UCK2-dependent conversion of cytidine to CTP is required for CTP uptake by Chlamydia trachomatis
Laure Blanchet, Agathe Subtil

TL;DR
The study shows that Chlamydia trachomatis needs host enzyme UCK2 to convert cytidine into CTP before it can be taken up by the bacteria.
Contribution
The study reveals that cytidine must be phosphorylated by host UCK2 for bacterial nucleic acid synthesis.
Findings
Cytidine is phosphorylated by host UCK2 before being incorporated into bacterial nucleic acids.
Nucleosides are likely converted into nucleotides in the host cytoplasm before entering the inclusion lumen.
Abstract
Chlamydia trachomatis , an obligate intracellular bacterium, develops into a vacuolar compartment called the inclusion. The bacteria import nucleoside triphosphates (NTPs) present in the inclusion lumen. It remains unclear whether nucleosides enter the inclusion lumen in their native form or as phosphorylated nucleotides. Using click chemistry coupled with fluorescence microscopy we provide evidence that cytidine requires phosphorylation by host uridine-cytidine kinase 2 (UCK2) prior to its incorporation into bacterial nucleic acids. These findings support the hypothesis that nucleosides are converted into nucleotides in the host cytoplasm prior to translocation into the inclusion lumen. Future work should therefore focus on the identification of nucleotide transporter(s) at the inclusion membrane.
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
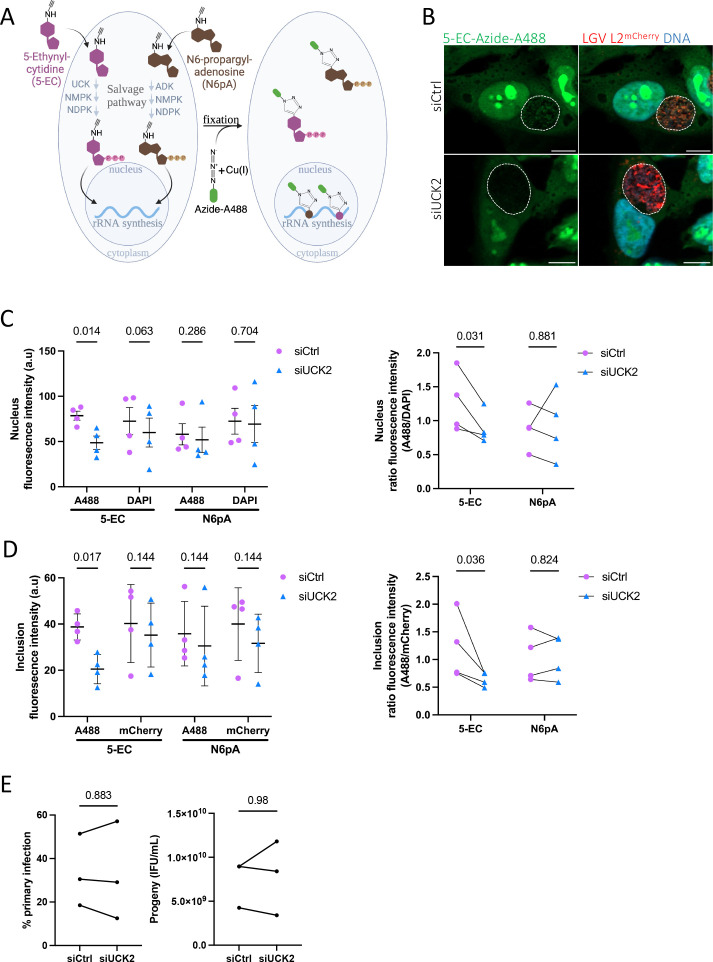 Figure 1
Figure 1|
|
|
|
|
|
10 mM in DMSO |
#CLK-N004-1, Jenabioscience |
|
|
100 mM in DMSO |
#CLK-087, Jenabioscience |
|
|
50 mM in DMSO |
#CLK-1275, Jenabioscience |
|
|
10 mM in H 2 O |
#209198, Sigma Aldrich |
|
|
250 mM in H 2 O |
#A7631, Sigma Aldrich |
|
|
100 mM in DMSO |
#762342, Sigma Aldrich |
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsBiochemical and Molecular Research · RNA modifications and cancer · Peptidase Inhibition and Analysis
Description
Chlamydia trachomatis is an obligate intracellular bacterium that infects human epithelial cells of the urogenital tract and of the ocular conjunctiva. It is the first cause of female infertility and of blindness of bacterial origin, representing a significant public health burden (Taylor, Burton, Haddad, West, & Wright, 2014; Van Gerwen, Muzny, & Marrazzo, 2022).
