KIF2C Deletion Causes Meiotic Abnormalities and Nonobstructive Azoospermia in Mice
Hiroaki Kitakaze, Haruhiko Miyata, Yuki Oyama, Chen Pan, Yuma Kujime, Go Tsujimura, Takahiro Imanaka, Sohei Kuribayashi, Norichika Ueda, Kentaro Takezawa, Shinichiro Fukuhara, Norio Nonomura, Masahito Ikawa

TL;DR
Deleting the KIF2C gene in mice causes infertility and lack of mature sperm, similar to a human condition called nonobstructive azoospermia.
Contribution
This study is the first to show KIF2C's essential role in mouse spermatogenesis and male fertility using global knockout mice.
Findings
Kif2c knockout mice are infertile with severely impaired spermatogenesis.
Mature spermatozoa are absent in Kif2c knockout mice.
Spermatogenic cells in Kif2c knockout mice are arrested during meiosis.
Abstract
Kinesin Family Member 2C (KIF2C) is a key regulator of microtubule dynamics and chromosome segregation in mitosis. However, its role in spermatogenesis remains unclear. Recent transcriptomic analyses suggest a potential link between KIF2C and male infertility. This study aimed to clarify KIF2C's roles in spermatogenesis using Kif2c knockout (KO) mice. To overcome the preweaning lethality associated with Kif2c deletion, we generated Kif2c KO mice with a mixed genetic background of 129X1/SvJ and B6D2. We assessed male fertility, epididymal sperm counts, and testicular sections of Kif2c KO mice. Global Kif2c KO mice were obtained and showed male infertility. Histological analyses and epididymal sperm count revealed that Kif2c KO mice exhibited severely impaired spermatogenesis and absence of mature spermatozoa. These findings are consistent with those observed in patients with…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
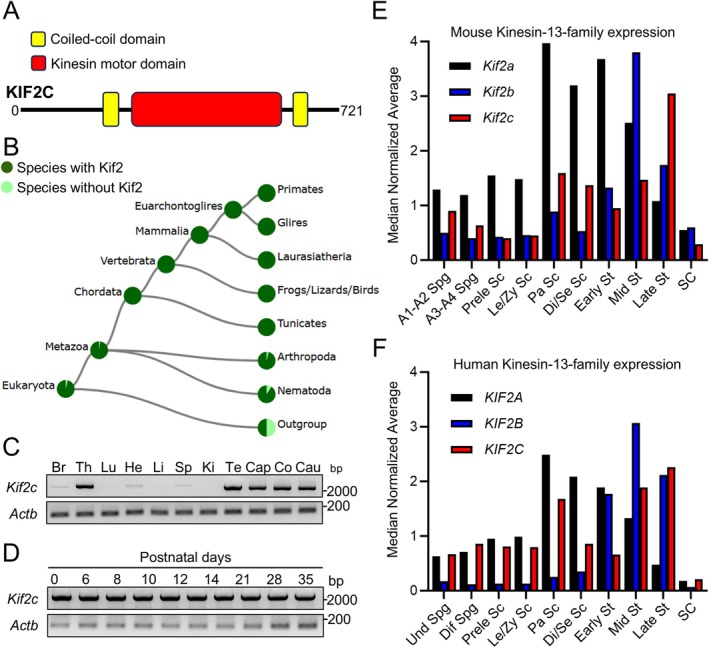 FIGURE 1
FIGURE 1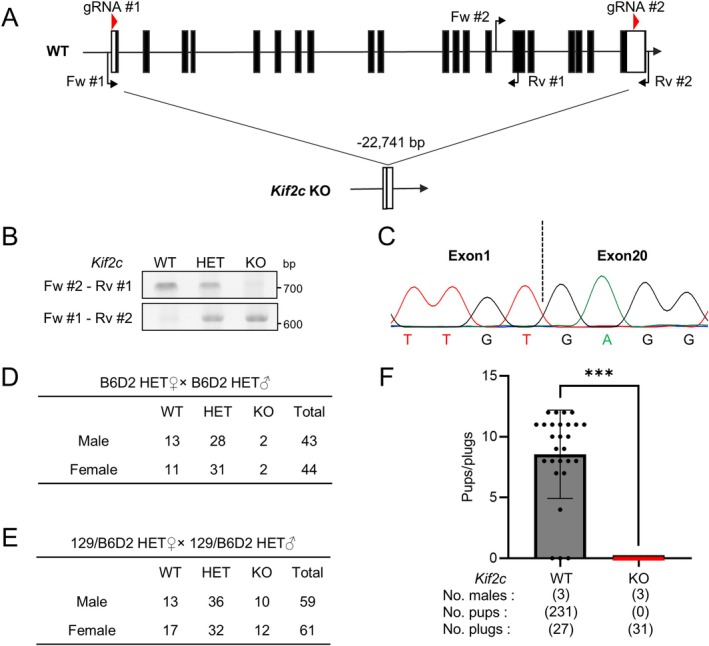 FIGURE 2
FIGURE 2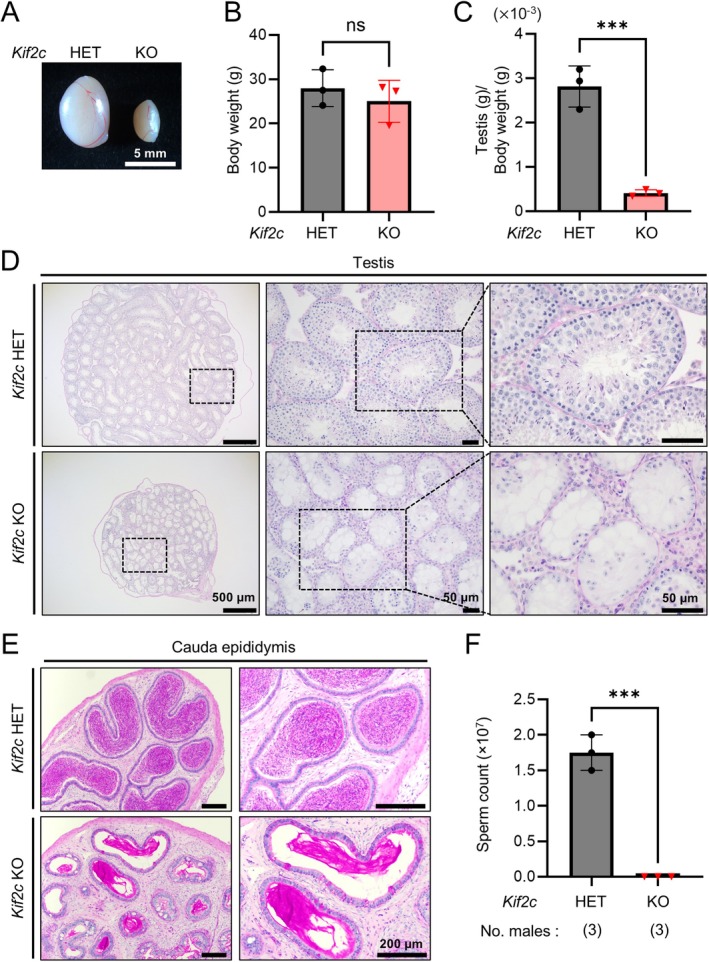 FIGURE 3
FIGURE 3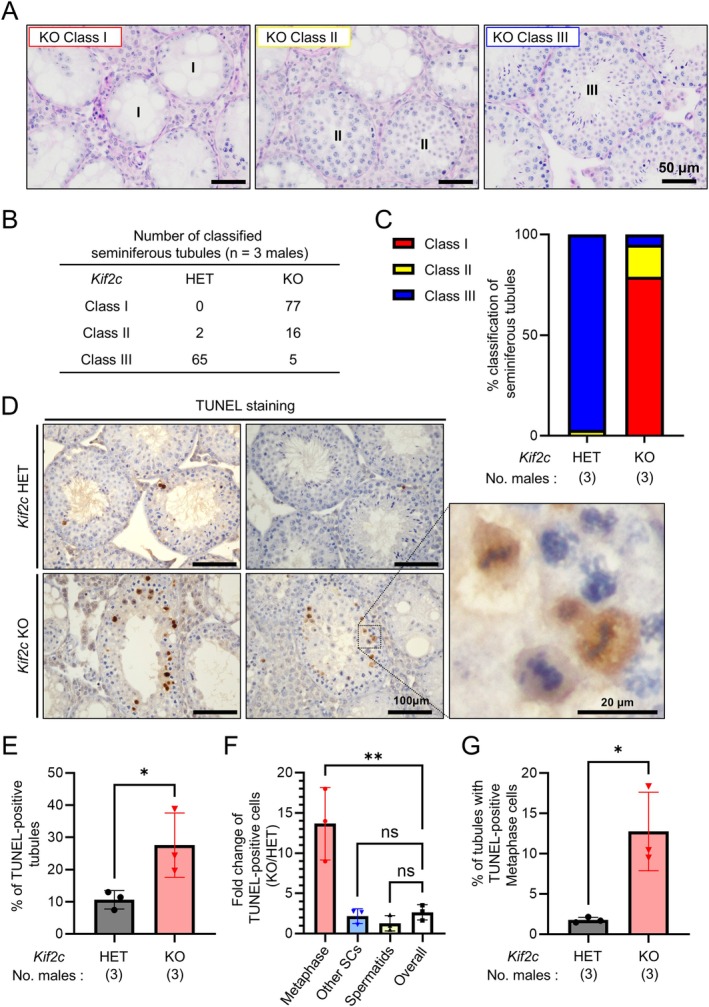 FIGURE 4
FIGURE 4- —Eunice Kennedy Shriver National Institute of Child Health and Human Development 10.13039/100009633
- —Japan Society for the Promotion of Science 10.13039/501100001691
