Toxoplasma gondii C2 Domain Protein Deletion Mutant as a Promising Vaccine Against Toxoplasmosis in Mice
Yifan Luo, Mingfeng He, Shengqiang Yang, Jiahui Qian, Zhengming He, Jiayin Xu, Liyu Guo, Siyu Xiao, Rui Fang

TL;DR
A new vaccine candidate for toxoplasmosis was developed using a modified parasite that is less harmful and triggers strong immune responses in mice.
Contribution
A C2 domain protein deletion mutant of Toxoplasma gondii is shown to be a promising live attenuated vaccine candidate.
Findings
Deleting the TGME49_203240 gene reduces T. gondii virulence and tissue cyst formation in mice.
ME49Δ203240 vaccination protects mice against lethal T. gondii strains and boosts immune responses.
The mutant elicits high IgG antibodies and key cytokines like IFN-γ, TNF-α, and IL-12.
Abstract
Toxoplasma gondii (T. gondii), a parasitic protozoan capable of infecting nearly all warm‐blooded animals, causes significant economic losses in livestock and poses a significant threat to both animal and public health. Despite its impact, no ideal vaccine is currently available to prevent toxoplasmosis. Vesicular transport plays a crucial role in the life cycle of T. gondii, and proteins involved in this process – such as those containing C2 domains – may serve as novel targets for the development of live attenuated vaccines. In this study, we evaluated the feasibility of a C2 domain‐containing protein (TGME49_203240) as a live attenuated vaccine candidate. Our findings suggest that TGME49_203240 may be involved in vesicular transport and that it is essential for T. gondii growth. Deletion of TGME49_203240 reduced parasite virulence and impaired tissue cyst formation in mice. Moreover,…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
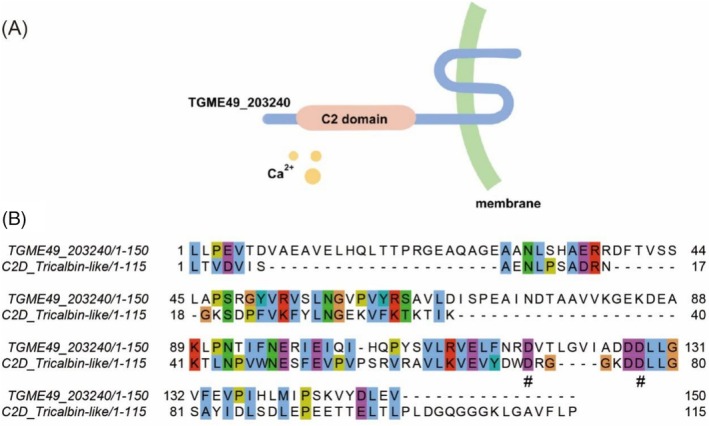 FIGURE 1
FIGURE 1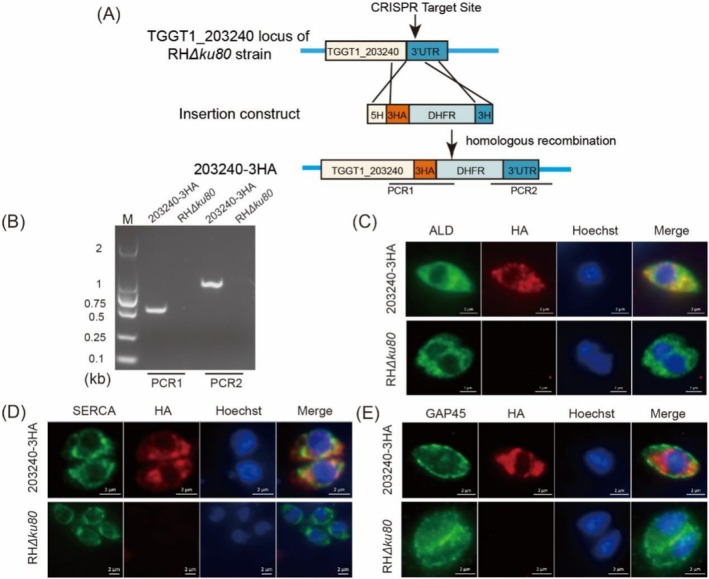 FIGURE 2
FIGURE 2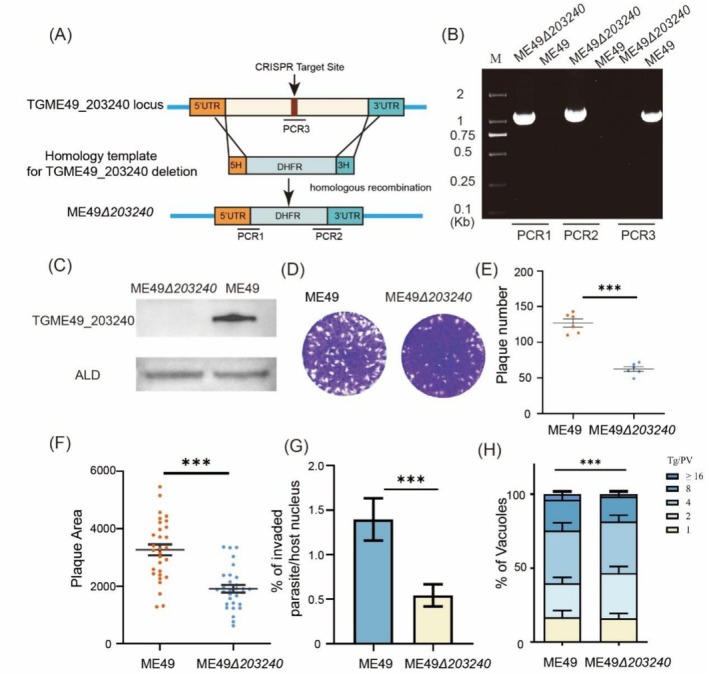 FIGURE 3
FIGURE 3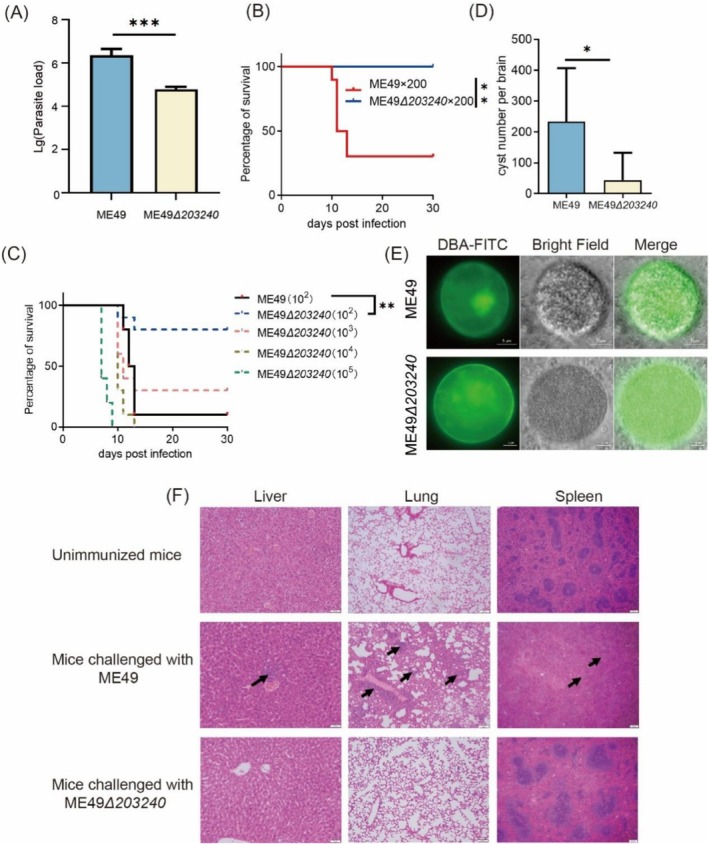 FIGURE 4
FIGURE 4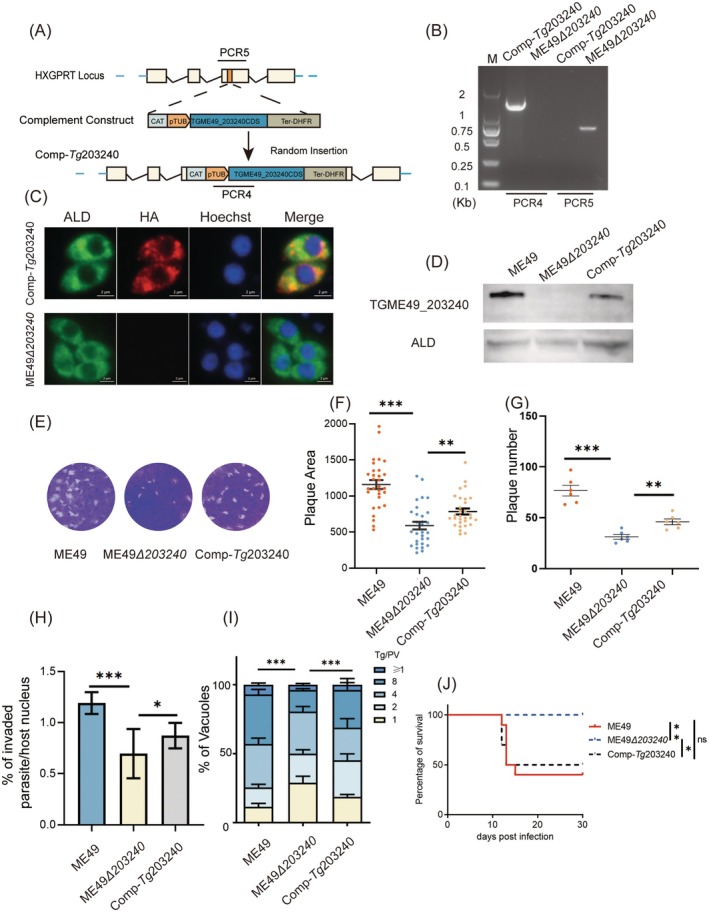 FIGURE 5
FIGURE 5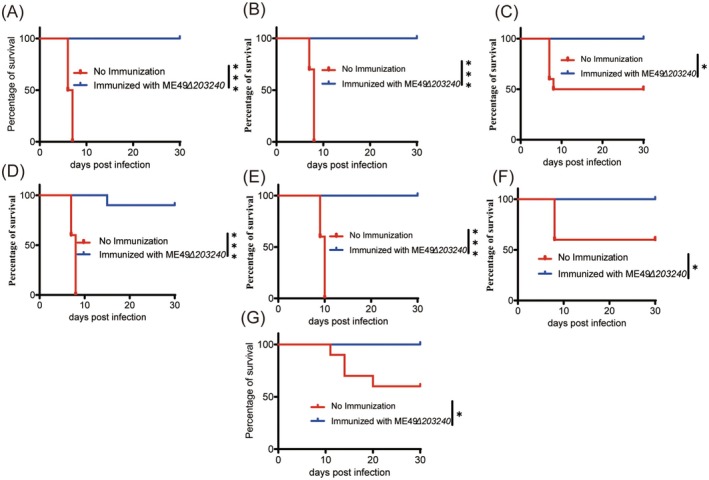 FIGURE 6
FIGURE 6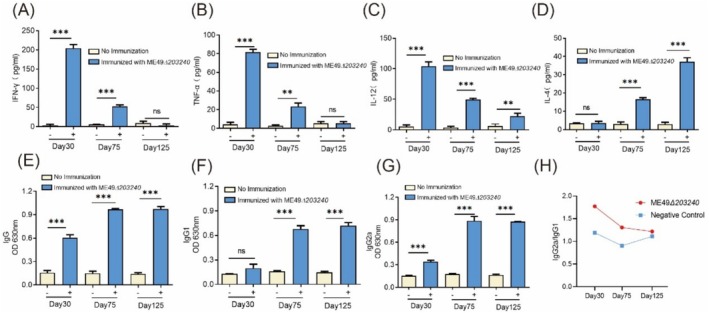 FIGURE 7
FIGURE 7| Primer | Sequence 5′‐3′ | Use |
|---|---|---|
| gRNA‐R | AACTTGACATCCCCATTTAC | To construct gene‐specific CRISPR plasmids |
| gRNA‐203240‐3HA‐F | GCATGACAGGGAACGCCATGGTTTTAGAGCTAGAAATAGC | To construct the 203240‐3HA specific CRISPR plasmid |
| 5H‐3HA‐DHFR‐F | GGCGCACGCGGCAGAACCTCGTAGCTTCACCCTTCATCCCTCGAGCAGCTCGCTACCCCTACGATGTGCCGGATT | Amplification of homologous templates 5H‐3HA‐DHFR‐3H for203240‐3HA strain construction |
| 3H‐3HA‐DHFR‐R | CTTTCCTGACCAACAGTCGCCCCCCTCTCTTCCCGCTCCTCTGTGTCGGCCACCGCTTTCTCAACAGGAAAAAGC | Amplification of homologous templates 5H‐3HA‐DHFR‐3H for203240‐3HA strain construction |
| 203240‐3HA‐PCR1‐F | GCGAGTGTGACTATGTGGCG | PCR1 of 203240‐3HA |
| 203240‐3HA‐PCR1‐R | CGTACTATGGCCGGTCGATG | PCR1 of 203240‐3HA |
| 203240‐3HA‐PCR2‐F | CCGCGAACGATTGGCGATACGTC | PCR2 of 203240‐3HA |
| 203240‐3HA‐PCR2‐R | GCTCGCACTCACCACACAGC | PCR2 of 203240‐3HA |
| gRNA‐ME49 | AGACGCCATACCCCGCACAGGTTTTAGAGCTAGAAATAGC | To construct the ME49 |
| ME49 | GTTGTAAAACGACGGCCAGTAATGTTTCTGCGTCCCCTGT | Amplification of 5′‐homology of TGME49_203240 for p203240:: DHFR homologous recombinant plasmid construction |
| ME49 | GATGTCTTCTGCGCGGGTTGTTTTTAGACAAGAAAGGCCT | Amplification of 5′‐homology of TGME49_203240 for p203240:: DHFR homologous recombinant plasmid construction |
| ME49 | GCCACAAGTTCAGCGTGTCCGGCTCCTACGGGGTTTCCTC | Amplification of 3′‐homology of TGME49_203240 for p203240:: DHFR homologous recombinant plasmid construction |
| ME49 | GCTATGACCATGATTACGCCACCGCTCGCACTCACCATAC | Amplification of 3′‐homology of TGME49_203240 for p203240:: DHFR homologous recombinant plasmid construction |
| PUC19‐F | GGCGTAATCATGGTCATAGCTGT | Amplification of pUC19 vector for p203240:: DHFR homologous recombinant plasmid construction |
| PUC19‐R | ACTGGCCGTCGTTTTACAACG | Amplification of pUC19 vector for p203240:: DHFR homologous recombinant plasmid construction |
| DHFR‐F | CAACCCGCGCAGAAGACATC | Amplification of DHFR for p203240:: DHFR homologous recombinant plasmid construction |
| DHFR‐R | GGACACGCTGAACTTGTGGC | Amplification of DHFR for p203240:: DHFR homologous recombinant plasmid construction |
| ME49 | GTTCTCTCGCGAGGCACTTT | PCR1 of ME49 |
