Two new species of Neohelicosporium (Tubeufiaceae, Tubeufiales) from freshwater and terrestrial habitats in China

Abstract
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
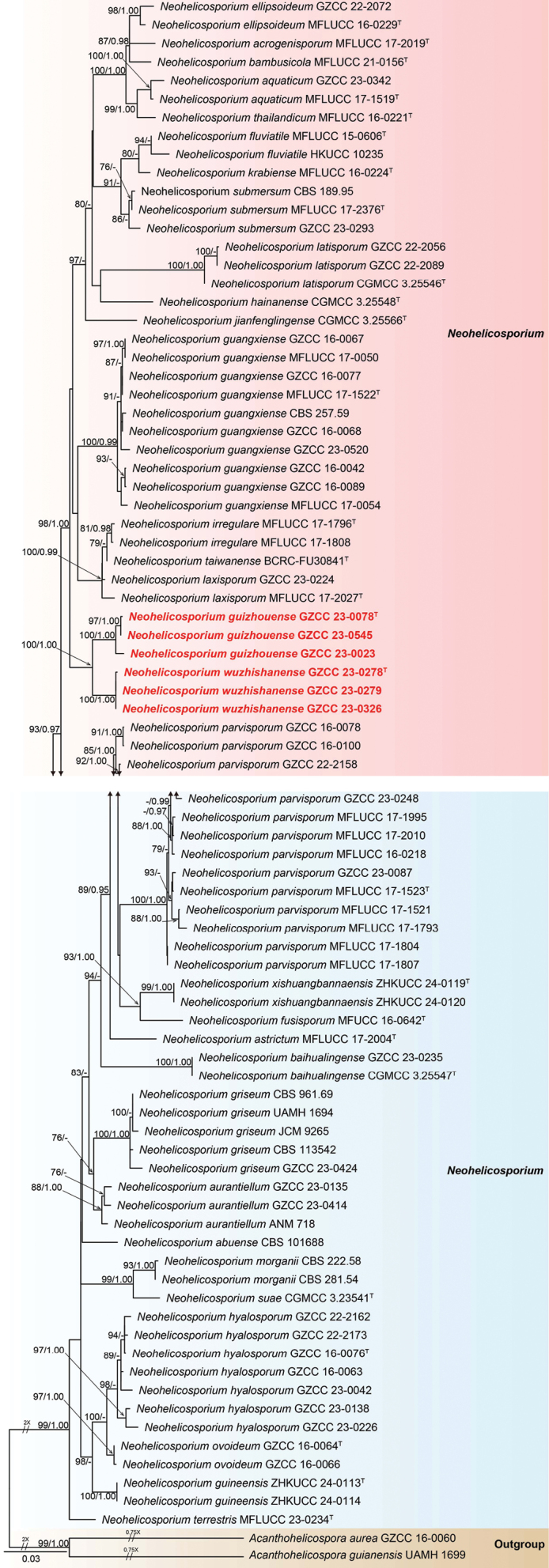 Figure 1
Figure 1 Figure 2
Figure 2 Figure 3
Figure 3| Taxon | Strain | GenBank Accession numbers | |||
|---|---|---|---|---|---|
| LSU | ITS |
| |||
|
|
|
|
|
| |
|
| UAMH 1699 |
|
| – | – |
|
| CBS 101688 | – |
| – | – |
|
| MFLUCC 17-2019T |
|
|
|
|
|
|
|
|
|
| |
|
| MFLUCC 17-1519T |
|
|
|
|
|
| MFLUCC 17-2004T |
|
|
|
|
|
|
|
| – | – | |
|
|
|
| – | – | |
|
| ANM 718 |
|
| – | – |
|
| CGMCC 3.25547T | – |
|
|
|
|
|
|
|
|
| |
|
| MFLUCC 21_0156T |
|
|
|
|
|
|
|
|
| – | |
|
| MFLUCC 16-0229T |
|
|
|
|
|
| MFLUCC 15-0606T |
| – |
|
|
|
| HKUCC 10235 |
| – | – | – |
|
| MFUCC 16-0642T |
|
|
| – |
|
| CBS 961.69 |
|
| – | – |
|
| CBS 113542 |
|
| – | – |
|
| UAMH 1694 |
|
| – | – |
|
|
|
|
| – | |
|
| JCM 9265 |
|
| – | – |
|
|
|
|
|
| |
|
| MFLUCC 17-0054 |
|
|
|
|
|
|
|
|
|
| |
|
|
|
|
|
| |
|
|
|
|
|
| |
|
|
|
|
|
| |
|
| MFLUCC 17-0050 |
|
|
|
|
|
| MFLUCC 17-1522T |
|
|
|
|
|
| CBS 257.59 |
|
| – | – |
|
|
|
|
| – | |
|
| ZHKUCC 24-0113T |
|
|
|
|
|
| ZHKUCC 24-0114 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
| CGMCC 3.25548T |
|
|
|
|
|
|
|
|
|
| |
|
|
|
|
|
| |
|
|
|
|
|
| |
|
|
|
|
|
| |
|
|
| – | – | – | |
|
|
|
|
|
| |
|
|
|
|
|
| |
|
| MFLUCC 17-1796T |
|
|
|
|
|
| MFLUCC 17-1808 |
|
|
|
|
|
| CGMCC 3.25566T |
|
|
|
|
|
| MFLUCC 16-0224T |
|
|
|
|
|
| CGMCC 3.25546T |
|
|
|
|
|
|
|
|
| – | |
|
|
|
|
| – | |
|
|
|
| – | – | |
|
| MFLUCC 17-2027T |
|
|
|
|
|
| CBS 281.54 |
|
| – | – |
|
| CBS 222.58 |
|
| – | – |
|
|
|
|
|
| |
|
|
|
|
|
| |
|
|
|
|
|
| |
|
|
|
|
|
| |
|
| MFLUCC 16-0218 |
|
|
|
|
|
| MFLUCC 17-1523T |
|
|
|
|
|
| MFLUCC 17-1793 |
|
|
|
|
|
| MFLUCC 17-1804 |
|
|
|
|
|
| MFLUCC 17-1807 |
|
|
|
|
|
| MFLUCC 17-1995 |
|
|
|
|
|
| MFLUCC 17-2010 |
|
|
|
|
|
|
|
|
|
| |
|
|
|
| – | – | |
|
|
|
|
|
| |
|
| MFLUCC 17-1521 |
|
|
|
|
|
| CGMCC 3.23541T |
|
|
| – |
|
| CBS 189.95 |
|
| – | – |
|
| MFLUCC 17-2376T |
|
| – | – |
|
|
|
| – | – | |
|
| BCRC-FU30841T | – |
| – | – |
|
| MFLUCC 23-0234T |
|
|
|
|
|
| MFLUCC 16-0221T |
|
|
|
|
