT1 transcripts were used as endogenous control. Data are collected from at least three independent experiments and presented as the mean ± SD. *P < 0.05; **P < 0.01; **P < 0.001.
Rap1 is also associated with the acquisition and utilization of hemin and hemoglobin
Other than extracellular iron sources, C. albicans can also take up heme-bound iron of hemoglobin and hemin-containing iron from human erythrocytes. In this context, cell growth with hemoglobin and hemin was examined.
In Fig. 4A, all tested strains exhibited severe growth defects in YPD broth containing 1 mM BPS. These growth defects of the wild-type and RAP1-reintegrated control strains were easily recovered by adding different concentrations of hemoglobin and hemin. Although hemoglobin and hemin could also rescue the growth of the rap1Δ/Δ mutant; however, the mutant exhibited a growth delay in the presence of these iron sources compared to its growth in YPD broth alone (Fig. 4A).
*Rap1 is involved in heme uptake and utilization. (A) Iron pre-starved cells were inoculated into YPD broth supplemented with 1 mM BPS to prevent growth of C. albicans due to iron limitation. The indicated concentration of hemoglobin and hemin was added, and the cell growth curve was plotted. Data are collected from three independent experiments and presented as the mean SD. (B) Blood agar assay. Cells were spotted onto blood agar and incubated at 37°C, 5% CO2 for 24 h. Zones of beta hemolysis were assessed (lower panel). The hemolytic index was calculated from five independent experiments and presented as the mean ± SD (upper panel). (C) Hemolysis liquid assay. Yeast cells were cultured with sheep red blood cells at 37°C, 5% CO2 for 24 h, and the hemolytic activity was measured by absorbance at 414 nm. Results were collected from five independent experiments and presented as the mean ± SD. (D) Expression of the heme transporter genes. Cells were harvested for RNA extraction, and gene expression was assessed by real-time qPCR. The ACT1 transcripts were used as endogenous control. Data are collected from at least three independent experiments and presented as the mean ± SD. (E) Expression and activity of the heme oxygenase Hmx1. Left panel: expression of the heme oxygenase gene was measured using real-time qPCR. The ACT1 transcripts were used as endogenous control. Data are collected from at least three independent experiments and presented as the mean ± SD. Right panel: cells were cultured in the medium containing hemoglobin for 12 h and collected. The heme oxygenase activity was determined by measuring absorption of biliverdin at 666 nm. Data are collected from three independent experiments and presented as the mean ± SD. *P < 0.05; **P < 0.01; **P < 0.001.
To access heme iron from the host, C. albicans secretes hemolytic factors to lyse host red blood cells (RBCs) (33). Accordingly, the hemolytic activity of the rap1Δ/Δ mutant was further examined using blood agar and liquid hemolysis assay. In the agar assay, cells were spotted on agar plates containing 7% sheep blood, and hemolytic zones were recorded (34). As demonstrated in Fig. 4B (lower panel), all test strains formed β-hemolysis (translucent halo) and α-hemolysis (dark green halo) that represent complete and incomplete RBC degradation, respectively. Interestingly, RAP1 deletion resulted in reduced hemolytic zones compared to the control strains. The hemolytic index was further measured as previously described (34). Evidently, the rap1Δ/Δ mutants had a lower hemolytic index than the control strains (Fig. 4B, upper panel). Furthermore, the liquid hemolysis assay was performed by co-culturing C. albicans and sheep RBCs, and concentrations of extracellular hemoglobin levels were measured (33). The results showed that the rap1Δ/Δ mutant possesses a lower hemolytic activity (Fig. 4C).
Once hemoglobin is released from RBCs, C. albicans uptakes heme via heme transporters on the cell envelope. Notably, among these heme transporters, RBT5 is one of the most highly induced genes in G. mellonella (35), and Rbt5-specific antibodies are notable in candidemia patients (36). Moreover, the deletion of PGA7 attenuates virulence in a murine model of disseminated candidiasis (11). Thus, RBT5 and PGA7 expression was measured, and the results found that these genes were highly induced in the low-iron condition (Fig. 4D). Moreover, in the low-iron condition, RBT5 was downregulated in the rap1Δ/Δ mutant compared to the control strains (Fig. 4D).