C. trachomatis develops within a membrane-bound compartment called the inclusion, located in the host cell cytoplasm. Upon co-evolution with its host C. trachomatis has lost genes for several biosynthetic pathways and has become dependent on the host for multiple metabolites (Stephens et al., 1998). Notably, the bacteria do not synthesize their own nucleoside triphosphates (NTPs), the building blocks of RNA and DNA (in the form of deoxynucleotides, dNTPs, for the latter), and acquire them from the host (Tipples & McClarty, 1993). Nucleosides and their phosphorylated derivatives, nucleotides, are transported across membranes only via specialized transporters. In mammalian cells, nucleosides, but not nucleotides, are imported from the extracellular environment through concentrative nucleoside transporters (CNTs) and equilibrative nucleoside transporters (ENTs) (Young, 2016). Once inside the cell, nucleosides are converted into NTP through the salvage pathway. C. trachomatis express two bacterial surface transporters, Npt1 and Npt2, that enable the uptake of NTPs, but not of nucleosides (Tjaden et al., 1999). The process by which host-derived NTPs become available for bacterial uptake within the inclusion lumen is unknown. This work aimed at testing the hypothesis that nucleosides needed to be converted to nucleotides before translocation into the inclusion lumen.
Click chemistry is commonly used to track the dynamics of nucleotide integration into DNA and RNA of eukaryotes (Fantoni, El-Sagheer, & Brown, 2021; Jao & Salic, 2008). Incorporation of nucleoside derivatives in nucleic acid polymers requires their conversion into nucleotides by dedicated enzymes. For instance, the cytidine derivative 5-ethynyl-cytidine (5-EC) needs to be converted into CMP by the uridine-cytidine kinase 2 (UCK2) before incorporation into RNA. It does not incorporate into DNA, likely because it is a poor substrate for the ribonucleotide reductase (Qu et al., 2013). We reasoned that if the conversion of 5-EC into CMP occurred prior to import in the inclusion, silencing UCK2 should decrease the incorporation of 5-EC derived nucleotide in bacteria. Alternatively, if nucleosides were transported across the inclusion membrane, and conversion of 5-EC into CMP occurred in the inclusion lumen, silencing UCK2 should not affect the 5-EC-derived signal associated with the bacteria. UCK2 expression was silenced using siRNA in HeLa cells before infection with a constitutively mCherry-expressing strain of C. trachomatis (LGV L2 ^mCherry^ ). At 16 hours post infection (hpi), 5-EC or N6pA (N6-propargyl-adenosine, a clickable analog of adenosine used as a negative control), were added to the medium culture for six hours, the samples were then fixed and processed for fluorescence microscopy, with the covalent fixation of a fluorophore to the probes (Figure 1.A). Fluorescence intensity of the probes was quantified in individual nuclei and inclusions. Fluorescence from DAPI and mCherry was used to estimate nuclear DNA content and bacterial load within inclusions, respectively. In the nucleus, the signal of nucleoside analogs mainly accumulated in structures corresponding to nucleoli, site of rRNA synthesis (Figure 1.B). Silencing of UCK2 led to a 50% decrease of this nuclear signal for the cytosine derivative (Figure 1.B-C), while the nuclear signal from the adenosine derivative remained stable (Figure 1.C). This observation indicated that, as expected, silencing UCK2 reduced the conversion of 5-EC into CTP in the host. The 5-EC derived signal was also observed in the inclusions as dots overlapping with bacteria (mCherry positive, Fig. 1B ). This finding suggests that the salvage pathway contributes to the nucleotide pool exploited by the bacteria, and that click chemistry is sensitive enough to detect NTP incorporation in Chlamydia . A 50% decrease in the 5-EC fluorescence signal in the inclusions was observed upon UCK 2 silencing (Figure 1.B and D), while the N6pA derived signal