|
|
|
|
| – | |
|
|
|
|
| – | – |
|
|
|
|
|
|
|
|
| ZHKUCC 24-0119T |
|
|
|
|
|
| ZHKUCC 24-0120 |
|
|
|
|
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsPlant Pathogens and Fungal Diseases · Plant-Microbe Interactions and Immunity · Mycorrhizal Fungi and Plant Interactions
Introduction
Neohelicosporium was established by Lu et al. (2018a) based on morphological characteristics and phylogenetic evidence. In their study, five new species, Neohelicosporiumaquaticum, N.guangxiense, N.hyalosporum, N.parvisporum (the type species), and N.thailandicum, were collected from submerged decaying wood in southern China and Thailand (Lu et al. 2018a). Jayasiri et al. (2017) reported the first asexual-sexual morph of Neohelicosporiumfusisporum, collected from decaying fruit of a Malvaceae species in Thailand. Subsequently, Lu et al. (2018b) reassessed the order Tubeufiales based on DNA sequence data and morphological characteristics, introducing seven new species and nine new combinations within Neohelicosporium (Morgan 1892; Penzig 1897; Linder 1929; Moore 1954; Rao and Rao 1964; Matsushima 1971, 1980; Rossman 1979, 1987; Chouhan and Panwar 1980; Goos 1985, 1986, 1989; Ho et al. 2001; Tsui et al. 2001, 2006; Cai et al. 2006; Zhao et al. 2007; da Cruz et al. 2009; Promputtha and Miller 2010; Singh and Singh 2016; Jayasiri et al. 2017; Kuo and Goh 2018; Lu et al. 2018a). Dong et al. (2020) introduced another species, Neohelicosporiumsubmersum, collected from a freshwater habitat in Thailand. Subsequent studies reporting novel Neohelicosporium species are listed chronologically: N.suae collected from submerged decaying wood in China (Li et al. 2022); N.bambusicola collected from dead culms of bamboo in Thailand (Tian et al. 2022); N.hainanensis collected from decaying wood in a terrestrial habitat in China (Lu et al. 2022); N.terrestris collected from a dead leaf petiole of Musa sp. in Thailand (Samarakoon et al. 2024); N.guineensis and N.xishuangbannaensis collected from rotting petioles of Elaeisguineensis in China (Xiong et al. 2024); and N.baihualingense, N.hainanense, N.jianfenglingense, and N.latisporum collected from freshwater and terrestrial habitats in China (Ma et al. 2024b).