| ME49 | GATTTGTGAGGACGACTCAC | PCR1 of ME49 |
| ME49 | CACGACAGCAGACAACTTTC | PCR2 of ME49 |
| ME49 | CACTTGCGGTTCTTGTGACC | PCR2 of ME49 |
| ME49 | GTCGGTGAACTCTCTTGTGC | PCR3 of ME49 |
| ME49 | TCAGGGGAGGAAGATAAGCC | PCR3 of ME49 |
| gRNA‐Comp‐F | CCTCGCCGAAGTAAGTTCTGGTTTTAGAGCTAGAAATAGC | To construct gene specific CRISPR plasmids |
| Comp‐ | CGGTTGGTGATCCTGGTTGGAC | PCR4 of Comp‐ |
| Comp‐ | CTGAGACCTGCGGTGCAGAAG | PCR4 of Comp‐ |
| Comp‐ | GCATGTTCGGTTGTACATCG | PCR5 of Comp‐ |
| Comp‐ | GCGAATACACATAGTCACACCG | PCR5 of Comp‐ |
| TGME49_203240‐CDS‐F | CTTACGATGTTCCAGATTATGCCGCACCTGCTGCTCCGAACAAG | Amplification of TGME49_203240‐CDS::HA for pTub::TGME49_203240::HA::CAT construction |
| TGME49_203240‐CDS‐R | GCAGCTTCTGTGGGCTGCATAGCTGCTCGAGGGATGAAGGGTGAAG | Amplification of TGME49_203240‐CDS::HA for pTub:: TGME49_203240::HA::CAT construction |
| Comp‐TGME49_203240‐F | CATGATTACGCCAAGCTCGG | Amplification of TGME49_203240‐CDS::HA for pTub:: TGME49_203240::HA::CAT construction |
| Comp‐TGME49_203240‐R | AAACGACGGCCAGTGAATTG | Amplification of TGME49_203240‐CDS::HA for pTub:: TGME49_203240::HA::CAT construction |
| pET32a‐F | GTCGACAAGCTTGCGGCCGCACTCG | Amplification of pET32a vector for all the |
| pET32a‐R | ATGATGATGATGATGGTGCATATGGCC | Amplification of pET32a vector for all the |
| TGME49_203240‐CDS‐F | CTTACGATGTTCCAGATTATGCCGCACCTGCTGCTCCGAACAAG | Amplification of TGME49_203240‐CDS for pET32a‐TGME49_203240 construction |
| TGME49_203240‐CDS‐R | GCAGCTTCTGTGGGCTGCATAGCTGCTCGAGGGATGAAGGGTGAAG | Amplification of TGME49_203240‐CDS for pET32a‐TGME49_203240 construction |
| pET32a‐TGME49_203240‐F | ATGCACCATCATCATCATCATGCACCTGCTGCTCCGAACAA | Construction of pET32a‐TGME49_203240 plasmid for |
| pET32a‐TGME49_203240‐R | GCGGCCGCAAGCTTGTCGACAGAGTAGCCCTGGGGCAGGA | Construction of pET32a‐TGME49_203240 plasmid for |
| RT‐tubulin‐F | CACTGGTACACGGGTGAAGGT | For qPCR detection in parasite burden assay |
| RT‐tubulin‐R | ATTCTCCCTCTTCCTCTGCG | For qPCR detection in parasite burden assay |
- —National Key Research and Development Program of China 10.13039/501100012166
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsToxoplasma gondii Research Studies · Herpesvirus Infections and Treatments · Rabies epidemiology and control
Introduction
1
Toxoplasma gondii is a highly specialised intracellular parasite that causes zoonotic infections in nearly all warm‐blooded animals and has a widespread global prevalence (Rostami et al. 2020). Its life cycle is notably complex, with sexual reproduction occurring exclusively in felines, the definitive hosts, while asexual replication takes place in intermediate hosts. Toxoplasmosis can be a life‐threatening disease, particularly for immunocompromised individuals, such as those with AIDS or cancers. If primary infection occurs during pregnancy, it can lead to miscarriage, stillbirth, congenital toxoplasmosis, fetal deformities and other severe complications (Nayeri et al. 2020; Almeria and Dubey 2021). Beyond its impact on human health, toxoplasmosis also causes substantial economic losses in livestock industries, particularly in small ruminants and swine production, leading to abortions, reduced animal product quality and significant zoonotic risks (Foroutan et al. 2019; Nayeri et al. 2021; Su et al. 2022). The development of a T. gondii vaccine remains a global challenge. Currently, only one commercial vaccine (Toxovax) is produced by attenuating T. gondii through multiple passages after isolating it from lambs. However, Toxovax is only approved for use in sheep, with a high risk of reversion to its virulence (Buxton and Innes 1995). Therefore, there is an urgent need to develop safe and effective vaccines as well as therapeutic strategies for the prevention, control and treatment of Toxoplasma infections.