After transferring across the cell envelope, heme is degraded by the heme oxygenase Hmx1 to release iron ions (37, 38). As expected, HMX1 expression was induced in the low-iron condition (Fig. 4E). Moreover, the rap1Δ/Δ mutant exhibited a higher HMX1 gene expression than the control strains in the low-iron condition (Fig. 4E). Finally, the activity of heme oxygenase was also assessed by growing cells in medium containing hemoglobin. As shown in Fig. 4E, the Hmx1 activity in the rap1Δ/Δ mutant was elevated compared to the controls, by measuring concentrations of biliverdin, a waste product of heme degradation (16).
In sum, our results showed that RAP1 deletion leads to alterations in iron utilization and iron acquisition gene expression, suggesting C. albicans Rap1 plays a role in iron homeostasis.
RAP1 deletion also affects fluconazole susceptibility in response to iron conditions
Azoles are commonly used antifungal drugs by targeting the sterol 14α-demethylase Erg11 to inhibit ergosterol biosynthesis, thereby blocking fungal cell membrane formation (3, 39). Disrupting ergosterol biosynthesis can also cause accumulation of toxic sterols catalyzed by the sterol C5-desaturase Erg3 (3, 39). Furthermore, iron availability affects fluconazole susceptibility in C. albicans (20). Interestingly, our RNA-seq data revealed differential expression of ergosterol biosynthesis genes in the rap1Δ/Δ mutant compared to the wild-type strain grown in YPD (Table S1). This observation raises the question of whether a relationship exists between Rap1 and fluconazole susceptibility with respect to iron availability.
To test this possibility, fluconazole susceptibility was determined using agar and broth microdilution methods. In the spot assay, cell growth of all tested strains showed no significant difference in YPD agar plates. However, the rap1Δ/Δ mutant displayed a slight resistance to fluconazole (1 µg/mL) than the control strains (Fig. 5A). Apparently, the rap1Δ/Δ mutant was even much more resistant to fluconazole than the control strains in YPD agar with BPS added (Fig. 5A). Similarly, the broth microdilution assay demonstrated that all tested strains had higher minimum inhibitory concentration 50 (MIC50) in YPD compared to that in YPD containing BPS (Table 1). Moreover, the rap1Δ/Δ mutant had higher MIC50, suggesting more resistance of the mutant to fluconazole than the control strains (Table 1).
*The RAP1 deletion mutant is more resistant to fluconazole in response to iron condition. (A) Fluconazole spot assay. Iron pre-starved cells were spotted on YPD agar plates in the presence or absence of BPS and fluconazole (FLC). (B) Expression of ergosterol biosynthesis and drug pump genes. Exponential phase cells were harvested for RNA extraction, and gene expression was assessed by real-time qPCR. The ACT1 transcripts were used as endogenous control. Data are collected from at least three independent experiments and presented as the mean ± SD. (C) The efflux activity of drug pump was measured using the dye Nile red. MCF, mean channel fluorescence. Data are collected from three independent experiments and presented as the mean ± SD. *P < 0.05; **P < 0.01; **P < 0.001.
Several mechanisms are known to contribute to fluconazole resistance. For example, overexpression or point mutations of the ERG11 gene, and mutations in Erg3 by preventing toxic sterol accumulation can all lead to drug resistance (40–43). To further investigate if fluconazole resistance in the rap1Δ/Δ mutant is linked to ergosterol biosynthesis in response to iron availability, the expression of ERG11 and ERG3 was measured from cells grown in the low- and high-iron condition. Expression of ERG11 and ERG3 genes was largely induced in the low-iron condition (Fig. 5B). Moreover, the rap1Δ/Δ mutant showed a lower expression level of ERG11 and ERG3 than the control strains, particularly under the low-iron condition (Fig. 5B).
Another mechanism of azole resistance involves overexpression of efflux pump genes. Therefore, real-time qPCR analysis was performed, and the results indicated that the expression of CDR1 and MDR1 was elevated in the rap1Δ/Δ mutant under both low- and high-iron conditions (Fig. 5B). Finally, since Nile red is a known fluorescent substrate for Cdr1 and Mdr1 transporters, efflux pump activity was further assessed by measuring Nile red accumulation in the cells (44). In Fig. 5C, Nile red accumulation was decreased particularly in the rap1Δ/Δ mutant compared to the control strains under low-iron condition, suggesting a stronger efflux activity in the rap1Δ/Δ. Together, the deletion of RAP1 led to fluconazole resistance under low-iron condition, and this phenotype is associated with alterations in ergosterol biosynthesis and drug efflux pumps.