To date, based on morphological and/or molecular data, Neohelicosporium contains 33 species, with four species (N.myrtacearum, N.nizamabadense, N.sympodiophorum, and N.vesiculiferum) lacking molecular data. Neohelicosporium species are widely distributed across freshwater and terrestrial habitats in Australia, Belgium, Brazil, Canada, China, Cuba, Germany, Honduras, India, Indonesia, Japan, New Zealand, Panama, Peru, the Solomon Islands, South Africa, Thailand, Venezuela, and the USA (Morgan 1892; Penzig 1897; Linder 1929; Moore 1954; Rao and Rao 1964; Matsushima 1971, 1980; Rossman 1979, 1987; Chouhan and Panwar 1980; Goos 1985, 1986, 1989; Ho et al. 2001; Tsui et al. 2001, 2006; Cai et al. 2006; Zhao et al. 2007; da Cruz et al. 2009; Promputtha and Miller 2010; Singh and Singh 2016; Jayasiri et al. 2017; Kuo and Goh 2018; Lu et al. 2018a, 2018b; Dong et al. 2020; Li et al. 2022; Tian et al. 2022; Yang et al. 2023; Zhang et al. 2023a, 2023b; Samarakoon et al. 2024; Ma et al. 2023, 2024b). Among them, 28 Neohelicosporium species exhibit a helicosporous asexual morph, 4 species display both asexual and sexual morphs, and one species (N.terrestris) produces only the sexual morph (Promputtha and Miller 2010; Jayasiri et al. 2017; Lu et al. 2018b; Samarakoon et al. 2024). The asexual morph of Neohelicosporium is characterized by white, light pink, or pale brown colonies; short and solitary conidiophores that are mostly branched and gregarious; mono- to polyblastic conidiogenous cells; and guttulate, hyaline, helicoid conidia (Lu et al. 2018a, 2018b; Dong et al. 2020; Li et al. 2022; Tian et al. 2022; Yang et al. 2023; Ma et al. 2024b). Its sexual morph is depicted by globose to subglobose, spherical or ellipsoidal-ovate, pale brown to yellow or dark brown to black ascomata, 8-spored or 2–4–6-spored, bitunicate asci, and fusiform, hyaline ascospores (Promputtha and Miller 2010; Jayasiri et al. 2017; Lu et al. 2018b; Samarakoon et al. 2024).
In this study, six helicosporous isolates representing two distinct species were collected from freshwater and terrestrial habitats in southern China. Based on comprehensive morphological descriptions and illustrations, as well as multi-gene phylogenetic analyses, two novel species, namely Neohelicosporiumguizhouense and N.wuzhishanense, are introduced.
Materials and methods
Sample collection and specimen examination
Specimens were collected from freshwater and terrestrial habitats between December 2021 and April 2022 in Qiannan Buyi and Miao Autonomous Prefecture, Sandu Shui Autonomous County, Guizhou Province, and Ledong Li Autonomous County and Wuzhishan City, Hainan Province, China, and the important collection details were noted (Rathnayaka et al. 2024). Once the specimens were transported to the laboratory, the specimens from freshwater habitats were incubated at room temperature and maintained in a moist environment for 1–2 weeks. Fungal colonies on the surface of the natural substrates, including conidiophores, conidiogenous cells, and conidia, were examined using a stereomicroscope (SMZ-168, Nikon, Japan). Photographs were taken with an ECLIPSE Ni compound microscope (Nikon, Tokyo, Japan) equipped with a Canon 90D digital camera.