Currently, multiple T. gondii vaccine candidates have been developed. However, inactivated vaccines have proven ineffective in eliciting robust immunoprotection in animals (Stanley et al. 2004; Saavedra et al. 2004). While subunit vaccines and DNA vaccines have demonstrated only partial immunoprotection, live attenuated vaccines have exhibited the strongest immunoprotective efficacy. As a result, live attenuated vaccines are considered the most promising strategy for T. gondii vaccine development (Bhopale 2003; Dunay et al. 2018; Onile et al. 2020; Chu and Quan 2021). With the advancement in gene editing technologies, the development of genetically modified live attenuated vaccines has become a major research focus. Techniques such as CRISPR/Cas9 and Cre‐loxp have enabled the precise deletion of virulence‐related genes, paving the way for safer and more effective vaccine strains. Several gene‐deletion‐based live vaccine strains, including ΔADSL, Δα‐AMY and ΔOMPDCΔUP, have been developed. However, their safety and long‐term efficacy still require further validation (Fox and Bzik 2010, 2015; Wang et al. 2020; Lyu et al. 2021).
In the field of live attenuated vaccines, most vaccines have been designed by targeting genes associated with parasite adhesion, invasion, replication and biosynthetic pathways of parasites (Fereig and Nishikawa 2016; Chu and Quan 2021). For example, microneme protein (MIC) is a class of proteins released by T. gondii during cell invasion. Low‐virulence RH strains with MIC1 and MIC3 genes knockout have been shown to protect mice against T. gondii cysts challenge (Cerede et al. 2005). Another promising approach involves disrupting the de novo uracil synthesis pathway. Deletion of two enzymes critical – the uridine phosphorylase (UP) and orotidine‐5′‐monophosphate decarboxylase (OMPDC) – renders T. gondii incapable of growth in standard environments, requiring high concentrations of uracil to persist. Mice immunised with either the UP or OMPDC deficient strains exhibited resistance to the challenge of T. gondii type I strain (Fox and Bzik 2010, 2015; Wilson et al. 2012). Similarly, Adenylosuccinate lyase (ADSL), an enzyme involved in the adenylate synthesis pathway, has been identified as a potential target for attenuation. Mice immunised with ADSL‐deficient T. gondii strains developed high levels of specific antibodies, leading to a significant reduction in tissue cyst formation (Wang et al. 2020).
Vesicular transport plays a crucial role in various biological processes and disrupting this pathway can significantly impact the growth and virulence of T. gondii. Therefore, proteins involved in vesicular transport may serve as potential antiparasitic targets. Proteins containing the C2 domain were speculated to be involved in the process of vesicular transport, as the C2 domain can bind a variety of substances, including phospholipids, inositol polyphosphates and other substances involved in membrane translocation and signal transduction (Cho and Stahelin 2006; Zhang and Aravind 2010; Larsen and Sansom 2021; Jahn et al. 2024). Given their functional significance, the C2 protein family may serve as a potential target for the development of live attenuated T. gondii vaccines.
In this study, we identified TGME49_203240, a C2 domain‐containing protein that partially co‐localised with the endoplasmic reticulum. Deletion of the TGME49_203240 gene significantly impacted the growth and virulence of T. gondii. We further evaluated the potential of the TGME49_203240 gene knockout strain (ME49Δ203240) as a live attenuated vaccine and found that ME49Δ203240 provided potent immunoprotection against tachyzoite challenge from T. gondii types I, II and III, as well as cyst challenge from the type II strain in mice. Moreover, ME49Δ203240 induced a robust humoral immune response and a Th1‐biased cellular immune response. These findings highlight TGME49_203240 as a promising target for developing a live attenuated T. gondii vaccine, offering a novel approach to toxoplasmosis prevention.
Experimental Procedures
2
Parasite Strains and Cell Lines
2.1
RHΔku80, RHΔHXGPRT, ME49 and VEG were kept in our laboratory. 203240‐3HA (TGGT1_203240 localised strain), ME49Δ203240 (TGME49_203240 gene knockout strain) and Comp‐Tg203240 (ME49Δ203240 complementation strain) strains were constructed in this study and propagated in human foreskin fibroblasts (HFF).
Human foreskin fibroblasts (HFF) were cultured in a DMEM medium containing 10% fetal bovine serum (FBS, HY, China) and 1% penicillin–streptomycin (Beyotime, China) at a ratio of 3:1 for cell passage.
T. gondii tachyzoites were propagated in HFF cells in a DMEM medium containing 2% FBS and 1% penicillin–streptomycin.
All cells and parasites were cultured at 37°C, 5% CO_2_.