- —Japan Agency for Medical Research and Development 10.13039/100009619
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsMicrotubule and mitosis dynamics · Reproductive Biology and Fertility · Chromosomal and Genetic Variations
Introduction
1
Kinesin is a motor protein involved in various cellular processes [1]. Kinesin Family Member 2C (KIF2C), also known as Mitotic Centromere‐Associated Kinesin (MCAK), is part of the kinesin‐13 family of proteins. In contrast to conventional kinesins, which have the motor domain in the N‐terminus and transport cargos along microtubules, kinesin‐13 family proteins contain the motor domain in the middle of the molecules (Figure 1A) and have the function of depolymerizing microtubules [2, 3]. It has been shown that KIF2C plays a critical role in microtubule dynamics and chromosome segregation in mitosis [4, 5]. Aberrant expression of KIF2C has been observed in various cancers, including breast, lung, gastric, and prostate tumors [6, 7, 8, 9]. In addition, overexpression of KIF2C could lead to chromosomal aberrations that promote tumor progression [10]. Conversely, inhibition of KIF2C has been shown to suppress tumor cell proliferation, suggesting that it may serve as a potential therapeutic target in cancer treatments [11, 12]. While KIF2C has been extensively studied in the context of mitosis and oncogenesis, its roles in meiosis and spermatogenesis are poorly understood.
Kif2c is evolutionarily conserved and expressed in testes. (A) Mouse KIF2C has two coiled‐coil domains and a kinesin motor, catalytic domain. (B) Conservation of Kif2 among different species. (C) RT‐PCR analysis of Kif2c expression in various mouse tissues. Actb was used as a loading control. Br: Brain, Th: Thymus, Lu: Lung, He: Heart, Li: Liver, Sp: Spleen, Ki: Kidney, Te: Testis, Cap: Caput epididymis Co: Corpus epididymis, Cau: Cauda epididymis. (D) RT‐PCR analysis of Kif2c expression in various mouse postnatal testes. Actb was used as a loading control. (E) Expression of mouse kinesin‐13‐family genes in testes was analyzed using single‐cell transcriptome data. A1‐A2 Spg: A1 and A2 