Isolation and material deposition
Single spore isolations were performed following the method described by Senanayake et al. (2020). The germinated coiled conidia were aseptically transferred onto fresh potato dextrose agar (PDA) plates as described by Senanayake et al. (2020). The fungal mycelia were cultured on PDA medium at a constant temperature of 25 °C for 33–41 days. During incubation, colony characteristics such as shape, color, size, margin, and elevation were systematically documented.
Dried fungal specimens were deposited in the Herbarium of Kunming Institute of Botany, Chinese Academy of Sciences (Herb. HKAS), Kunming, China, and the Herbarium of Guizhou Academy of Agriculture Sciences (Herb. GZAAS), Guiyang, China. Pure cultures were deposited at the Guizhou Culture Collection (GZCC), Guiyang, China. Descriptions of the new taxa were uploaded to the Faces of Fungi webpage, as per the guidelines provided by Jayasiri et al. (2015). The new species were registered in the MycoBank database (https://www.mycobank.org/), and MycoBank numbers were obtained.
DNA extraction, PCR amplification, and sequencing
Fresh mycelia were scraped using sterilized toothpicks. Genomic DNA was extracted using the Biospin Fungus Genomic DNA Extraction Kit (BioFlux, China), following the manufacturer’s protocol. Primer pairs LR0R/LR5 (Vilgalys and Hester 1990), ITS5/ITS4 (White et al. 1990), EF1-983F/EF1-2218R (Rehner and Buckley 2005), and fRPB2-5F/fRPB2-7cR (Liu et al. 1999) were used to amplify LSU, ITS, tef1-α, and rpb2 sequence fragments, respectively. The PCR amplification reactions were performed according to the protocol by Ma et al. (2024a). The PCR products were purified and sequenced by Qingke Biotechnology, Chongqing, China.
Phylogenetic analyses
Newly generated sequences were verified and assembled using BioEdit v. 7.0.5.3 (Hall 1999) and SeqMan v. 7.0.0 (Swindell and Plasterer 1997). Additional sequences used in this study were obtained from GenBank (Table 1; https://www.ncbi.nlm.nih.gov/). Alignments for the LSU, ITS, tef1-α, and rpb2 datasets were performed using MAFFT v. 7.473 (https://mafft.cbrc.jp/alignment/server/, Katoh et al. 2019), and were visually checked and converted to the nexus format using AliView v. 1.27 (Daniel et al. 2010; Larsson 2014). Subsequently, trimAl v. 1.2 software was used to trim each gene dataset (Capella-Gutiérrez et al. 2009). The LSU, ITS, tef1-α, and rpb2 datasets were then concatenated using SequenceMatrix v. 1.7.8 (Vaidya et al. 2011).
The maximum likelihood (ML) tree was constructed using the IQ-Tree web server (http://iqtree.cibiv.univie.ac.at/, Nguyen et al. 2015). Bayesian inference (BI) was performed following the methods described by Ma et al. (2022). The best-fit substitution model of the LSU, ITS, tef1-α, and rpb2 datasets was determined using MrModeltest v. 2.3 under the Akaike Information Criterion (AIC) (Nylander et al. 2008).
Phylogenetic trees were visualized using FigTree v. 1.4.4 and edited with Adobe Illustrator CC 2019 v. 23.1.0 (Adobe Systems, USA). Photo-plates were prepared using Adobe Photoshop CC 2019 (Adobe Systems, USA), and measurements were made using the Tarosoft (R) Image Frame Work program v. 1.3.4.
Phylogenetic results
The phylogenetic placements of our six isolates were validated based on a multi-gene phylogenetic analysis incorporating ITS, LSU, tef1-α, and rpb2 sequence data. A total of 84 Tubeufiaceae strains, including isolates obtained in this study and two outgroups, Acanthohelicosporaaurea (GZCC 16-0060) and A.guianensis (UAMH 1699), were analyzed. The concatenated sequence matrix contains 3,386 characters (LSU: 1–842, ITS: 843–1,429, tef1-α: 1,430–2,341, rpb2: 2,342–3,386). Fig. 1 illustrates the best-scoring RAxML tree, with a final likelihood value of -15233.387. Following the guidelines of Chethana et al. (2021), a biphasic approach incorporating both morphological and phylogenetic species concepts was employed to describe the new species.
