Auxin! here you go again: Spatiotemporal dynamic regulation of auxin promotes proper nodule formation in Medicago truncatula
Gunjan Sharma, Héctor H Torres-Martínez

Abstract
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
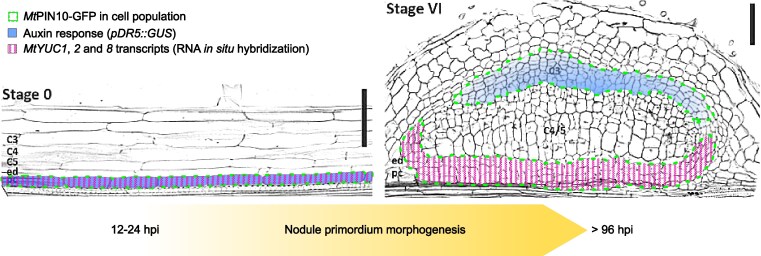 Figure 1
Figure 1Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsPlant Disease Resistance and Genetics · Legume Nitrogen Fixing Symbiosis · Plant Pathogens and Resistance
Legumes such as peas, pulses, and soybeans are a primary source of proteins. Legumes have a unique property to produce knob-like structures called nodules on their roots after infection by nitrogen-fixing bacteria, rhizobia. Legumes and rhizobia form a mutual symbiotic relationship in which plants provide nutrients and rhizobia provide nitrates to plants by fixing atmospheric nitrogen (Soltis et al. 1995). The symbiotic relationship between legumes and rhizobia is crucial and beneficial to reduce the use of nitrogen fertilizers (Goyal et al. 2021). Therefore, understanding the process of symbiosis and nodule formation in different legume species is of great interest.
Nodulation requires a sophisticated array of coordinated cellular processes in specific root cells involving an interplay of plant hormones (Lin et al. 2020; Liu et al. 2023). A pioneering study reported that the auxin influx transporter proteins AUXIN RESISTANT 1 (AUX1) and LIKE-AUX1 (LAX) are expressed in nodule primordia (de Billy et al. 2001). Auxins are known to essentially regulate all plant developmental pathways (Teale et al. 2006). It is no surprise that auxin promotes root nodule development through cell cycle control and vascular tissue specialization for transporting water and nutrients (Kohlen et al. 2018; Lin et al. 2020).
A growing body of evidence has further strengthened the idea that differential auxin distribution is necessary to induce nodule development. However, how the auxin responsive gene network shapes the formation and maintenance of precise auxin concentrations is still not fully understood.
In a recently published article in Plant Physiology, Xiao et al. (2025) have dissected the expression pattern of auxin biosynthesis, YUCCA (YUC), and transport PIN-formed (PIN) genes during nodule primordium initiation and development. Authors revealed that auxin is present even before the start of cell division in pericycle cells and its concentration is dynamically maintained in subsequent stages of root nodule primordium organogenesis. Interestingly, auxin is mainly synthesized at the site of nodule primordia development instead of long-distance transport from shoots to roots, known as acropetal transport.
To investigate the auxin gradient during nodule primordium initiation and development, the authors introduced an artificial auxin-responsive DR5-GUS reporter into Medicago truncatula plants as a proxy for observing the auxin concentration gradient (Ulmasov et al. 1997). Medicago plants expressing DR5-GUS reporter were subjected to infection with rhizobia, and the formation of nodule primordia and auxin gradients was observed from 12 to 96 h post infection. Progression of nodule primordium development was divided into progressive stages from 0 to VI based on the initiation of cell divisions in pericycle, cortical, and nodule meristem cells. Interestingly, DR5-GUS was expressed in stage 0 in 6 to 8 pericycle cells before the onset of divisions (Figure). The auxin reporter was visible in subsequent stages (I to VI) in dividing pericycle, endodermis, cortex, nodule primordium, vasculature, and nodule meristem. However, DR5-GUS expression was dramatically reduced in inner cortical derived layers during later stages, and outer cortical derived layers exhibited auxin response maxima potentially for nodule meristem formation (Figure).
To further explore the auxin distribution in nodule primordium development, the authors employed the in situ hybridization technique to observe the expression of known bona fide auxin biosynthetic YUC genes in fixed Medicago roots that have been inoculated with rhizobia. Expression of the YUC genes coincided with DR5-GUS reporter in rhizobia-inoculated roots during nodule stage 0 in pericycle cells and in developing nodule vasculature during later stages (Figure). Surprisingly, these YUC genes were not expressed in cortical cell layers in contrast to the observed DR5 expression.