differentiating spermatogonia, A3‐A4 Spg: A3 and A4 differentiating spermatogonia, Prele Sc: Preleptotene spermatocytes, Le/Zy Sc: Leptotene/zygotene spermatocytes, Pa Sc: Pachytene spermatocytes, Di/Se Sc: Diplotene/secondary spermatocytes, Early St: Early round spermatids, Mid St: Mid round spermatids, Late St: Late round spermatids, SC: Sertoli cells. (F) Expression of human kinesin‐13‐family genes in testes was analyzed using single‐cell transcriptome data. Und Spg: Undifferentiating spermatogonia, Dif Spg: Differentiating spermatogonia.
Several studies have suggested a potential association between KIF2C and male infertility. Analyses of Gene Expression Omnibus (GEO) datasets from patients with nonobstructive azoospermia show that KIF2C is significantly downregulated in spermatogenesis [13]. Additionally, transcriptomic and single‐cell RNA sequencing analyses have identified KIF2C as one of four novel hub genes linked to Klinefelter syndrome [14], a major cause of azoospermia and oligospermia due to impaired spermatogenesis [15, 16]. It has also been shown that KIF2C is localized to the inner centromere during meiosis in mouse spermatocytes [17]. These findings suggest that KIF2C may be a crucial regulatory gene in male fertility and could serve as a potential biomarker or therapeutic target for male infertility.
Very recently, KIF2C was reported to be essential for male fertility using mice with conditional knockout (cKO) of Kif2c in germ cells [18]; however, it is unclear if global Kif2c KO mice show similar spermatogenetic phenotypes. Although cKO models enable tissue‐specific functional analyses, recombination efficiency is not always perfect, which could complicate the interpretation of phenotypes [19, 20]. According to the International Mouse Phenotyping Consortium (IMPC), Kif2c KO mice show preweaning lethality [21, 22], making it challenging to analyze KIF2C functions using a global KO mouse model. To overcome this limitation, we generated Kif2c KO mice with a mixed genetic background of B6D2 and 129X1/SvJ, and obtained sexually mature mice for analyzing their fertility phenotypes.