Bioinformatics Analysis
2.2
The sequence of TGME49_203240 was obtained from the ToxoDB database and subjected to analysis in the InterPro database. The conserved sequence of the protein structural domain of the C2D_Tricalbin‐like protein family was downloaded from the NCBI database. The protein sequences were imported into DNASTAR 11 Protean for analysis of antigenic epitopes and hydrophobicity.
Construction of TGME49_203240 Gene Knockout Strain and Complementation Strain
2.3
The corresponding gRNAs were designed using the ToxoDB database, and the CRISPR plasmids were constructed by replacing the UPRT target RNA (gRNA) in pSAG1‐Cas9‐sgUPRT with the corresponding gRNA by Q5 targeted mutagenesis kit (NEB, USA). Homology templates used for gene replacements (pTGME49_203240::DHFR) were constructed by multi‐fragment ligation using the ClonExpress II one‐step cloning kit (Vazyme, Nanjing, China). Briefly, the 5′ and 3′ homology arms of the gene were amplified from the genomic DNA of the ME49 strain and cloned into the pUC19 vector along with DHFR. The CRISPR plasmid targeting TGME49_203240 and the homologous templates, which displace TGME49_203240, were cotransfected into fresh ME49 tachyzoites (Shen and Sibley 2014; Yang et al. 2017). The correct positive clone (ME49Δ203240) was screened by the pyrimethamine and identified by PCR (Shen and Sibley 2014; Yang et al. 2017). To construct the complementation strain, we replaced TgLDH 1 in pCom‐LDH 1 (Xia et al. 2018) with the CDS sequence of TGME49_203240. Then pTub::TGME49_203240‐CAT fragment was transfected into ME49Δ203240, and the correct complementation strain (Comp‐Tg203240) was identified through the use of a drug (chloramphenicol), a limited dilution method, PCR and IFA. All primer sequences are shown in Table 1. The Western Blot and IFA were performed as described previously (Sugi et al. 2017).
Localisation Assay of TGME49_203240
2.4
The 203240‐3HA strain was constructed by transfecting the targeting CRISPR plasmid and a homologous template with a 3HA‐DHFR tag into fresh RHΔku80 tachyzoites by CRISPR/Cas9 gene editing. It was also characterised by screening with 1 μM pyrimethamine, PCR and IFA.
Fresh tachyzoites of the 203240‐3HA strain were transferred to glass slides full of HFF cells for overnight growth and then fixed with 4% paraformaldehyde at room temperature, permeabilised with 0.1% Triton X‐100, closed in 10% FBS (diluted with PBS) at 37°C and stained with mouse‐anti‐HA, mouse‐anti‐GRASP, mouse‐anti‐GAP50, mouse‐anti‐MIC‐2, rabbit‐anti‐TgALD, rabbit‐anti‐TgSERCA, rabbit‐anti‐GRA7, rabbit‐anti‐ROP17 and rabbit‐anti TgGAP45 as primary antibodies. Alexa‐488‐R, Alexa‐596‐M and Hoechst were used as secondary antibodies for staining, and after adding an anti‐fluorescence quencher, the staining was observed under a fluorescence microscope.
Preparation of Polyclonal Antibodies to TGME49_203240
2.5
The CDS sequence of TGME49_203240 was amplified from the cDNA of the ME49 strain. Following this, it was ligated with the pET32a vector to construct the pET32a‐203240 prokaryotic expression plasmid. Protein expression was then carried out with E. coli BL21(DE3) engineering bacteria. Once the bacterial culture had reached the logarithmic growth phase (as indicated by an OD600 reading of 0.5–0.6), the protein expression process was initiated by the addition of IPTG (1 mM) and continued for 4 h at 37°C. Following the disruption of the bacterial fluid, inclusion body proteins were isolated, denatured and purified using a nickel column. Finally, the purified proteins were identified through SDS‐PAGE analysis.
The purified proteins were combined with the Fourier adjuvant in a 1:1 ratio and subsequently administered to mice via subcutaneous injection at separate points. Each mouse received 75 μg of protein. The initial immunisation was conducted with complete F‐adjuvant, while all subsequent immunisations were performed with incomplete F‐adjuvant. Following the fourth immunisation, serum was collected. The binding capacity of the antigen and polyclonal antibody was evaluated through Western blot analysis. (All the results are in the Supplementary chart Figure S1).
Plaque Assay
2.6
The fresh tachyzoites of T. gondii were inoculated into 6‐well plates full of HFF cells (200 Tg/per well) and cultured for 10 days at 37°C in a 5% CO_2_ incubator. After the medium was washed away with PBS, the infected HFF cells were fixed with 4% paraformaldehyde at room temperature, stained with 0.1% crystalline violet at 37°C, dried and the pictures were scanned with a scanner.
Invasive Capability Assay
2.7
Purified tachyzoites were inoculated onto glass slides pre‐seeded with HFF cells (10^6 Tg/well) and incubated at 37°C for 15 min. After washing three times with PBS, the cells were fixed with 4% paraformaldehyde at room temperature for 7 min. Subsequently, the slides were incubated with mouse anti‐T. gondii IgG antibody (1:100 dilution) at 37°C for 30 min, followed by permeabilisation with 0.1% Triton X‐100 for 20 min at 37°C and blocking nonspecific binding with 10% FBS (diluted in PBS) at 37°C for 20 min. Next, the slides were incubated with rabbit anti‐TgALD antibody at 37°C for 30 min, followed by staining with Alexa‐488‐R, Alexa‐596‐M and Hoechst for 20 min. The number of invasive and non‐invasive T. gondii tachyzoites and host cell nuclei were quantified under a fluorescence microscope. A total of 15–20 fields of view were analysed, with at least 200 tachyzoites counted per experiment. The invasion rate was calculated as follows: (number of invaded T. gondii—number of uninvaded T. gondii) / total number of nuclei.
Replication Assay
2.8
Freshly purified T. gondii tachyzoites were inoculated into the glass slides full of HFF cells at 37°C for 1 h. Uninvaded T. gondii tachyzoites were washed away with PBS, and incubation was continued at 37°C for 24 h. Then, the cells were treated as above. The number of parasites per parasitophorous vacuole (PV) was calculated, and at least 200 PVs were counted in each group.
Parasite Burden Assay
2.9
Seven‐week‐old female ICR mice were injected intraperitoneally with 1 × 10^4^ ME49Δ203240 or wild‐type ME49 tachyzoites, respectively, in 5 mice per group. After 5 days of infection, the mice were euthanised, and the peritoneal fluids were collected and centrifuged. Then, the gDNA was extracted by taking 1/20 of the mixed ascites for the detection of the parasite‐loaded volume using qPCR.
Detection of T. gondii Virulence and Tissue Cyst‐Forming Ability
2.10
Purified fresh T. gondii tachyzoites were counted and diluted with FBS‐free DMEM medium. For the virulence experiments, each mouse was injected with 200 ME49Δ203240 tachyzoites, and the control was injected with the same number of wild‐type ME49 strains. Brain tissues of mice were collected after infected parasites for 30 days, and the number of tissue cysts was detected by DBA‐FITC.
For the gradient virulence experiments, each mouse was injected with 10^2^, 10^3^, 10^4^ and 10^5^ ME49Δ203240 strains, and the control group was injected with 200 tachyzoites of wild‐type ME49 strains.
HE Staining of the Liver, Spleen and Lung
2.11
After mice were immunised with 200 ME49Δ203240 and ME49 tachyzoites for 30 days, the liver, lungs, spleen and liver were taken and fixed in 4% paraformaldehyde at room temperature for 24 h (3 mice per group), dehydrated with ethanol, cleared with xylene, embedded in wax and sliced with a slicer. The unimmunised mice were used as controls. Sections were then stained with haematoxylin and eosin (H&E).
Immunoprotection Against Acute T. gondii and Chronic Infection
2.12
Female ICR mice were immunised with 200 tachyzoites of ME49 Δ203240 strains through intraperitoneal injection at 30‐ or 75‐day post‐immunisation. Then, the immunised mice were challenged with 10^4^ tachyzoites of RHΔHXGPRT, ME49, or VEG, by intraperitoneal injection (10 mice for each group), or 50 fresh cysts of ME49 by oral administration. The survival curves of the mice were observed, recorded and statistically analysed for 30 days.