Phylogenetic tree generated from the RAxML analysis based on the combined dataset of LSU, ITS, tef1-α, and rpb2 sequences. Bootstrap support values for ML (≥ 75%) and posterior probability values (PP) (≥ 0.95) are indicated near respective nodes. Acanthohelicosporaaurea (GZCC 16-0060) and A.guianensis (UAMH 1699) were selected as outgroup taxa. Ex-type strains are indicated by “T”, and newly obtained isolates are in bold red font. A dash (“-”) indicates bootstrap support values below 75% for ML and PP values less than 0.95.
Our collections in our multi-gene phylogenetic tree (Fig. 1) represent two distinct novel Neohelicosporium species within Tubeufiaceae. Our isolates, GZCC 23–0078 and GZCC 23–0545, cluster together as a sister group to GZCC 23–0023, with 100% ML and 1.00 PP support. Additionally, GZCC 23–0278, GZCC 23–0279, and GZCC 23–0326 group together, forming a distinct lineage with N.guizhouense (GZCC 23–0023, GZCC 23–0078, and GZCC 23–0545), supported by 100% ML and 1.00 PP.
Taxonomy
Neohelicosporium
guizhouense
Taxon classificationFungi
X.Y. Ma, J. Ma & Y.Z. Lu sp. nov.
BBAA5E13-348B-5187-8619-BBD3E357F368
903462
Facesoffungi Number: FoF17243
Etymology.
The epithet “guizhouense” refers to Guizhou Province, where the fungus was collected.
Holotype.
HKAS 128908
Description.
Saprobic on decaying wood in terrestrial habitats. Sexual morph Undetermined. Asexual morph Hyphomycetous, helicosporous. Colonies on natural substrate superficial, effuse, gregarious, white. Mycelium mostly immersed, partly superficial, composed of pale brown to brown, branched, septate, guttulate, smooth. Conidiophores 56.5–165 μm long, 4–6.5 μm wide (x̄ = 118.5 × 5.5 μm, n = 20), macronematous, mononematous, procumbent, aggregated, cylindrical, tapering towards the tip, straight or slightly flexuous, branched, septate, smooth- and thick-walled, brown at the base and hyaline towards the apex. Conidiogenous cells 9–16.5 μm long, 3.5–6 μm wide (x̄ = 12.5 × 4.5 μm, n = 30), holoblastic, mono- to poly-blastic, integrated, intercalary or terminal, hyaline, smooth-walled, cylindrical or subcylindrical, becoming truncate towards the apex after conidial secession with tiny tooth-like protrusions, mostly bearing one (rarely two) tiny conidiogenous loci. Conidia solitary, acropleurogenous, helicoid, tapering towards the ends, developing on tooth-like protrusions, 20.5–27.5 μm diam. and conidial filament 3.5–5 μm wide (x̄ = 23.5 × 4 μm, n = 30), 78–109.5 μm long (x̄ = 88 μm, n = 30), indistinctly multi-septate, slightly constricted at septa, tightly coiled up to 3 times, becoming loosely coiled or uncoiled in water, guttulate, hyaline, smooth-walled.
Neohelicosporiumguizhouense (HKAS 128908, holotype) a, b Colonies on the natural substrate c–e Conidiophores and conidiogenous cells f–j Conidiogenous cells k–o Conidia p Germinated conidium q, r Surface and reverse view of colonies on PDA after 33 days of incubation at 25 °C. Scale bars: 50 μm (c); 40 μm (d, e); 10 μm (f–p).
Culture characteristics.
Conidia germinating on PDA within 13 h and germ tubes arising from the terminal ends and the middle of the conidium. Colonies reached 26 mm diam. after 33 days of incubation at 25 °C, irregular, with flat, brown to black mycelia on the surface, in reverse pale brown to black with undulate margin.
Material examined.