Loss of RAP1 impedes C. albicans virulence in a mouse model of systemic infection
The human body maintains extremely low levels of free iron to defend against pathogens, and iron acquisition is closely associated with the virulence of C. albicans (45). Given that Rap1 is relevant to iron homeostasis, we, therefore, used a mouse model of systemic candidiasis and histological analysis to determine the role of Rap1 on C. albicans virulence and pathology (Fig. 6A). The wild-type and RAP1-reintegrated strains had high virulence, by exhibiting rapid death of the mice beginning on the second day post-infection, and almost all mice in these two groups died during the experiment (Fig. 6B). Conversely, more than 90% of mice injected with the rap1Δ/Δ mutant remained alive within 14 days post-infection (Fig. 6B). The death rate of mice infected with the rap1Δ/Δ mutant was significantly lower than that of the wild-type (P-value < 0.0001) and RAP1-reintegrated strain (P-value = 0.0014), indicating that the deletion of RAP1 attenuated C. albicans virulence.
Deletion of RAP1 attenuates C. albicans virulence. (A) Diagram of the experimental design. Nine-week-old BALB/c mice (n = 19) were injected via the tail vein with 1 × 106 C. albicans cells, including wild-type (WT), rap1Δ/Δ mutant, and RAP1-reintegrated strains (RAP1R). The mice kidneys were collected on the second day post-infection for fungal burden and histological examination. The rest of the mice were euthanized at 14 days post-infection. (B) Assessment of the survival rate of mice. The number of surviving mice was plotted against time. (C) Assessment of fungal burden. Mice were sacrificed at 2 days post-infection, and kidneys were collected and homogenized. Homogenates were used for counting the number of colony-forming unit (CFU) by plating on YPD agar plates. Results are expressed as log CFU/g kidney and expressed as mean ± SD from mice infected with each C. albicans strain. The H&E (D) and PAS (E) staining. Upper panel: images of stained kidney samples were captured at 100× magnification. Arrowheads indicate inflammatory foci and highlight leukocyte infiltration in the H&E staining and fungal colonization in the PAS staining. Middle and lower panel: images of stained kidney samples were captured at 400× and 630× magnification. In the lower panel of H&E staining, basophil, lymphocyte, and neutrophil are represented by yellow, green, and red arrowheads, respectively. Scale bars denote magnification. The representative images from five infected mice with similar results are shown.
To determine organ colonization, kidneys were collected. The kidney fungal burden was significantly lower in mice infected with the rap1Δ/Δ mutant than that with the infection of the wild-type and RAP1-reintegrated control strains (Fig. 6C). Histological examinations of kidneys were also performed by hematoxylin and eosin (H&E) staining. The examination demonstrated that kidneys from mice infected with the control strains displayed inflammation with leukocytes, whereas no obvious cell infiltration was observed in that injected with the rap1Δ/Δ mutant and PBS (Fig. 6D). Finally, Periodic acid-Schiff (PAS) staining was also applied to detect fungal invasion (Fig. 6E). In the kidney sections examined, the control cells were observed as filamentous forms surrounded by leukocytes, while little to none of the rap1Δ/Δ mutant was observed. Collectively, these findings indicate the involvement of Rap1 in C. albicans virulence and pathogenicity.
DISCUSSION
Iron is essential but potentially toxic to cause oxidative stress (46). Importantly, the host uses nutritional immunity to limit iron availability to pathogens (6, 7). Thus, maintaining iron balance by regulating iron homeostasis is crucial for both host and pathogen.
Rap1 is a protein that participates in telomere maintenance (22, 28, 47). Notably, C. albicans Rap1 lacks the C-terminal domain implicated in telomere regulation of ScRap1 (28). Moreover, ScRAP1 is an essential gene, whereas RAP1 is not essential for cell viability of C. albicans (28). In addition, our recent studies demonstrated that C. albicans Rap1 regulates cell wall integrity, biofilm formation, and oxidative stress response (29, 48). The rap1Δ/Δ mutant also modulated macrophage-pathogen interaction and showed attenuated virulence in a G. mellonella infection model (29). These results highlighted the significant role of Rap1 in C. albicans pathogenicity and virulence. In this work, we explored other functions of C. albicans Rap1 and found its connection with cellular response to iron.