Detection of Serum T. gondii‐Specific Antibodies and Cytokines by ELISA
2.13
Sera from mice immunised with the 200 ME49Δ203240 strain for 30, 75 and 125 days were collected. The TSA of T. gondii is prepared as in this description (Yang et al. 2020). The 96‐well plates were coated overnight with TSA at a concentration of 16 μg/mL. After blocking, mouse serum (diluted 1:100) was added and incubated at 37°C for 45 min. Subsequently, the plates were incubated with goat anti‐mouse IgG‐HRP, goat anti‐mouse IgG2a and goat anti‐mouse IgG1 for 30 min. Colour development was initiated by adding TMB substrate for 5 min, followed by the addition of a stop solution. The OD630 was measured using a microplate reader.
Cytokines were detected by the Elisa Max Deluxe Set Mouse kit (Biolegend, USA) for IL‐4, IL‐12, IFN‐γ and TNF‐α.
Statistical Analysis
2.14
Statistical analyses were performed in Prism 10 software (GraphPad Software Inc., La Jolla, CA, USA) with unpaired t‐test, two‐way ANOVA test, Mantel‐Cox log‐rank test, p > 0.05 for ns (not significant differences), p < 0.05 for * (statistical difference), p < 0.01 for ** (statistically significant difference) and p < 0.001 for *** (extremely statistically significant difference).
Results
3
Bioinformatics Analysis
3.1
Using the ToxoDB database and InterPro database, we identified TGME49_203240, a protein containing C2 domains which may be involved in vesicle transport in T. gondii. It has a C2 domain at the N‐terminal (amino acids 100–249) and three transmembrane structural domains at the C‐terminal end (Figure 1A). However, not all C2 domains are functionally active; some serve merely as structural scaffolds or remain nonfunctional (Cho and Stahelin 2006; Zhang and Aravind 2010; Corbalan‐Garcia and Gomez‐Fernandez 2014; Larsen and Sansom 2021). To further investigate TGME49_203240, we conducted a sequence comparison of its C2 domain using the NCBI database. Our results revealed that the sequence of the TGME49_203240 C2 domain protein exhibits a high degree of similarity to the sequence of the C2 domain protein family of C2D_Tricalbin‐like proteins which are involved in membrane transport and sorting, suggesting that the C2 domains of TGME49_203240 may be functionally active (Figure 1B) (Essen et al. 1996; Xu et al. 1998; Verdaguer et al. 1999). Based on these findings, we hypothesise that TGME49_203240 may be involved in the process of vesicle transport in T. gondii.
Bioinformatics analysis of TGME49_203240. (A) Schematic structure of TGME49_203240. Data from InterPro. (B) Comparison of the structural domain of the C2 domain of TGME49_203240 with conserved sequences of the C2D_Tricalbin‐like family. Putative Ca2+ ion‐binding sites are labelled with the ‘#’ under the sequence.
TGME49_203240 Is Partially Co‐Localised With T. gondii Endoplasmic Reticulum
3.2
To further explore whether TGME49_203240 is involved in the vesicular transport process of T. gondii, its subcellular localisation needs to be determined. Thus, the 203240‐3HA strain was constructed, and the PCR identification confirmed its successful generation (Figure 2A,B). Subsequently, the subcellular localisation of TGME49_203240 was determined by indirect immunofluorescence (IFA) (Figure 2C–E). The results showed that the HA tag was partially co‐localised with cytoplasmic protein (TgALD) and endoplasmic reticulum (TgSERCA). However, it does not colocalise with gliding‐associated protein (TgGAP45). These findings suggest that TGME49_203240 primarily functions in the endoplasmic reticulum and cytoplasm of T. gondii. Additionally, a murine polyclonal antibody against TGME49_203240 was generated to examine potential differences in localisation across different T. gondii strains. The results indicated that TGME49_203240 localisation remains consistent between RH and ME49 strains (Figure S2).
Localisation assay of TGME49_203240. (A) Schematic diagram of 203240‐3HA strain construction. The 3HA‐DHFR tag was integrated into the TGGT1_203240 gene of RHΔku80 strain by CRISPR/Cas9 gene editing system. (B) PCR identification of 203240‐3HA strain. PCR1 and PCR2 represent the correct position of the 3HA‐DHFR insertion into the RHΔku80 strain. (C–E) IFA to determine TGME49_203240 subcellular localisation. Scale bar = 2 μm. (C) Immunofluorescence analysis of the transgenic parasites with mouse anti‐HA and rabbit anti‐TgALD antibodies. (D) Immunofluorescence analysis of the transgenic parasites with mouse anti‐HA and rabbit anti‐TgSERCA antibodies. (E) Immunofluorescence analysis of the transgenic parasites with mouse anti‐HA and rabbit anti‐TgGAP45 antibodies.
TGME49_203240 Is Important for T. gondii Growth In Vitro
3.3
To investigate the role of TGME49_203240 in T. gondii, a knockout strain (ME49Δ203240) was generated using CRISPR/Cas9 gene‐editing technology (Figure 3A). Following pyrimethamine selection, the correct mutant strain was identified by PCR analysis (Figure 3B). Western blot analysis further confirmed the successful deletion of TGME49_203240 at the protein level (Figure 3C).
*Deletion of TGME49_203240 and effects on growth of T. gondii in vitro. (A) Schematic diagram of TGME49_203240 gene deletion strain construction. (B) PCR identification of ME49Δ203240 strain with deletion of TGME49_203240. PCR1 and PCR2 represent the successful insertion of the DHFR tag into the correct gene position, and PCR3 represents the successful deletion of the TGME49_203240 gene. (C) Western Blot identification of ME49Δ203240 strain with deletion of TGME49_203240. Mouse‐anti‐TGME49_203240 and rabbit‐anti‐TgALD as primary antibodies. (D) Plaque assay of ME49Δ203240 and ME49 strain. (E) Plaque number of ME49 and ME49Δ203240 strain, ***p < 0.001, unpaired t‐test. (F) Plaque area of ME49 and ME49Δ203240 strain, ***p < 0.001, unpaired t‐test. (G and H) In vitro growth assay of strain ME49 and ME49Δ203240 strain with mouse‐anti‐T. gondii and rabbit‐anti‐TgALD. (G) Invasion assay of ME49 and ME49Δ203240 strain, ***p < 0.001, two‐way ANOVA. (H) Replication assay of ME49Δ203240 and ME49 strain, **p < 0.001, two‐way ANOVA.
To assess the impact of TGME49_203240 gene deletion on the growth of Toxoplasma tachyzoites in vitro, the phenotype assays of ME49Δ203240 were performed. Plaque assay showed that the size and number of plaques formed by strain ME49Δ203240 were significantly different from those formed by wild‐type strain ME49, with plaques formed by strain ME49Δ203240 being smaller than those formed by wild‐type strain ME49, and with fewer plaques formed by strain ME49Δ203240 (Figure 3D–F). Additionally, the results of the invasion capability assay showed that the deletion of the TGME49_203240 gene significantly reduced the invasion ability of the ME49Δ203240 strain compared with the ME49 strain (Figure 3G). Besides, the replication assay demonstrated that the ME49Δ203240 strain exhibited significantly slower replication in vitro compared to the wild‐type ME49 strain. Collectively, these results demonstrated that TGME49_203240 gene deletion significantly impairs the in vitro growth of Toxoplasma tachyzoites (Figure 3H).
Furthermore, we aimed to investigate whether the deletion of TGME49_203240 affects the localisation of several organelle markers involved in T. gondii vesicle transport. Notably, the deletion of TGME49_203240 did not significantly alter the localisation of cytoplasmic, Golgi, rhoptry, microneme, dense granule, or gliding‐associated proteins (Figure S3).
TGME49_203240 Gene Deletion Affects the Proliferation Rate of T. gondii in Mice
3.4
To evaluate whether the deletion of the TGME49_203240 gene impacts the replication and proliferation rate of tachyzoites in vivo, fluorescence qPCR was used to quantify the relative content of tachyzoites in the peritoneal fluid of mice. The results showed that the proliferation rate of the ME49Δ203240 strain was reduced significantly in mice compared with the wild type (Figure 4A). These findings suggest that TGME49_203240 gene deletion leads to a significant reduction in the replication and proliferation rate of tachyzoites in vivo.