Materials and Methods
2
Animals
2.1
All animal experiments were approved by the Animal Care and Use Committee of the Research Institute for Microbial Diseases, The University of Osaka (approval number: #Biken‐AP‐H30‐01 and #Biken‐AP‐R03‐01). Mice were purchased from CLEA Japan (Tokyo, Japan) or Japan SLC (Shizuoka, Japan). Wild‐type (WT) or Kif2c heterozygous (HET) mice were used as controls. Frozen spermatozoa of Kif2c HET mice generated in this study will be made available at the RIKEN BioResource Research Center (Ibaraki, Japan) and the Center for Animal Resources and Development, Kumamoto University (Kumamoto, Japan).
RT‐PCR
2.2
RNA was extracted from multiple adult tissues and postnatal testes of C57BL/6N mice using TRIzol (Thermo Fisher Scientific, MA, USA) according to the manufacturer's protocol. The obtained RNA was reverse transcribed to cDNA with a SuperScript III first‐strand synthesis system (Thermo Fisher Scientific). PCR of Kif2c and Actb was performed using 10 ng of cDNA with KOD Fx Neo DNA Polymerase (Toyobo, Tokyo, Japan) and primers listed in Table S1. The amplification conditions for the PCR were 35 s at 94°C, followed by 30 cycles of 98°C for 10 s, 60°C for 30 s, and 72°C for 2 min 30 s, with a final 3 min extension at 72°C.
In Silico Analyses
2.3
The domain of mouse KIF2C was analyzed using the Simple Modular Architecture Research Tool (SMART) (http://smart.embl‐heidelberg.de/). Single‐cell transcriptome data of Kinesin‐13‐family in the mouse and human testes were obtained from the previously reported database [23]. Kinesin‐13‐family expression was analyzed using the Loupe Browser 7 (10× Genomics, Pleasanton, CA, USA). To compare the amino acid sequences of human KIF2C and mouse KIF2A, 2B, and 2C, BLAST (https://blast.ncbi.nlm.nih.gov/Blast.cgi), Clustal Omega [24], and Jalview [25] were used.
Generation of Kif2c Knockout (KO) Mice
2.4
Kif2c KO mice were generated using the CRISPR/Cas9 system as described previously [26]. Female B6D2F1 mice were super‐ovulated by intraperitoneal injection of CARD HyperOva (Kyudo, Saga, Japan) and human chorionic gonadotropin (hCG) (ASKA Pharmaceutica, Tokyo, Japan). Subsequently, female mice were caged with WT B6D2F1 males. The resulting two‐pronuclear (2PN) zygotes were isolated from the female mice. The sequences of two gRNAs were listed in Table S1, which target exon 1 and exon 20 of Kif2c, respectively. 2PN zygotes were electroporated by super electroporator NEAP21 (NEPA GENE, Chiba, Japan) with the crRNA/tracrRNA/Cas9 ribonucleoproteins (tracrRNA: #TRACRRNA05N‐5NMOL, Sigma‐Aldrich, MO, USA, and CAS9 protein: #A36497, Thermo Fisher Scientific). Treated zygotes were cultured and developed into 2‐cell stage embryos in potassium simplex optimized medium (KSOM) [27], and were transplanted into the oviducts of pseudopregnant ICR female mice the next day. Pups were either naturally delivered or obtained through Cesarean section and genotyped by PCR with the primers listed in Table S1 and subsequent Sanger sequencing of the PCR products.
Mating Tests
2.5
Sexually matured Kif2c WT and KO male mice were housed with three 8‐week‐old B6D2F1 female mice for 3 months, and vaginal plugs and the number of pups were checked and recorded every morning during the first 2 months. Only the number of pups was recorded for the rest of the month.
Sperm Count per Epididymis
2.6
All spermatozoa collected from the cauda epididymis were suspended in a 100 μL drop of TYH medium [28]. After incubation for 10 min at 37°C under 5% CO_2_, the spermatozoa were diluted with water, and the sperm count per epididymis was calculated using a hemocytometer.
Periodic Acid‐Schiff (PAS) Staining
2.7
PAS staining was performed as previously described [29]. Testes or epididymis were fixed at 4°C in Bouin's solution (Polysciences Inc., Warrington, PA, USA) and processed for paraffin embedding. Paraffin sections were cut at a thickness of 5 μm using an HM325 microtome (Microm, Walldorf, Germany). After rehydrating the sections, they were stained with 1% periodic acid (Nacalai Tesque, Kyoto, Japan) and Schiff's reagent (FUJIFILM WakoPure Chemical, Osaka, Japan) for 20 min each at room temperature. The sections were then counterstained with Mayer's hematoxylin solution (FUJIFILM WakoPure Chemical). The sections were observed with an Olympus BX‐53 microscope (Tokyo, Japan).