China • Guizhou Province, Qiannan Buyi and Miao Autonomous Prefecture, Libo County, Maolan Nature Reserve, on decaying wood in a terrestrial habitat, 10 April 2022, Jian Ma, MN4.1 (HKAS 128908, holotype), ex-type living culture GZCC 23-0078; China, Guizhou Province, Sandu Shui Autonomous County, Yao Renshan National Forest Park, on decaying wood in a terrestrial habitat, 10 April 2022, Jian Ma, YS17.4 (GZAAS 23-0550, paratype), living culture GZCC 23-0545; China, Hainan Province, Ledong Li Autonomous County, Jianfengling National Forest Park, on decaying wood in a forest, 28 December 2021, Jian Ma, JB2 (HKAS 128876, paratype), living culture GZCC 23-0023.
Notes.
In the phylogenetic tree (Fig. 1), three strains of Neohelicosporiumguizhouense (GZCC 23-0078, GZCC 23-0023, and GZCC 23-0545) formed a distinct lineage and were sister to N.wuzhishanense (GZCC 23-0278, GZCC 23-0279, and GZCC 23-0326), supported by 100% ML and 1.00 PP. A comparison of LSU, ITS, tef1-α, and rpb2 sequences between the ex-type strain of N.guizhouense (GZCC 23-0078) and N.wuzhishanense (GZCC 23-0326) revealed 2/837 bp (0.2%, including one gap), 26/773 bp (3.4%, including one gap), 21/885 bp (2.4%, without a gap), and 35/914 bp (3.8%, including one gap) nucleotide base differences, respectively, which strongly support them as two distinct species. Morphologically, Neohelicosporiumguizhouense (HKAS 128908) differs from N.wuzhishanense (HKAS 128903) by its distinct conidiogenous cells (tooth-like vs. tooth-like and/or bladder-like), smaller conidial filaments (3.5–5 μm vs. 4.5–6 μm), and different coiled states in water (loosely coiled vs. tightly coiled). Therefore, we designate the three isolates (GZCC 23-0023, GZCC 23-0078, and GZCC 23-0545) as a new species, Neohelicosporiumguizhouense.
Neohelicosporium
wuzhishanense
Taxon classificationFungi
X.Y. Ma, J. Ma & Y.Z. Lu sp. nov.
5A9891C3-7491-537F-AA99-390117570EC5
903463
Facesoffungi Number: FoF17244
Etymology.
The epithet “wuzhishanense” refers to Wuzhishan City, Hainan Province, where the fungus was collected.
Holotype.
HKAS 128903
Description.
Saprobic on submerged decaying wood in freshwater habitats. Sexual morph Undetermined. Asexual morph Hyphomycetous, helicosporous. Colonies on natural substrate superficial, effuse, solitary, scattered, or gregarious, white to pale brown. Mycelium mostly superficial, partly immersed, composed of pale brown to brown, branched, septate, guttulate, smooth, with mass glistening conidia. Conidiophores 75.5–203 μm long, 5.5–6.5 μm wide (x̄ = 134.5 × 6 μm, n = 25), macronematous, mononematous, erect, solitary, cylindrical, straight or slightly flexuous, occasionally branched, septate, smooth-walled, thick-walled, wider at the base and narrower towards the apex, and brown at the base, becoming hyaline to pale brown towards the apex. Conidiogenous cells 13–29 μm long, 3.5–5 μm wide (x̄ = 19 × 4.5 μm, n = 25), holoblastic, mono- to poly-blastic, integrated, intercalary or terminal, determinate, hyaline to brown, smooth-walled, cylindrical, truncate at the apex after conidial secession, with tiny tooth-like and/or bladder-like protrusions (7–16 μm long, 3–6 μm wide (x̄ = 10.5 × 4.5 μm, n = 15)). Conidia solitary, acropleurogenous, helicoid, tapering toward the ends, developing on tooth-like or bladder-like protrusions, 20.5–28.5 μm diam. and conidial filament 4.5–6 μm wide (x̄ = 25 × 5 μm, n = 30), 92.5–138 μm long (x̄ = 118 μm, n = 30), indistinctly multi-septate, slightly constricted at septa, tightly coiled 2½–3½ times, not becoming loose in water, guttulate, hyaline, smooth-walled.
Neohelicosporiumwuzhishanense (HKAS 128903, holotype) a, b Colonies on the natural substrate c–e, k Conidiophores, conidiogenous cells, and attachment of conidia f–j Conidiogenous cells and attachment of conidia l–p Conidia q Germinated conidium r, s Surface and reverse view of colonies on PDA after 41 days of incubation at 25 °C. Scale bars: 50 μm (c); 40 μm (d, e); 10 μm (f–q).