*TGME49_203240 deletion results in reduced parasite virulence and reduced tissue cyst formation. (A) Parasite load experiment of ME49 and ME49Δ203240 in mouse. 7‐week‐old female ICR mice were injected with 104 ME49 or ME49Δ203240 strain, and parasite burdens in the peritoneal fluid of mice were detected after infecting 5 days by qPCR, ***p < 0.001, unpaired t‐test. (B) Virulence test, 7‐week‐old ICR mice, 10 in each group, were infected with 200 ME49 or ME49Δ203240 strain, mantel Cox log‐rank test analysis, **p < 0.01. (C) Gradient virulence test, 7‐week‐old ICR mice, 10 in each group, were infected with 102 ME49 and 102, 103, 104, 105 ME49Δ203240 strain, mantel Cox log‐rank test analysis, **p < 0.01. (D) Brain cyst loads. The surviving mice in (B) were sacrificed, and the number of brain cysts were stained with DBA‐FITC and counted under a fluorescence microscope, p < 0.05, unpaired t‐test. (E) DBA staining of brain cysts. Tissue cysts stained with DBA‐FITC and imaged on fluorescence microscopy. Scale bar = 5 μm. (F) HE stained liver, lung and spleen sections from mice injected with ME49Δ203240 and ME49 strains after 30 days. Arrows represent pathological changes. In mice infected with the ME49 strain, the arrows in the histopathological sections of the liver represent foci of mixed cell infiltration (macrophages and neutrophils), the arrows in the lungs represent a large perivascular mixed cell infiltration (macrophages and neutrophils), and the arrows in the spleen represent a severe reduction in the number of lymphocytes within the splenic white and red marrow.
TGME49_203240 Deletion Results in Reduced Parasite Virulence and Reduced Tissue Cyst Formation
3.5
Virulence experiments and gradient virulence experiments were conducted to investigate whether the deletion of the TGME49_203240 gene affects the virulence of T. gondii. In the virulence experiment, the 30‐day survival curves of mice are shown in Figure 4B, and the survival rate of mice injected with the ME49Δ203240 strain was 100%, whereas the mortality rate of mice injected with the wild‐type ME49 strain was 70%. The result suggests that the deletion of the TGME49_203240 gene reduces the virulence of parasites. Subsequently, to further confirm the extent of reduced virulence of Toxoplasma after deletion of the TGME49_203240 gene, the gradient virulence assay was performed, and mice were injected intraperitoneally with 10^2^, 10^3^, 10^4^ and 10^5^ ME49Δ203240 strains, while the controls were injected with 200 ME49 strains. As shown in Figure 4C, inoculation with 10^4^ and 10^5^ ME49Δ 203240 strains resulted in the death of all mice within 30 days, with a survival rate of 30% in the 10^3^ group, 80% in the 10^2^ group and 10% in the mice inoculated with wild‐type ME49 strains, suggesting that the deletion of the TGME49_203240 gene reduced the virulence of T. gondii to a certain degree, but the ME49Δ203240 strain remained highly virulent at high doses.
In addition, the ability to form brain cysts of the ME49Δ203240 strain was evaluated. After being injected with 200 tachyzoites of the ME49Δ203240 strain for 30 days, live mice were euthanised, and their brain tissues were collected for counting. The results are shown in Figure 4D,E. Compared with mice injected with the ME49 strain, those mice injected with the ME49Δ203240 strain had a significantly lower number of brain cysts (p < 0.05). This indicates that deletion of the TGME49_203240 gene significantly reduces the ability of T. gondii to form brain cysts.
To further evaluate the safety of the ME49Δ203240 strain, lung, spleen and liver tissues were collected from mice injected with 200 ME49Δ203240 and ME49 tachyzoites after 30 days and examined for tissue damage by HE staining. Histopathological analysis revealed almost no significant changes in the tissue sections of mice immunised with ME49Δ203240 strain compared to the uninfected mice (Figure 4F). In contrast, mice infected with wild‐type ME49 exhibited marked inflammation in the liver and lungs, characterised by macrophage and neutrophil infiltration. Lung tissue displayed alveolar wall thickening and structural damage, while the spleen showed a severe reduction in lymphocytes in both the white and red pulp.
The Complement of TGME49_203240 Restores T. gondii In Vitro Growth Defects and In Vivo Virulence
3.6
To verify that the reduced virulence and growth defects observed in ME49Δ203240 strain were indeed due to the deletion of the TGME49_203240 gene, a complement strain (Comp‐Tg203240) was constructed by inserting the CDS fragment of TGME49_203240 into the HXGPRT locus of the ME49Δ203240 strain (Figure 5A). Following the selection with chloramphenicol, PCR analysis confirmed the correct insertion of the TGME49_203240 CDS fragment (Figure 5B). IFA further verified the successful construction of the Comp‐Tg203240 strain by detecting the expression of the HA‐tagged protein (Figure 5C). Additionally, Western blot analysis demonstrated the presence of TGME49_203240 in the Comp‐Tg203240 strain, while no expression was detected in ME49Δ203240, confirming the successful restoration of TGME49_203240 expression (Figure 5D).
*The complement of TGME49_203240 restores T. gondii in vitro growth defects and in vivo virulence. (A) Schematic diagram of Comp‐Tg203240 strain construction. (B) PCR identification of Comp‐Tg203240 strain. PCR4 represents the successful insertion of the TGME49_203240 CDS sequence, and PCR5 represents the disruption of the HXGPRT site. (C) IFA identification of Comp‐Tg203240 complementation strain. Scale bar = 2 μm. (D) Western Blot identification of Comp‐Tg203240 strain with complement of TGME49_203240. Primary antibodies were selected from mouse‐anti‐TGME49_203240 and rabbit‐anti‐TgALD. (E–I) ME49, ME49Δ203240 and Comp‐Tg203240 phenotyping experiments. (E) Plaque assay, 200 parasites infected HFF cells, grown for 10 days and stained with crystal violet. (F) Plaque area, unpaired t‐test, ***p < 0.001, **p < 0.01. (G) Plaque number, unpaired t‐test, ***p < 0.001, **p < 0.01. (H) Invasive capability assay, ***p < 0.001, *p < 0.05, two‐way ANOVA. (I) Replication assay, ***p < 0.001, two‐way ANOVA. (J) Virulence test, seven‐week‐old ICR mice, 10 in each group, were infected with 200 ME49, ME49Δ203240 and Comp‐Tg203240 strain, **p < 0.01, p < 0.05, ns, not significant, mantel Cox log‐rank test analysis.
Plaque assay results demonstrated that the Comp‐Tg203240 strain formed larger plaques compared to the ME49Δ203240 strain (Figure 5E), with a statistically significant difference in plaque area and numbers between the two strains (Figure 5F,G). Furthermore, the results of the invasive capability and replication assay showed that the phenotypic defects of the ME49Δ203240 strain were somewhat restored after complement with TGME49_203240 CDS (Figure 5H,I). These results further verified that TGME49_203240 is important for the in vitro growth of T. gondii.
The virulence experiments showed that the survival rate of mice infected with the wild‐type ME49 strain was 40%, mice infected with the Comp‐Tg203240 strain had a 50% survival rate while mice infected with the ME49Δ203240 strain had a 100% survival rate (Figure 5J). These results suggest that the complementation of TGME49_203240 significantly enhances virulence, highlighting the critical role of TGME49_203240 in T. gondii virulence.
ME49Δ203240
Vaccination Provides Robust Immune Protection in Mice
3.7
The deletion of the TGME49_203240 gene significantly reduced the virulence of T. gondii and its proliferative ability in mice, which made us speculate that the ME49Δ203240 strain may be a potential live attenuated vaccine against T. gondii. To evaluate whether the ME49Δ203240 strain could provide immunoprotection in mice, immunoprotection experiments were performed.