TUNEL Staining
2.8
Apoptotic cells in seminiferous tubules were detected using the in situ apoptosis detection kit (Takara Bio Inc., Shiga, Japan), which is designed to detect fragmented DNA histochemically by TUNEL [Terminal deoxynucleotidyl Transferase (TdT)‐mediated dUTP Nick End Labeling]. Testes were fixed at 4°C in Bouin's solution (Polysciences Inc.) and processed for paraffin embedding. The paraffin sections were cut to a thickness of 5 μm, and after rehydration, the sections were incubated for 20 min in 10 mM citric acid buffer at pH 6.0 heated to 95°C. To inactivate endogenous peroxidase activity, the sections were incubated in 3% H_2_O_2_ at room temperature for 5 min. After washing three times with PBS, the sections were incubated with TdT enzyme and FITC‐conjugated dUTP at 37°C for 1 h, and then washed with PBS. Next, the sections were incubated with HRP‐conjugated anti‐FITC antibody at 37°C for 30 min, and incubated with ImmPACT DAB substrate (Vector Laboratories, Burlingame, CA, USA) for 2–5 min. After washing with distilled water, the sections were stained with Mayer's hematoxylin solution for 3 min and observed with an Olympus BX‐53 microscope (Tokyo, Japan).
Statistical Analysis
2.9
All statistical analyses were performed using GraphPad Prism 10 (GraphPad Software, San Diego, CA, USA). Data are presented as the mean ± standard deviation (SD) unless otherwise stated. Differences between two groups were analyzed using Student's t‐test. For comparisons among three or more groups, one‐way ANOVA followed by Dunnett's multiple comparison test was performed. Statistical significance was defined as follows: ns (not significant), p < 0.05 (), p < 0.01 (), and p < 0.001 ().
Results
3
Kif2c Is Evolutionarily Conserved and Expressed in Testes
3.1
Mouse KIF2C contains two coiled‐coil domains and a kinesin motor domain (Figure 1A), and is evolutionarily conserved in animals with high homology of amino acid sequences between mice and humans (Figure 1B and Figure S1). To evaluate the tissue expression of mouse Kif2c, we conducted reverse transcription polymerase chain reaction (RT‐PCR) using the primers listed in Table S1. Our results showed that Kif2c is highly expressed in the testis, epididymis, and thymus, while there is also a slight expression detected in the brain, heart, and spleen (Figure 1C). Furthermore, we conducted RT‐PCR on mouse testes at various postnatal days to determine when Kif2c begins to express during the first wave of spermatogenesis and found that Kif2c is expressed even at birth (postnatal Day 0) (Figure 1D). Consistently, the single‐cell transcriptome database [21] indicates that Kif2c is expressed from the early stage of mouse spermatogenesis (Figure 1E). The same database shows that KIF2C is also widely expressed from spermatogonia to spermatids in human spermatogenesis (Figure 1F). In mice and humans, there are three KIF2 proteins, KIF2A‐C. A BLAST search (https://blast.ncbi.nlm.nih.gov/Blast.cgi) shows that the amino acid sequence of mouse KIF2C is similar to that of mouse KIF2A (53% identity) and KIF2B (50% identity). Kif2b/KIF2B is highly expressed in spermatids compared to spermatogonia and spermatocytes, while Kif2a/KIF2A is widely expressed from spermatogonia to spermatids like Kif2c/KIF2C (Figure 1E,F).
Kif2c Deletion Leads to Preweaning Lethality in Mice With the B6D2 Genetic Background, but Crossing With the 129X1/SvJ Strain Mitigates the Lethality
3.2
To explore the function of Kif2c in mouse spermatogenesis, we generated knockout (KO) mice using the CRISPR/Cas9 system to delete exons 1 to 20 of Kif2c in B6D2 mice (Figure 2A). We electroporated 70 zygotes obtained by B6D2F1 x B6D2F1 matings and transplanted 60 embryos into the oviducts of pseudopregnant females. We then obtained eight pups with the confirmation of a large deletion in two pups. These mice were genotyped with genomic PCR using primers shown in Figure 2A and Table S1 (Figure 2B) and subsequent Sanger sequencing of the PCR product (Figure 2C). F0 mice were mated with B6D2F1 WT mice to obtain Kif2c heterozygous (HET) mice. We then crossed B6D2 Kif2c HET mice and obtained a total of 87 pups; however, only 4 of these pups were homozygous for Kif2c KO when genotyped within 1 week after birth (Figure 2D), which is lower than the expected number according to the Mendelian ratio. Additionally, four Kif2c homozygous KO mice died within 10 weeks of birth. These results indicate that Kif2c KO mice with the B6D2 genetic background show preweaning lethality, which is consistent with the IMPC database.