remained stable (Figure 1.D). This indicates that 5-EC is converted to CTP by UCK2 before being transported into the inclusion lumen. However, we cannot formally exclude that UCK2 itself is translocated inside the inclusion lumen, where the conversion could occur. Finally, we assessed whether C. trachomatis could complete its developmental cycle under these conditions. Reinfection assays revealed that UCK2 silencing did not affect bacterial development (Figure 1.E). Thus, UCK2 depletion did not impair bacterial growth while significantly decreasing incorporation of 5-EC derived CTP in the bacteria. This implies that alternative source(s) of CTP compensate for the decrease in cytidine to CTP conversion upon UCK2 silencing. De novo CTP biosynthesis in the host is likely solicited. The salvage pathway of UMP biosynthesis from uracil and phosphoribosyl pyrophosphate (PRPP) may also contribute, as C. trachomatis possesses a CTP synthase that can convert UTP into CTP (Wylie, Wang, Tipples, & McClarty, 1996).
In conclusion, our data indicate that the salvage pathway contributes to the nucleotide pools hijacked by the bacteria and that cytidine is converted to CTP before its translocation into the inclusion lumen. This implies the existence of NTP transporter(s) in the inclusion membrane, which could originate from the host or from the bacteria. Interestingly, several enzymes of purine biosynthesis were found in proximity to the inclusion membrane (Olson et al., 2019), indicating that de novo nucleotide biosynthesis might be manipulated by the bacteria to feed these elusive NTP transporters.
Methods
Cells and bacteria
HeLa cells (ATCC) were grown in Dulbecco’s modified Eagle’s medium with Glutamax (Invitrogen), supplemented with 10 % (v/v) heat-inactivated fetal bovine serum and maintained at 37 °C, in 5 % CO 2 atmosphere. *C. trachomatis * serovar LGV L2 strain 434 (obtained from ATCC) stably expressing mCherry (click chemistry) or the green fluorescent protein (progeny) were used (Agaisse & Derré, 2013). Bacteria were stored in sucrose-phosphate-glutamic acid buffer (SPG: 10 mM sodium phosphate [8 mM Na2HPO4- 2 mM NaH2PO4], 220 mM sucrose, 0.50 mM l-glutamic acid) at -80 °C (Scidmore, 2005; Vromman, Laverriere, Perrinet, Dufour, & Subtil, 2014).
siRNA treatment
100 000 HeLa cells were seeded in a 24-well plate. Eight hours later the cells were treated with a mix containing Lipofectamine RNAiMAX (Invitrogen) and 10 nM of control siRNAs (siCtrl, #SR-CL000-005) or siUCK2 (5′-GGGAUCUUGAGCAGAUUUUtt-3′) purchased from Eurogentec (Belgium), following the manufacturer’s recommendation. A second siRNA transfection was conducted 48 h after the first transfection for click chemistry assay or 2 h post infection for progeny assay. The silencing efficiency was confirmed by RT-qPCR 72 hours after the first siRNA treatment.
Click chemistry
30 000 cells treated twice, 72 h and 24 h earlier, with siRNA were seeded on coverslips in a 24-well plate. Eight hours later the cells were infected with LGV L2 ^mCherry^ (MOI = 1). Sixteen hpi 5 µM N6pA or 10 µM 5-EC were added to the culture medium and 6 hours later cells were washed and fixed in 4 % paraformaldehyde (w:v), 4 % (w:v) sucrose in PBS for 20 min. Cells were incubated for 10 min in 50 mM NH 4 Cl in PBS and permeabilized in 0.3 % Triton X-100 in PBS for 10 min prior incubation for 1 h in a 25 µl drop containing 1 mM copper sulfate; 2.5 mM THPTA; 20 µM Azide-AF488; 12.5 mM acid ascorbic; and 0.5 µg/mL DAPI in PBS. Coverslips were mounted on slides with Mowiol (Sigma-Aldrich). Images were obtained on an Axio observer Z1 microscope equipped with an ApoTome module (Zeiss, Germany) 63× Apochromat lens. Images were taken with an ORCAflash4.OLT camera (Hamamatsu, Japan) using the Zen software from Zeiss. Images were analyzed with the Fiji software. The intensity fluorescence of the A488 fluorophore, DAPI, and mCherry were measured in the inclusion and the corresponding nuclei for approximately 50 cells per condition.