Following a 30‐day immunisation period with 200 ME49Δ203240 strains, mice were challenged with 1 × 10^4^ T. gondii tachyzoites of type I strains (RHΔHXGPRT), type II strains (ME49) and type III strains (VEG). The results demonstrated that the survival rate of all mice in the immunised group was 100%, whereas the survival rate of mice in the unimmunised group was 0% (type I), 0% (type II) and 50% (type III), respectively (Figure 6A–C). These findings suggest that the ME49Δ203240 strain provides robust immune protection against the challenge of T. gondii I, II and III strains within 30 days. Moreover, to determine whether the ME49Δ203240 strain could provide long‐term immune protection for mice, the duration of immunisation was extended to 75 days in this study. The results demonstrated that the survival rates of mice immunised with 200 ME49Δ203240 strains for 75 days were 90% (type I), 100% (type II) and 100% (type III), whereas the survival rates of mice in the unimmunised group were 0% (type I), 0% (type II) and 60% (type III) (Figure 6D–F).
*ME49Δ203240 immunisation protected mice from T. gondii tachyzoite and cyst challenge. (A‐C) Seven‐week‐old ICR mice were immunised with ME49Δ203240 tachyzoites. After 30 days post‐immunisation, the mice were challenged with 104 tachyzoites of RHΔHXGPRT (A), ME49 (B), or VEG (C) via intraperitoneal injection and their survival was monitored for another 30 days. (D–F) Seven‐week‐old ICR mice were immunised with ME49Δ203240 tachyzoites. After 75 days post‐immunisation, the mice were challenged with 104 tachyzoites of RHΔHXGPRT (D), ME49 (E) or VEG (F) via intraperitoneal injection, and their survival was monitored for another 30 days. Unimmunised mice (n = 10 mice for each strain) were included as a control. (G) Seven‐week‐old ICR mice were immunised with ME49Δ203240 tachyzoites. After 30 days post‐immunisation, the mice were challenged with 50 cysts of the ME49 strain, and the survival of the mice was monitored for 30 days. Unimmunised mice (n = 10 mice for each strain) were included as a control. Mantel‐Cox log‐rank test analysis, *p < 0.05, **p < 0.001.
Besides, to evaluate whether ME49Δ203240 vaccination could provide protective immunity against cyst infection, the mice immunised with ME49Δ203240 were challenged with 50 cysts of ME49. The results showed that the survival rate of mice in the immunised group was 100%, whereas the survival rate of mice in the unimmunised group was 60% (Figure 6G). This indicates that ME49Δ203240 vaccination effectively induced protective immunity against chronic Toxoplasma infection. In summary, these findings further indicated that the ME49Δ203240 strain confers long‐term immunoprotection in mice following immunisation.
ME49Δ203240
Vaccinated Mice Induced Strong Humoral and Cellular Immune Responses
3.8
To investigate the immunoprotective mechanisms induced by the ME49Δ203240 strain against T. gondii challenge, we analysed specific antibody and cytokine responses in the sera of immunised mice.
The results showed that the immunised mice produced high levels of IFN‐γ, TNF‐α and IL‐12 at 30 and 75 days post‐immunisation. However, after 75 and 125 days, IFN‐γ, TNF‐α and IL‐12 significantly declined compared to 30 days, suggesting that the immune response peaked early around day 30 and gradually waned over time. In contrast, IL‐4 was significantly elevated after days 75 and 125, indicating a potential activation of anti‐inflammatory mechanisms and a shift in the immune response over time (Figure 7A–D).
*ME49Δ203240 vaccinated mice induced strong humoral and cellular immune responses. (A–D) ELISA for detection of serum cytokines in mice, which immunised ME49Δ203240 strain 30, 75 and 125 days. (A) IFN‐γ; (B) TFN‐α; (C) IL‐12; (D) IL‐4, ns = not significant, **p < 0.01, ***p < 0.001, unpaired t‐test. (E and F) ELISA for detection of serum‐specific antibodies in mice, which immunised ME49Δ203240 strain 30 days, 75 days and 125 days. (E) IgG; (F) IgG1; (G) IgG2a; (H) IgG2a/IgG1, yellow for unimmunised group, blue for immunised group, ns = not significant, **p < 0.001, unpaired t‐test.
The results also showed that the level of specific IgG antibodies in mice inoculated with the ME49Δ203240 strain gradually increased with time, peaking at about 75 days and maintaining a high level of IgG at 125 days. This suggests that the ME49Δ203240 strain induced a strong humoral immune response in mice (Figure 7E). To further explore the link between the immune responses immunised in mice by the ME49Δ203240 strain, we examined the IgG2a and IgG1 antibodies. Based on the results of the IgG2a/IgG1 ratio, we found that after 30 days of immunisation of mice with the strain, the IgG2a/IgG1 ratio was much greater than 1 and the negative control, which indicated that the ME49Δ203240 strain induced a Th1‐biased immune response in mice; however, the IgG2a/IgG1 ratio gradually decreased at 75 and 125 days, which may reflect the body's regulatory adjustment of the Th1/Th2 immune balance over time (Figure 7F–H).
In conclusion, immunisation with the ME49Δ203240 strain induced a strong humoral immune response along with a Th1‐biased cellular immune response in mice, which in turn protected mice against the challenge of T. gondii.
Discussion
4
T. gondii is widespread globally, posing a significant threat to public health and farming. Nevertheless, there is currently no effective vaccine or drug available for the prevention and treatment of T. gondii infection (Chu and Quan 2021). In this study, we explored novel strategies for developing a T. gondii vaccine by targeting vesicular transport proteins which may play a critical role in the parasite's growth and virulence. Through database and bioinformatic analysis, we found a protein containing a C2 domain (TGME49_203240) and identified that it was localised in the cytoplasm and endoplasmic reticulum. Additionally, we evaluated the potential of the ME49Δ203240 strain as a live attenuated vaccine candidate and investigated the immune protection mechanism it provides in mice. Our findings demonstrated that the ME49Δ203240 strain conferred strong immune protection in mice.
Bioinformatics analysis suggests that TGME49_203240 may be involved in vesicular transport in T. gondii, a process essential for the parasite's life activities and virulence. Numerous proteins were found to be involved in the vesicular transport process of T. gondii, including members of the SNARE protein family, T. gondii SM proteins, endosomal vesicle plugging complex (HOPS), GTPase (DrpB) and Rab (Rab11a, Rab11b) proteins, etc. (Breinich et al. 2009; Agop‐Nersesian et al. 2010; Jackson et al. 2013; Cao et al. 2020; Shu et al. 2020; Venugopal et al. 2020; Fu et al. 2023). Additionally, proteins containing the C2 domain, such as Ferlin family proteins and TgDOC2, have also been implicated in T. gondii vesicle translocation (Tagoe et al. 2021). It is reported that the C2 domain‐containing proteins may function as Ca^2+^‐dependent targeting proteins involved in membrane translocation and signal transduction (Nalefski and Falke 1996; Corbalan‐Garcia and Gomez‐Fernandez 2014).
In this study, we identified TGME49_203240, a C2 domain‐containing protein, through the ToxoDB database. Sequence analysis revealed that TGME49_203240 shares structural similarities with SNARE family proteins involved in vesicle fusion, with one side anchoring to the membrane and the other binding to interaction partners. Further sequence comparison, we found that the C2 domain of TGME49_203240 resembles that of C2D_Tricalbin‐like proteins and has two Ca^2+^ binding sites. We also found that TGME49_203240 is localised in the cytoplasm and endoplasmic reticulum of T. gondii, which further suggests that TGME49_203240 may be involved in vesicular transport. However, TGME49_203240 does not colocalise with gliding‐associated protein (TgGAP45), suggesting that TGME49_203240 may be involved in the transport of substances between the endoplasmic reticulum and other organelles as well as vesicle synthesis of T. gondii but not participating in extracellular vesicle transport. Additionally, deletion of TGME49_203240 had no significant effect on the localisation of cytoplasmic, Golgi, rhoptry, microneme, dense granule, or gliding‐associated proteins, indicating that its function may be distinct from known vesicular transport pathways and additional research may be needed to clarify the function of TGME49_203240 (Figure S3).