*Deletion of Kif2c results in male infertility in mice. (A) A strategy for generating Kif2c KO mice using CRISPR/Cas9. The gRNA#1 and gRNA#2 were designed to target exon1 and exon 20, respectively, resulting in the deletion of 22,741 bp. Genotyping primers are labeled Fw #1–2 and Rv #1–2. Black rectangle: Translated region, white rectangle: Untranslated region. (B) Genotyping of Kif2c WT, HET, KO mice. Fw #2‐Rv #1 (for WT) and Fw #1‐Rv #2 (for KO) primers shown in (A) were used. (C) A truncation of 22,741 bp was detected in the Kif2c KO allele by Sanger sequencing. (D) The number of pups born from crosses of B6D2 Kif2c HET mice. (E) The number of pups born from crosses of 129/B6D2 Kif2c HET mice. (F) The number of pups born per plug. **p < 0.001, Student's t‐test.
Previous reports have shown that crossing mice with another strain can be an effective strategy to avoid lethality when generating KO mice [30, 31, 32]. In this study, we crossed B6D2 Kif2c HET male mice with 129X1/SvJ WT female mice to generate Kif2c HET mice with the mixed genetic background (129/B6D2). We then crossed 129/B6D2 Kif2c HET mice and obtained 22 homozygous Kif2c KO mice out of 120 pups (Figure 2E). Although the number is less than the expected number (30 pups) according to the Mendelian ratio, KO mice were sexually mature with no obvious abnormalities in appearance or behavior, and subjected to further analyses.
Kif2c Is Indispensable for Male Fertility in Mice
3.3
To investigate the effect of Kif2c deletion on male fertility, we performed mating tests of Kif2c WT or KO male mice with WT females. The average pup number per plug was 8.6 in Kif2c WT males, while no pups were obtained from Kif2c KO males despite observing 31 plugs (Figure 2F). These results indicate that Kif2c is essential for male fertility in mice.
Deletion of Kif2c Causes Nonobstructive Azoospermia With Hypospermatogenesis
3.4
To understand the cause of male infertility, we analyzed the testes of Kif2c KO mice. The gross morphology of the testis was obviously smaller in Kif2c KO mice than that in Kif2c HET mice (Figure 3A). In addition, while there were no significant differences in the body weight between Kif2c HET and KO mice (Figure 3B), the ratio of testis weight to body weight was significantly lower in Kif2c KO mice compared to Kif2c HET mice (Figure 3C).
*Kif2c KO mice show nonobstructive azoospermia with hypospermatogenesis. (A) Gross appearance of testes from adult Kif2c HET and KO mice. (B) Average body weight of adult Kif2c HET and KO mice. n = 3 males, p = 0.8208, Student's t‐test. (C) Average testis weight/body weight of adult Kif2c HET and KO mice. n = 3 males, ***p < 0.001, Student's t‐test. (D) PAS staining of testicular sections of adult Kif2c HET and KO mice. (E) PAS staining of the cauda epididymis of adult Kif2c HET and KO mice. (F) The number of mature spermatozoa obtained from Kif2c HET and KO cauda epididymis. **p < 0.001, Student's t‐test.
To further investigate the cause of small testes, we performed histological analyses of testicular sections from adult Kif2c HET and KO mice (Figure 3D). PAS staining of testicular sections from Kif2c HET mice showed normal spermatogenesis. In contrast, the lumens of the seminiferous tubules in Kif2c KO mice were filled with vacuoles, indicating severe spermatogenesis defects. Furthermore, no mature spermatozoa were present in the cauda epididymis of Kif2c KO mice, while numerous spermatozoa were found in that of Kif2c HET mice (Figure 3E). We also counted the number of spermatozoa collected from the cauda epididymis and found no spermatozoa in Kif2c KO mice (Figure 3F). These results indicate that Kif2c deletion leads to nonobstructive azoospermia associated with hypospermatogenesis.
KIF2C Plays a Crucial Role in Meiosis During Spermatogenesis
3.5
Kif2c KO mice exhibit nonobstructive azoospermia; however, the severity of hypospermatogenesis remains unclear. Mouse spermatogenesis progresses from spermatogonial stem cells through meiosis I and II, ultimately producing haploid spermatocytes [33]. Following the second meiotic division, round spermatids undergo drastic morphological changes called spermiogenesis, transforming into elongated spermatids and spermatozoa [34].