To address the potential auxin transport from pericycle to cortical cell layers, Xiao et al. (2025) studied the expression of Medicago auxin export carrier PINs and influx carrier LAX2 using in situ hybridization approach. Authors revealed nodule primordia stage–specific expression patterns of MtPINs and MtLAX2 facilitating the formation of auxin maxima in pericycle and cortical cell derivatives throughout nodule organogenesis. The authors' claim of auxin transport from pericycle to cortical cells was further strengthened by subcellular localization of GFP-tagged MtPIN10 transporter protein in stages II to IV of nodule primordium development. In later stages (V to VI), MtPIN10 expression was predominant in cortex derivatives associated with nodule meristem (Figure).
To further investigate the role of YUCs and PIN proteins in precise auxin distribution, authors performed a combinatorial approach using RNA interference and YUC inhibitor 4-phenoxyphenylboronic acid to reduce the MtYUCs and MtPIN expression/activity. The treatment of roots with 4-phenoxyphenylboronic acid before rhizobia infection severely reduced the root nodule formation. A simultaneous downregulation of MtYUC members in constitutive and pericycle-specific manner reduced root nodule formation between 80% and 60%. Similarly, Medicago PINi (an RNAi construct to reduce MtPIN2, 4, and 10 expressions simultaneously) plants exhibited significantly reduced nodule numbers. Interestingly, pericycle- and symbiosis-specific downregulation of PINs substantially reduced nodule number along with poorly developed nodule meristem. These observations establish the necessity of the precise auxin dosage and transportation in formation of root nodules.
In conclusion, Xiao et al. (2025) have improved our understanding of how auxin gradient is established through a well-orchestrated mechanism of auxin transportation and local synthesis during nodule primordium development. Interestingly, auxin, like other phytohormones, is also transported through plasmodesmata (Tee and Faulkner 2024). It would be intriguing to further elucidate the roles of plasmodesmata-mediated transport of auxin in nodule development. Collectively, the presented study has created a landscape of spatiotemporal auxin gradient determinants of nodule development in the model legume Medicago. This provides a foundation for further studies aiming to understand atmospheric nitrogen fixation in plants through rhizobia, with a goal to reduce the use of nitrogen fertilizers.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1de Billy F, Grosjean C, May S, Bennett M, Cullimore JV. Expression studies on AUX 1-like genes in Medicago truncatula suggest that auxin is required at two steps in early nodule development. Mol Plant Microbe Interact. 2001:14(3):267–277. 10.1094/MPMI.2001.14.3.26711277424 · doi ↗ · pubmed ↗
- 2Goyal RK, Mattoo AK, Schmidt MA. Rhizobial-host interactions and symbiotic nitrogen fixation in legume crops toward agriculture sustainability. Front Microbiol. 2021:12:669404. 10.3389/fmicb.2021.66940434177848 PMC 8226219 · doi ↗ · pubmed ↗
- 3Kohlen W, Ng JLP, Deinum EE, Mathesius U. Auxin transport, metabolism, and signalling during nodule initiation: indeterminate and determinate nodules. J Exp Bot. 2018:69(2):229–244. 10.1093/jxb/erx 30828992078 · doi ↗ · pubmed ↗
- 4Lin J, Frank M, Reid D. No home without hormones: how plant hormones control legume nodule organogenesis. Plant Commun. 2020:1(5):100104. 10.1016/j.xplc.2020.10010433367261 PMC 7747975 · doi ↗ · pubmed ↗
- 5Liu Z, Yang J, Long Y, Zhang C, Wang D, Zhang X, Dong W, Zhao L, Liu C, Zhai J, et al Single-nucleus transcriptomes reveal spatiotemporal symbiotic perception and early response in Medicago. Nat Plants. 2023:9(10):1734–1748. 10.1038/s 41477-023-01524-837749242 · doi ↗ · pubmed ↗
- 6Soltis DE, Soltis PS, Morgan DR, Swensen SM, Mullin BC, Dowd JM, Martin PG. Chloroplast gene sequence data suggest a single origin of the predisposition for symbiotic nitrogen fixation in angiosperms. Proc Natl Acad Sci U S A. 1995:92(7):2647–2651. 10.1073/pnas.92.7.26477708699 PMC 42275 · doi ↗ · pubmed ↗
- 7Teale WD, Paponov IA, Palme K. Auxin in action: signalling, transport and the control of plant growth and development. Nat Rev Mol Cell Biol. 2006:7(11):847–859. 10.1038/nrm 202016990790 · doi ↗ · pubmed ↗
- 8Tee EE, Faulkner C. Plasmodesmata and intercellular molecular traffic control. New Phytol. 2024:243(1):32–47. 10.1111/nph.1966638494438 · doi ↗ · pubmed ↗