Researches support that some C2 domain‐containing proteins are essential for T. gondii growth. The absence of these proteins impacts Toxoplasma biological processes such as adhesion, invasion, proliferation, egress and tissue cyst formation. For example, the deletion of FER1 and FER2 of the Ferlin family of proteins affects protein secretion from T. gondii microsomes and rods (Coleman et al. 2018; Tagoe et al. 2021). Whereas knockdown of TgDOC2, a protein containing a double C2 structural domain, affects the secretion of Toxoplasma microsomal proteins. The deletion of the RASP2 protein, a protein containing a C2 domain and a PH domain, affects the invasive ability of Toxoplasma tachyzoites and reduces rod‐shaped body protein secretion (Suarez et al. 2019). In our study, we found that TGME49_203240 gene deletion causes a defective phenotype characterised by reduced invasive and replicative capacity of T. gondii. Given that T. gondii FER1 and FER2 also have the same invasive ability‐deficient phenotype, we hypothesised that the functional impairment observed in the TGME49_203240 knockout strain may be attributed to its C2 domain, which is to be further investigated in subsequent studies.
The immunoprotective mechanisms against T. gondii are complex, involving both humoral and cellular immunity. As an intracellular parasite, T. gondii cannot be effectively controlled by high titers of specific IgG alone, making the cellular immune response crucial for protection (Chu and Quan 2021; Sana et al. 2022). Previous studies have shown that while many subunit vaccines for T. gondii elicit strong humoral immune responses, they often fail to provide adequate protection against highly virulent strains. For example, Toxoplasma tyrosine hydroxylase (TgTH) provided only partial immunoprotection in mice, while subunit vaccines incorporating GRA2 and GRA5 enhanced the Th1‐type immune response in mice but only prolonged the survival of mice infected with RH strains of the parasite (Ching et al. 2016; Zhang et al. 2020). In contrast, live attenuated vaccines have demonstrated superior protective efficacy. For example, mice immunised with the ME49Δ6gpdh strain for 30 days exhibit a survival rate of 80% and 100% when challenged with T. gondii RHΔku80 and ME49 strains, respectively (Guo et al. 2023). In this study, mice immunised with the ME49Δ203240 strain for 30 days and subsequently challenged with T. gondii tachyzoites of RHΔHXGPRT, ME49 and VEG strains, as well as cysts of the ME49 strain, exhibited a 100% survival. These findings indicate that the ME49Δ203240 strain induced an immune response similar to that generated by natural T. gondii infections and provides robust protection against T. gondii challenge.
Meanwhile, we found that ME49Δ203240 induced high levels of specific antibody IgG and cytokines such as cytokines IFN‐γ and TNF‐α compared to live attenuated vaccines such as Δα‐amy, ΔADSL, etc. (Wang et al. 2020; Yang et al. 2020). This suggests that the mechanism of immune response produced by the ME49Δ203240 strain immunising the organism may be similar to that of these vaccines. To further characterise the humoral immune response, we examined the IgG antibody subtypes, IgG2a and IgG1, which are generally considered indicators of a Th1‐type and Th2‐type immune response, respectively. The IgG2a/IgG1 ratio, a commonly used marker for evaluating vaccine immune response, with a ratio greater than 1, indicating a Th1‐biased response, which is critical for the clearance of intracellular pathogens (Golkar et al. 2007; Xu et al. 2014; Abasi et al. 2019; Rostami et al. 2020). In this study, the IgG2a/IgG1 ratio of mice immunised with the ME49Δ203240 strain at 30 days post‐immunisation was much greater than 1, indicating that the ME49Δ203240 strain induced a Th1‐biased immune response in mice. These findings collectively demonstrated that ME49Δ203240 is a highly promising attenuated T. gondii vaccine candidate. However, further evaluation of its immunoprotective efficacy in other host species is necessary, along with rigorous safety assessments. In addition, ME49Δ203240 remains highly virulent at high doses and, like most live attenuated vaccines, can still form tissue cysts in mice. Therefore, co‐deletion with other genes could be considered to further reduce the virulence.
In conclusion, this study demonstrated that the deletion of TGME49_203240 impairs the ability of the T. gondii strain ME49 to invade, proliferate and form tissue cysts, resulting in a reduction of its virulence. Additionally, the immunoprotective efficacy and safety of the ME49Δ203240 strain were evaluated in depth in this study. Our findings revealed that the ME49Δ203240 strain induces a Th1‐biased cellular immune response as well as a humoral immune response in mice, providing immunoprotection against challenges with both T. gondii tachyzoites and bradyzoites. These results underscore the potential of TGME49_203240 as a promising target for the development of a live attenuated T. gondii vaccine. However, further refinement is necessary to ensure the safety and optimise the immunoprotection capabilities of the ME49Δ203240 strain.
Author Contributions
Yifan Luo: formal analysis, investigation, writing – original draft. Mingfeng He: investigation, methodology. Shengqiang Yang: investigation, methodology, writing – review and editing. Jiahui Qian: investigation, methodology, writing – review and editing. Zhengming He: investigation, methodology. Jiayin Xu: investigation, methodology. Liyu Guo: investigation, methodology. Siyu Xiao: conceptualization, data curation, formal analysis, funding acquisition, project administration, resources, supervision, visualization, writing – original draft, writing – review and editing. Rui Fang: conceptualization, data curation, formal analysis, funding acquisition, project administration, supervision, resources, writing – original draft, writing – review and editing, visualization.
Ethics Statement
All 7‐week‐old female ICR mice were purchased from Hubei Provincial Laboratory Animal Research Centre, Licence No.: SCXK‐2020‐0084. All mice were kept in a standard SPF environment according to the requirements of the Animal Ethics Committee of Huazhong Agricultural University, following the requirements of animal welfare and related ethical requirements (Approval No.: HZAUMO‐2024‐0076).
Conflicts of Interest
The authors declare no conflicts of interest.
Supporting information
Figure S1.–S3.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Abasi, E. , S. Shahabi , M. Golkar , P. Khezri , and H. H. Mohammadzadeh . 2019. “Evaluation of Immunogenic Effect of Toxoplasma gondii Recombinant SAG‐1 Antigen With Propranolol as Adjuvant in BALB/c Mice.” Advances in Pharmaceutical Bulletin 9: 632–639.10.15171/apb.2019.073PMC 691218531857968 · doi ↗ · pubmed ↗
- 2Agop‐Nersesian, C. , S. Egarter , G. Langsley , B. J. Foth , D. J. P. Ferguson , and M. Meissner . 2010. “Biogenesis of the Inner Membrane Complex Is Dependent on Vesicular Transport by the Alveolate Specific GT Pase Rab 11B.” P Lo S Pathogens 6, no. 7: e 1001029.20686666 10.1371/journal.ppat.1001029 PMC 2912401 · doi ↗ · pubmed ↗
- 3Almeria, S. , and J. P. Dubey . 2021. “Foodborne Transmission of Toxoplasma gondii Infection in the Last Decade. An Overview.” Research in Veterinary Science 135: 371–385.33148402 10.1016/j.rvsc.2020.10.019 · doi ↗ · pubmed ↗
- 4Bhopale, G. M. 2003. “Development of a Vaccine for Toxoplasmosis: Current Status.” Microbes and Infection 5: 457–462.12738002 10.1016/s 1286-4579(03)00048-0 · doi ↗ · pubmed ↗
- 5Breinich, M. S. , D. J. Ferguson , B. J. Foth , et al. 2009. “A Dynamin Is Required for the Biogenesis of Secretory Organelles in Toxoplasma gondii .” Current Biology 19: 277–286.19217293 10.1016/j.cub.2009.01.039PMC 3941470 · doi ↗ · pubmed ↗
- 6Buxton, D. , and E. A. Innes . 1995. “A Commercial Vaccine for Ovine Toxoplasmosis.” Parasitology 110, no. Suppl: S 11–S 16.7784124 10.1017/s 003118200000144 x · doi ↗ · pubmed ↗
- 7Cao, S. , H. Chen , X. Liang , et al. 2020. “The Sec 1/Munc 18‐Like Proteins Tg Sec 1 and Tg Vps 45 Play Pivotal Roles in Assembly of the Pellicle and Sub‐Pellicle Network in Toxoplasma gondii .” Molecular Microbiology 113, no. 1: 208–221. 10.1111/mmi.14411.31670849 · doi ↗ · pubmed ↗
- 8Cerede, O. , J. F. Dubremetz , M. Soete , et al. 2005. “Synergistic Role of Micronemal Proteins in Toxoplasma gondii Virulence.” Journal of Experimental Medicine 201: 453–463.15684324 10.1084/jem.20041672 PMC 2213027 · doi ↗ · pubmed ↗