To assess the severity of hypospermatogenesis in Kif2c KO mice, we classified the seminiferous tubules into three categories (Class I to Class III) based on the progression of spermatogenesis as depicted in Figure 4A. Seminiferous tubules were categorized as Class I when only spermatogonia and/or spermatocytes were present. Class II tubules contained round but no elongating spermatids, while Class III tubules contained elongating spermatids. In Kif2c HET mice, almost all seminiferous tubules contained elongating spermatids, with 97% classified as Class III (Figure 4B,C). In contrast, in Kif2c KO mice, 79% of seminiferous tubules were classified as Class I without spermatids in the tubules. In addition, 16% were classified as Class II with round but no elongating spermatids in the tubules, and 5% of the seminiferous tubules were classified as Class III, containing elongating spermatids (Figure 4B,C). These results indicate that spermatogenesis was disrupted before the formation of round spermatids in most of the Kif2c KO seminiferous tubules.
*Kif2c KO male mice exhibited meiotic arrest at metaphase cells. (A) PAS staining of Kif2c KO seminiferous tubules. Seminiferous tubules were classified into three classes. The seminiferous tubules with only spermatogonia and/or spermatocytes were classified as Class I. Class II seminiferous tubules contain round spermatids but no elongating spermatids. Class III seminiferous tubules contain elongating spermatids. (B, C) The number of Class I‐III seminiferous tubules was counted in Kif2c HET and KO mice. (D) The TUNEL staining of Kif2c HET KO testes. Apoptotic cells were stained brown. (E) The percentage of seminiferous tubules containing apoptotic cells was compared between Kif2c HET and KO testes. *p < 0.05, Student's t‐test. (F) TUNEL‐positive cells in Kif2c HET and KO seminiferous tubules were categorized into three groups: Spermatocytes at metaphase, other spermatocytes (SCs), and spermatids. The number of TUNEL‐positive cells in KO divided by the number of TUNEL‐positive cells in HET for each category was calculated. n = 3 males, **p < 0.01, One‐way ANOVA followed by Dunnett's multiple comparison test. (G) The percentage of seminiferous tubules containing TUNEL‐positive spermatocytes at metaphase was compared between Kif2c HET and KO mice. p < 0.05, Student's t‐test.
We also performed TUNEL staining on testicular sections to analyze apoptosis (Figure 4D). We found a significantly higher percentage of TUNEL‐positive seminiferous tubules in Kif2c KO mice compared to Kif2c HET mice (Figure 4E). Furthermore, by investigating the spermatogenic stages of apoptotic cells, we found that apoptosis often occurs during the meiotic metaphase, where the chromosomes are aligned at the center of spermatocytes, in Kif2c KO testes (Figure 4F,G).
Discussion
4
In this study, we generated global Kif2c KO mice by crossing B6D2 mice with the 129 strain and overcame preweaning lethality associated with Kif2c deletion. The successful generation of the global Kif2c KO mouse model enabled us to investigate the roles of KIF2C in spermatogenesis and male fertility. Our results demonstrated that Kif2c KO male mice exhibit meiotic arrest and severe hypospermatogenesis, resulting in nonobstructive azoospermia (NOA). These findings provide evidence that KIF2C is essential for spermatogenesis and male fertility in mice.
Although cKO mice are often used to overcome lethality, CRE‐dependent recombination is not 100% efficient in the cKO approach [19, 20], which can make the interpretation of results difficult. In contrast, our strategy using mixed genetic backgrounds allows us to analyze the effects of gene deletion in all cells throughout the body. However, we should be aware of the potential influence of mixed genetic backgrounds on phenotypes [35]. For Kif2c deletion, the testicular phenotypes of our Kif2c KO mice are consistent with those of Kif2c germ cell‐specific cKO mice [18], indicating that the mixed genetic background does not mitigate the spermatogenetic phenotypes in contrast to the lethality. These phenotypic similarities also suggest that the impaired spermatogenesis observed in our global Kif2c KO mice is likely attributable to the loss of Kif2c in germ cells.
Our histological analysis revealed that the majority of seminiferous tubules were arrested at the early stages of spermatogenesis, particularly during meiosis, with only a small proportion progressing to the stages of round and elongating spermatids. These results suggest that KIF2C is important for the normal completion of meiosis in mouse spermatocytes. In mouse spermatocytes, KIF2C co‐localizes with Aurora B, a key protein in chromosome segregation, in the inner domain of kinetochores in metaphases I and II [17], and it has been suggested that KIF2C ensures proper spindle attachment, corrects erroneous microtubule‐kinetochore interactions, and contributes to spindle formation near the centrosome [36, 37, 38]. These KIF2C functions are consistent with the meiotic arrest phenotypes observed in our Kif2c KO mice.