Progeny assay
100 000 HeLa cells were treated with siRNA in duplicate wells and infected 48 h later with LGV L2 ^GFP^ (MOI = 0.3) and a second siRNA transfection was performed 2 hpi. Twenty-four hpi cells were collected and the percent of primary infection was quantified by flow cytometry. Forty-eight hpi, bacteria from the duplicate well were collected by breaking the cells and used to reinfect in serial dilutions fresh HeLa cells plated in a 24-well plate. The next day, cells were collected from three wells infected at less than 30 % (estimated by visual inspection), the infection rate was determined by flow cytometry to deduce the number of IFUs present at 48 hpi. 20 000-30 000 events per samples were acquired on a CytoFLEX S (Beckman Coulter). Analysis was performed using FlowJo (version 10.0.7).
Reagents
**: **
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Agaisse Hervé DerréIsabelle 2013218 A C. trachomatis Cloning Vector and the Generation of C. trachomatis Strains Expressing Fluorescent Proteins under the Control of a C. trachomatis Promoter P Lo S ONE 821932-6203 e 57090 e 5709010.1371/journal.pone.005709023441233 PMC 3575495 · doi ↗ · pubmed ↗
- 2Fantoni Nicolò Zuin El-Sagheer Afaf H. Brown Tom 2021114 A Hitchhiker’s Guide to Click-Chemistry with Nucleic Acids Chemical Reviews 121120009-26657122715410.1021/acs.chemrev.0c 0092833443411 · doi ↗ · pubmed ↗
- 3Jao Cindy Y. Salic Adrian 20081014 Exploring RNA transcription and turnover in vivo by using click chemistry Proceedings of the National Academy of Sciences 105410027-8424157791578410.1073/pnas.0808480105 PMC 257291718840688 · doi ↗ · pubmed ↗
- 4Olson Macy G. Widner Ray E. Jorgenson Lisa M. Lawrence Alyssa Lagundzin Dragana Woods Nicholas T. Ouellette Scot P. Rucks Elizabeth A. 2019111 Proximity Labeling To Map Host-Pathogen Interactions at the Membrane of a Bacterium-Containing Vacuole in Chlamydia trachomatis-Infected Human Cells Infection and Immunity 87110019-956710.1128/iai.00537-19PMC 680332731405957 · doi ↗ · pubmed ↗
- 5Qu Dezhong Zhou Li Wang Wei Wang Zhe Wang Guoxin Chi Weilin Zhang Biliang 2013315-Ethynylcytidine as a new agent for detecting RNA synthesis in live cells by “click” chemistry Analytical Biochemistry 43410003-269712813510.1016/j.ab.2012.11.02323219562 · doi ↗ · pubmed ↗
- 6Scidmore Marci A. 200621 Cultivation and Laboratory Maintenance of Chlamydia trachomatis Current Protocols in Microbiology 0011934-852510.1002/9780471729259.mc 11a 01s 0018770550 · doi ↗ · pubmed ↗
- 7Stephens RS Kalman S Lammel C Fan J Marathe R Aravind L Mitchell W Olinger L Tatusov RL Zhao Q Koonin EV Davis RW 19981023 Genome sequence of an obligate intracellular pathogen of humans: Chlamydia trachomatis.Science 28253890036-807575475910.1126/science.282.5389.7549784136 · doi ↗ · pubmed ↗
- 8Taylor Hugh R Burton Matthew J Haddad Danny West Sheila Wright Heathcote 2014121 Trachoma The Lancet 38499600140-67362142215210.1016/s 0140-6736(13)62182-025043452 · doi ↗ · pubmed ↗