Culture characteristics.
Conidia germinating on PDA within 18 h and germ tubes arising from the middle of the conidium. Colonies reached 37 mm diam. after 41 days of incubation at 25 °C, irregular, with raised, white to brown mycelia on the surface, in reverse brown to black-brown, with undulate margin.
Material examined.
China • Hainan Province, Wuzhishan City, Wuzhishan National Nature Reserve, on decaying wood in a freshwater stream, 28 December 2021, Jian Ma, WS68 (HKAS 128903, holotype), ex-type living culture GZCC 23-0326; • Ibid., WS19 (GZAAS 23-0282, paratype), living culture GZCC 23-0278; • Ibid., WS20 (GZAAS 23-0283), living culture GZCC 23-0279.
Notes.
Morphologically, Neohelicosporiumwuzhishanense (HKAS 128903) closely resembles N.latisporum (HKAS 128955) in having macronematous, mononematous, erect, cylindrical, septate conidiophores; holoblastic, monoblastic, or polyblastic, integrated, cylindrical conidiogenous cells; and solitary, acrogenous, helicoid conidia (Ma et al. 2024b). However, based on multi-gene phylogenetic analyses, Neohelicosporiumwuzhishanense formed a sister clade with N.guizhouense, which is phylogenetically distant from N.latisporum (Fig. 1). Morphologically, Neohelicosporiumwuzhishanense (HKAS 128903) differs from N.latisporum (HKAS 128955) by its unique conidiogenous cells (tooth-like and/or bladder-like vs. tooth-like) and longer conidia (92.5–138 μm vs. 48.5–67.5) (Ma et al. 2024b). Based on morphological characteristics and DNA sequence data, we identified GZCC 23-0278, GZCC 23-0279, and GZCC 23-0326 as a new species, Neohelicosporiumwuzhishanense.
Discussion
Including the newly described species (N.guizhouense and N.wuzhishanense), the genus Neohelicosporium comprises a total of 35 species (Samarakoon et al. 2024; Ma et al. 2024b). Among these, 15 species have been found in terrestrial habitats, 12 species in freshwater habitats, and eight species have been reported from both freshwater and terrestrial habitats (Morgan 1892; Penzig 1897; Linder 1929; Moore 1954; Rao and Rao 1964; Matsushima 1971, 1980; Rossman 1979, 1987; Chouhan and Panwar 1980; Goos 1985, 1986, 1989; Ho et al. 2001; Tsui et al. 2001, 2006; Cai et al. 2006; Zhao et al. 2007; da Cruz et al. 2009; Promputtha and Miller 2010; Singh and Singh 2016; Jayasiri et al. 2017; Kuo and Goh 2018; Lu et al. 2018a, 2018b; Dong et al. 2020; Li et al. 2022; Tian et al. 2022; Yang et al. 2023; Zhang et al. 2023a, 2023b; Samarakoon et al. 2024; Ma et al. 2023, 2024b). These species occur as saprobes on bamboo, Elaeisguineensis, Malvaceae sp., Musa sp., and decaying wood of unknown origin (Jayasiri et al. 2017; Lu et al. 2018a, b; Tian et al. 2022; Ma et al. 2024a; Samarakoon et al. 2024; Xiong et al. 2024).
Significant intra-species morphological variations have been observed within Neohelicosporium. For example, two collections (HKAS 128940 from submerged decaying wood in China and MFLU 17-1734 from submerged decaying wood in Thailand) represent the same species, Neohelicosporiumsubmersum (Dong et al. 2020; Ma et al. 2024b). However, HKAS 128940 exhibits shorter conidiophores (38–77 µm) compared to MFLU 17-1734 (50–260 µm). Furthermore, the conidiogenous cells in HKAS 128940 are sympodial, a feature not observed in MFLU 17-1734 (Dong et al. 2020; Ma et al. 2024b). Similarly, Neohelicosporiumlaxisporum (MFLU 17-1107 and GZAAS 23-0228), collected from terrestrial habitats in China and Thailand, display notable morphological differences. GZAAS 23-0228 has wider conidiophores (4.5–8 μm vs. 3.5–5 μm) and wider conidiogenous cells (4.5–6 μm vs. 3–4 μm) than MFLU 17-1107 (Lu et al. 2018b; Ma et al. 2024b). Additionally, the conidiogenous cells of MFLU 17-1107 consist of tiny tooth-like protrusions, whereas GZAAS 23-0228 exhibits tiny tooth-like protrusions as well as bladder-like protrusions (Lu et al. 2018b; Ma et al. 2024b). These morphological variations are likely due to geographical differences.