While KIF2C plays important roles in meiosis, some spermatogenic cells go through meiosis to become round and elongating spermatids without KIF2C. KIF2A, a paralog of KIF2C, may compensate for the KIF2C functions as Kif2a is also expressed in the early stages of spermatogenesis (Figure 1E). Although Kif2a global KO mice showed neonatal lethality [39], Kif2a knockdown in mouse oocytes resulted in metaphase arrest [40, 41], suggesting that KIF2A function may be similar to that of KIF2C. In contrast, Kif2b is highly expressed in spermatids but not in the early stages of spermatogenesis (Figure 1E), and its global KO male mice were fertile [42]. Further analyses are needed to understand the functional differences among KIF2A, 2B, and 2C in mitosis and meiosis.
In conclusion, using global Kif2c KO mice, we reveal that Kif2c deletion results in NOA, which is one of the most severe causes of male infertility without curative treatment available [43, 44]. Global Kif2c KO mice generated in this study may provide a model to study meiotic abnormalities and male infertility, which may lead to a better understanding of the etiology of NOA and the development of new therapeutic strategies.
Ethics Statement
All animal procedures were approved by the Institutional Animal Care and Use Committee at Research Institute for Microbial Diseases, The University of Osaka.
Consent
The authors have nothing to report.
Conflicts of Interest
The authors declare no conflicts of interest.
Supporting information
Data S1. Figure S1. Comparison of KIF2C amino acid sequences between humans and mice. Table S1. Primers and gRNAs used in this study.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1N. Hirokawa , Y. Noda , Y. Tanaka , and S. Niwa , “Kinesin Superfamily Motor Proteins and Intracellular Transport,” Nature Reviews. Molecular Cell Biology 10, no. 10 (2009): 682–696.19773780 10.1038/nrm 2774 · doi ↗ · pubmed ↗
- 2J. G. Ferreira , A. L. Pereira , and H. Maiato , “Microtubule Plus‐End Tracking Proteins and Their Roles in Cell Division,” International Review of Cell and Molecular Biology 309 (2014): 59–140.24529722 10.1016/B 978-0-12-800255-1.00002-8 · doi ↗ · pubmed ↗
- 3N. Hirokawa and Y. Noda , “Intracellular Transport and Kinesin Superfamily Proteins, KI Fs: Structure, Function, and Dynamics,” Physiological Reviews 88, no. 3 (2008): 1089–1118.18626067 10.1152/physrev.00023.2007 · doi ↗ · pubmed ↗
- 4S. F. Bakhoum , S. L. Thompson , A. L. Manning , and D. A. Compton , “Genome Stability Is Ensured by Temporal Control of Kinetochore‐Microtubule Dynamics,” Nature Cell Biology 11, no. 1 (2009): 27–35.19060894 10.1038/ncb 1809 PMC 2614462 · doi ↗ · pubmed ↗
- 5A. L. Manning , N. J. Ganem , S. F. Bakhoum , M. Wagenbach , L. Wordeman , and D. A. Compton , “The Kinesin‐13 Proteins Kif 2a, Kif 2b, and Kif 2c/MCAK Have Distinct Roles During Mitosis in Human Cells,” Molecular Biology of the Cell 18, no. 8 (2007): 2970–2979.17538014 10.1091/mbc.E 07-02-0110 PMC 1949365 · doi ↗ · pubmed ↗
- 6S. Liu , Z. Ye , V. W. Xue , Q. Sun , H. Li , and D. Lu , “KIF 2C Is a Prognostic Biomarker Associated With Immune Cell Infiltration in Breast Cancer,” BMC Cancer 23, no. 1 (2023): 307.37016301 10.1186/s 12885-023-10788-4PMC 10071625 · doi ↗ · pubmed ↗
- 7J. Guo , W. Zhang , L. Sun , et al., “KIF 2C Accelerates the Development of Non‐Small Cell Lung Cancer and Is Suppressed by mi R‐186‐3p via the AKT‐GSK 3β‐β‐Catenin Pathway,” Scientific Reports 13, no. 1 (2023): 7288.37142638 10.1038/s 41598-023-30073-5PMC 10160078 · doi ↗ · pubmed ↗
- 8B. Zhang , P. Liu , Y. Li , et al., “Multi‐Omics Analysis of Kinesin Family Member 2C in Human Tumors: Novel Prognostic Biomarker and Tumor Microenvironment Regulator,” American Journal of Cancer Research 12, no. 11 (2022): 4954–4976.36504885 PMC 9729912 · pubmed ↗