Several iron-related traits were disclosed to correlate functions of Rap1 with iron response. Although the action of Rap1 under high-iron conditions could not be excluded, RAP1 deletion affected those traits more obviously under conditions of low iron (Fig. 2A through C). A specific example is the strong effect of RAP1 deletion on low-iron-induced flavinogenesis (Fig. 2C). Flavins such as flavin-adenine dinucleotide (FAD), flavin mononucleotide (FMN), and riboflavin (RF) are responsible for electron transfer and redox processes (49, 50). FAD and FMN are essential for forming flavoproteins, which are involved in the respiratory chain and fatty acid oxidation (51, 52). Recently, RF production and its regulation are also considered a novel antifungal drug target (53). Interestingly, RF is involved in ferric iron reduction, thereby promoting iron acquisition (54, 55). Once reduced, flavin-bound reductases release iron from ferric-siderophores, driving iron assimilation (56, 57). Although the detailed mechanisms of inducible flavinogenesis remain unclear, C. albicans flavinogenesis is controlled by different transcription factors in response to iron availability (32, 53, 58–60). For example, Tup1 is a global regulator controlling different features including filamentation and biofilm formation (61–64), whereas Hap43 and Sef1 are required for low-iron response. Interestingly, flavin production was largely decreased and completely abrogated in iron depletion in the TUP1-deleted and HAP43-deleted mutant, respectively (32). In addition, overexpression of SEF1 resulted in higher RF secretion under an iron-replete condition (59), while deletion of SEF1 reduced flavin production under iron deprivation (53). Finally, overexpression of Irf1, a regulator for morphogenesis and iron homeostasis, also led to higher flavin production (60). To determine whether a relationship exists between Rap1 and other regulators in iron-responsive flavinogenesis would be an interesting direction for future research.
The present study also linked Rap1 to the utilization of free iron and its impact on the reductive iron uptake gene expression (Fig. 3A and B). A regulatory circuit consisting of three transcription factors Sef1, Hap43, and Sfu1 has been previously identified to control iron homeostasis in C. albicans (65). Sef1 and Hap43 function under low-iron conditions, whereas Sfu1 is a high-iron regulator (32, 66–69). Sef1 promotes iron acquisition and activates HAP43 expression (66, 67). Reversely, Hap43 represses iron utilization genes and regulates intracellular iron homeostasis (32, 69). Additionally, Hap43 represses Sfu1, which prevents toxicity of iron overload by repressing Sef1 and iron uptake genes (68–70). Therefore, determining whether Rap1 mutually interacts with Sef1, Hap43, and Sfu1 in iron regulation is also intriguing. Furthermore, Rap1 seems to be involved in hemin and hemoglobin utilization. The rap1Δ/Δ mutant exhibited reduced hemin and hemoglobin utilization, as well as reduced hemolysis (Fig. 4A through E). C. albicans secretes hemolytic factors to lyse host RBCs and release iron from hemoglobin (71, 72). Recently, candidalysin is identified as a hemolytic factor of C. albicans (33). Candidalysin is a peptide derived from a precursor protein Ece1 and can form membrane pores to damage host cells (73–75). Interestingly, our RNA-seq data showed an upregulation for the ECE1 expression in the rap1Δ/Δ mutant compared to the wild type in YPD broth (Table S1). Therefore, whether Rap1 can affect candidalysin production in response to iron conditions is of interest for further investigation.
Fluconazole represents one of the first-line anti-Candida drugs (76, 77). Azoles block ergosterol biosynthesis, and complex links between iron and azole susceptibility in Candida species have been reported (19). For example, iron acts as a cofactor for several ergosterol biosynthesis-related enzymes including Erg11 and Erg3, and ergosterol levels are tightly regulated by iron availability (78). Moreover, iron depletion leads to reduced ergosterol production, resulting in higher membrane fluidity and enhanced azole susceptibility (20). Finally, the deletion of the iron permease gene FTR1 makes cells more susceptible to fluconazole (20), and heme transporter gene RBT5 is upregulated in cells treated with fluconazole (79). In this study, the rap1Δ/Δ mutant was more resistant to fluconazole and had lower expression of ERG11 and ERG3, particularly under low-iron condition (Fig. 5A and B). Thus, alterations in membrane lipid phase and asymmetry membrane may occur in this mutant, impacting membrane fluidity and passive diffusion to contribute to azole resistance (80, 81). In addition, the rap1Δ/Δ mutant appears to have higher expression of drug pump genes in both low- and high-iron condition (Fig. 5B). Of note, the rap1Δ/Δ mutant also showed a higher efflux pump activity than the control strains in the low-iron condition (Fig. 6D). Therefore, our findings suggest an intricate association between Rap1, iron, and fluconazole.