The conidiophores and conidia of some Neohelicosporium species closely resemble those of Parahelicomyces and Tubeufia (Lu et al. 2018b; Li et al. 2022; Ma et al. 2024b). Neohelicosporiumlatisporum (HKAS 128955) resembles Parahelicomycesparvisporus (HKAS 128868) in having erect, solitary conidiophores and wider, tightly coiled, brown conidia (Jayasiri et al. 2017; Kuo and Goh 2018; Lu et al. 2018a, 2018b; Dong et al. 2020; Li et al. 2022; Tian et al. 2022; Yang et al. 2023; Samarakoon et al. 2024; Ma et al. 2023, 2024b). In addition, five Neohelicosporium species, namely N.ellipsoideum (GZAAS 22-2072), N.hainanense (HKAS 128921), N.jianfenglingense (HKAS 128914), N.suae (HKAS 124610), and N.submersum (HKAS 128940), morphologically resemble Tubeufia in having short, simple, erect, irregularly cylindrical conidiophores (Lu et al. 2017, 2018a, 2018b; Li et al. 2022; Ma et al. 2023, 2024b). These morphological similarities highlight the challenges of distinguishing genera based solely on morphology, emphasizing the importance of molecular and phylogenetic analyses for accurate identification and delineation.
Supplementary Material
XML Treatment for Neohelicosporium guizhouense
XML Treatment for Neohelicosporium wuzhishanense
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Cai L Ji KF Hyde KD (2006) Variation between freshwater and terrestrial fungal communities on decaying bamboo culms.Antonie van Leeuwenhoek 89(2): 293–301. 10.1007/s 10482-005-9030-116710640 · doi ↗ · pubmed ↗
- 2Capella-Gutiérrez S Silla-Martínez JM Gabaldón T (2009) trim Al: A tool for automated alignment trimming in large-scale phylogenetic analyses.Bioinformatics (Oxford, England)25(15): 1972–1973. 10.1093/bioinformatics/btp 34819505945 PMC 2712344 · doi ↗ · pubmed ↗
- 3Chethana KT Manawasinghe IS Hurdeal V Bhunjun CS Appadoo M Gentekaki E RaspéO Promputtha I Hyde KD (2021) What are fungal species and how to delineate them? Fungal Diversity 109(1): 1–25. 10.1007/s 13225-021-00483-9 · doi ↗
- 4Chouhan J Panwar K (1980) Hyphomycetes of Mount Abu-V.Indian Phytopathology 33: 285–291.
- 5da Cruz AC Rdos Santos Santa Izabel T Leão-Ferreira SM Gusmão LFP (2009) Conidial fungi from the semi-arid Caatinga biome of Brazil. New species and new records of Helicosporium.Mycotaxon 110(1): 53–64. 10.5248/110.53 · doi ↗
- 6Daniel GP Daniel GB Miguel RJ Florentino FR David P (2010) ALTER: Program-oriented conversion of DNA and protein alignments. Nucleic Acids Research 38: W 14–W 18. 10.1093/nar/gkq 321PMC 289612820439312 · doi ↗ · pubmed ↗
- 7Dong W Wang B Hyde KD Mc Kenzie EHC Raja HA Tanaka K Abdel-Wahab MA Abdel-Aziz FA Doilom M Phookamsak R Hongsanan S Wanasinghe DN Yu XD Wang GN Yang H Yang J Thambugala KM Tian Q Luo ZL Yang JB Miller AN Fournier J Boonmee S Hu DM Nalumpang S Zhang H (2020) Freshwater Dothideomycetes.Fungal Diversity 105(1): 319–575. 10.1007/s 13225-020-00463-5 · doi ↗
- 8Goos R (1985) A review of the anamorph genus Helicomyces. Mycologia 77(4): 606–618. 10.1080/00275514.1985.12025146 · doi ↗