Finally, although Rap1 impacts iron homeostasis and virulence in this study, it has also been connected to multiple other functions in C. albicans (27–29, 48). In humans, a correlation exists between iron homeostasis, telomere maintenance, and diseases, involving complex processes such as oxidative stress and DNA damage (82, 83). Interestingly, the role of telomeric and sub-telomeric structures in fungal pathogens has become a focus of recent studies (84). Since Rap1 exerts pleiotropic functions and is involved in telomere maintenance in C. albicans, it is possible that some or all of its observed effects are indirect, potentially involving telomere-associated factors such as genome structure and stability. However, further studies are needed to clarify these mechanisms.
MATERIALS AND METHODS
Strains and growth conditions
C. albicans strains used in this study are listed in Table S3. Cells were routinely maintained at −80°C and plated on YPD plates (1% yeast extract, 2% peptone, 2% glucose, and 1.5% agar). A single colony was inoculated into respective medium and incubated overnight (~16 h) at 30°C with shaking (180 rpm). All reagents were purchased from Sigma-Aldrich (St. Louis, MO, USA) unless indicated otherwise.
RNA-seq analysis
Methods for RNA-seq and its data analysis are included in the supplemental material.
Iron-dependent growth analysis
To assess iron-dependent cell growth, a single colony was inoculated into YPD and grown overnight at 30°C. The overnight cultures were subcultured in YPD containing 400 µM BPS and grown at 30℃ with shaking (180 rpm) for 24 h to deplete intracellular iron content. These iron pre-starved cultures were harvested, washed with sterile double-distilled water (ddH_2_O), and subcultured into high- and low-iron media, respectively. Cells were inoculated in a 96-well flat microplate and incubated at 30℃ for 5 days. Cell growth was measured using a SPECTROstar Nano (BMG Labtech, Ortenberg, Germany).
RNA extraction and reverse transcription real-time qPCR
Iron pre-starved cells were cultured in high- and low-iron media for 5 h at 30℃. Total RNA extraction, RT for cDNA synthesis, and real-time qPCR were performed as previously described (32). The primers used are listed in Table S4. All experiments were performed in duplicate, with at least three biological replicates for each strain, and the relative fold change in gene expression was calculated using the 2^−ΔΔ^CT method (85).
Measurement of intracellular iron level
Intracellular iron level was measured as previously described (86). Iron-prestarved cells were cultured in high- and low-iron media for 5 h at 30℃. To quantify intracellular iron level, cells were resuspended in 200 µL of 70% nitric acid and incubated at room temperature for 24 h, and 800 µL of ddH_2_O was added. The resulting mixture was analyzed using a Thermo Fisher Scientific iCAP TQ ICP-MS. The iron contents were normalized to cell numbers for each sample.
Measurement of flavin production
To measure iron starvation-induced flavin production, NIM was used as previously described (32, 68). The NIM contains 0.17% YNB without Fe and Cu, 0.079% complete supplement mixture, 2% glucose, 0.5% ammonium sulfate, and 0.25 µM CuSO_4_ and is supplemented with 100 µM BPS. Briefly, cells were iron-starved in NIM at 30°C with shaking for 2 days, and cell densities were determined. Then, supernatants were collected, and the fluorescence was measured (emission: 530 nm; excitation: 450 nm). The relative ratio of fluorescence was normalized to the cell densities for each strain. Moreover, the supernatants of each sample were photographed under UV light.
Measurement of hemolytic activity
Blood agar plates were prepared as previously described (36) with some modifications. Briefly, a single colony was inoculated in Sabouraud dextrose broth (1% peptone and 4% glucose, pH 5.6) and incubated at 37℃ overnight. Cells were collected and resuspended in sterile ddH_2_O to a concentration at OD_600_ of ~10. Ten microliters of each diluent was spotted onto sugar-enriched blood agar plates, which were prepared by adding 7 mL of fresh sheep blood into 100 mL Sabouraud dextrose containing 3% (wt/vol) glucose and agar (final pH 5.6 ± 0.2). After incubation at 37℃ in 5% CO_2_ for 24 h, the hemolytic index was calculated using the following formula: hemolytic index = β-hemolysis diameter/colony diameter.
The liquid hemolysis assay was performed as previously described (35) with some modifications. Sheep RBCs and C. albicans cells were mixed in a 1:1 ratio in RPMI 1640 medium (Gibco) and incubated with horizontal shaking (50 rpm) at 37℃ in 5% CO_2_ for 24 h. After centrifugation, the supernatant was transferred to a 96-well microplate to measure absorbance (Abs) at 414 nm. Relative hemolysis was defined as follows: .
Measurement of heme oxygenase activity
Activity of heme oxygenase was determined by measuring biliverdin production as previously described (16). Iron pre-starved cells were harvested, washed, and resuspended in ddH_2_O. Cell suspension was inoculated into YPD broth containing 25 µM hemoglobin and 1 mM BPS. After incubation for 12 h, 6 × 10^9^ cells were collected, resuspended in 1 mL methanol, vortexed for 30 s, and centrifuged (5,000 × g) at room temperature. Biliverdin concentrations were determined by absorbance measurement at 666 nm and calculated using a coefficient of 14,400 M^−1^cm^−1^ (87)
Virulence assay
Methods for virulence assays are included in the supplemental material.
Fluconazole susceptibility testing
Fluconazole susceptibility was determined by agar and broth microdilution assay. Iron-prestarved cells were 10-fold diluted with sterile ddH_2_O and spotted onto YPD agar plates with and without BPS and fluconazole. The plates were incubated at 30℃ for 3 days. Broth microdilution assay was determined according to the Clinical and Laboratory Standards Institute’s M27-A3 guideline (88) with some modifications. Iron-prestarved cells were inoculated into 96-well flat-bottomed plates in YPD medium with or without BPS and different concentrations of fluconazole. After incubation at 30℃ for 24 and 48 h, MIC50 was defined as the drug concentrations capable of reducing cell turbidity by more than 50%.
Measurement of Nile red accumulation
The accumulation of Nile red was determined as previously described (44). Iron-prestarved cells were cultured in high- and low-iron media for 5 h at 30℃. Cells were collected, washed, resuspended in PBS containing 7 µM of Nile red, and incubated at 30°C for 30 min. One hundred microliters of cell suspension was transferred to a black 96-well microplate, and the fluorescence intensity of Nile red was detected using a CLARIOstar Plus microplate reader (BMG Labtech, Ortenberg, Germany).
Statistical analysis
The two-tailed Student’s t-test was used to determine significant differences between samples. Statistical significance was indicated with a P-value < 0.05.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Limon JJ, Skalski JH, Underhill DM. 2017. Commensal fungi in health and disease. Cell Host Microbe 22:156–165. doi:10.1016/j.chom.2017.07.00228799901 PMC 5573128 · doi ↗ · pubmed ↗
- 2Lopes JP, Lionakis MS. 2022. Pathogenesis and virulence of Candida albicans. Virulence 13:89–121. doi:10.1080/21505594.2021.201995034964702 PMC 9728475 · doi ↗ · pubmed ↗
- 3Lockhart SR, Chowdhary A, Gold JAW. 2023. The rapid emergence of antifungal-resistant human-pathogenic fungi. Nat Rev Microbiol 21:818–832. doi:10.1038/s 41579-023-00960-937648790 PMC 10859884 · doi ↗ · pubmed ↗
- 4WHO. 2022. WHO fungal priority pathogens list to guide research, development and public health action. World Health Organization
- 5Fourie R, Kuloyo OO, Mochochoko BM, Albertyn J, Pohl CH. 2018. Iron at the centre of Candida albicans interactions. Front Cell Infect Microbiol 8:185. doi:10.3389/fcimb.2018.0018529922600 PMC 5996042 · doi ↗ · pubmed ↗
- 6Hood MI, Skaar EP. 2012. Nutritional immunity: transition metals at the pathogen-host interface. Nat Rev Microbiol 10:525–537. doi:10.1038/nrmicro 283622796883 PMC 3875331 · doi ↗ · pubmed ↗
- 7Murdoch CC, Skaar EP. 2022. Nutritional immunity: the battle for nutrient metals at the host-pathogen interface. Nat Rev Microbiol 20:657–670. doi:10.1038/s 41579-022-00745-635641670 PMC 9153222 · doi ↗ · pubmed ↗
- 8Almeida RS, Brunke S, Albrecht A, Thewes S, Laue M, Edwards JE, Filler SG, Hube B. 2008. The hyphal-associated adhesin and invasin Als 3 of Candida albicans mediates iron acquisition from host ferritin. P Lo S Pathog 4:e 1000217. doi:10.1371/journal.ppat.100021719023418 PMC 2581891 · doi ↗ · pubmed ↗