Genomic Complexity of ccdc40 in Xenopus : Implications for CRISPR Targeting and Disease Modeling
Takuya Nakayama, Saurabh Kulkarni

TL;DR
This paper explores the genomic structure of CCDC40 in Xenopus frogs to better understand its role in a human disease and improve genetic modeling.
Contribution
The study reveals two distinct ccdc40 genes in Xenopus tropicalis and identifies a single homoeolog in Xenopus laevis suitable for genetic studies.
Findings
Xenopus tropicalis has two distinct ccdc40 genes near the previously annotated locus.
Xenopus laevis contains only one homoeolog (ccdc40.S), making it a better model for studying CCDC40.
The findings may help in understanding CCDC40 variants linked to primary ciliary dyskinesia.
Abstract
Mutations in CCDC40 cause primary ciliary dyskinesia in humans. To evaluate the pathogenicity of variants in CCDC40 , we examined the genomic structure of this gene in Xenopus tropicalis , a diploid frog suitable as a model for genetic studies. We identified inconsistencies in the current ccdc40 gene model and discovered two distinct ccdc40 genes near the previously annotated locus. Surprisingly, Xenopus laevis , an allotetraploid species that typically has two homoeologs, contains only one homoeolog ( ccdc40.S ), making it a more suitable genetic model for studying ccdc40 function and potentially expediting the functional characterization of CCDC40 variants linked to primary ciliary dyskinesia.
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
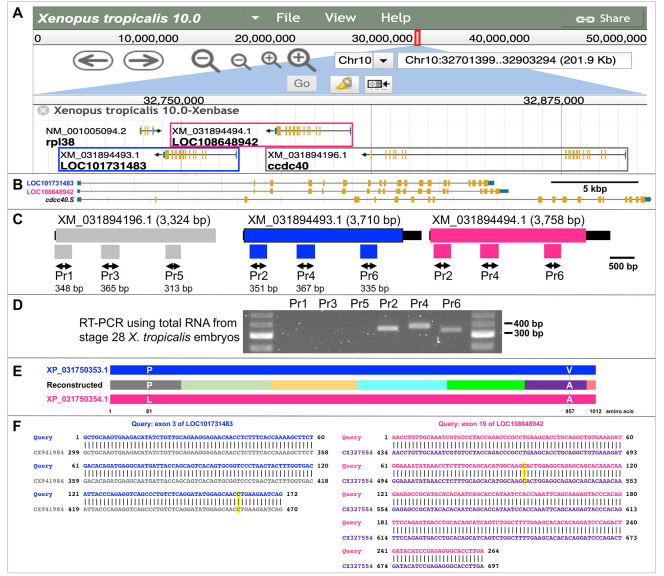 Figure 1
Figure 1|
Primers: sequence (5' to 3') |
|
|
| |
|
Pr Set 1 |
Pr1_5Pr: CCCATCTCACCCTGCACTAT * Pr1_3Pr: ATGACCCCACCACTCCTGA |
8-355(348 bp) |
N/A |
N/A |
|
Pr Set 3 |
Pr3_5Pr: GAAAAAAAAACAGGATATTTTTGTGG Pr3_3Pr; CTGCACGGTTCAACATTAGC |
972-1,336(365 bp) |
N/A |
N/A |
|
Pr Set 5 |
Pr5_5Pr: TGCAGGAAAATCTACGAAACC ** Pr5_3Pr: CCTGTTTCACCTGATTCTTCAA |
2,306-2,618(313 bp) |
N/A |
N/A |
|
Pr Set 2 |
Pr2_5Pr: CCCATCTCACCCTGCACTAT * Pr2_3Pr: CCACCACTCCTGTTTCACCT |
N/A |
133-483(351 bp) |
83-433(351 bp) |
|
Pr Set 4 |
Pr4_5Pr: AAAAAAGCAGGATATTTTTGTGG Pr4_3Pr: TGCACGGTTCAACATGAAGT |
N/A |
1,115-1,481(367 bp) |
1,065-1,431(367 bp) |
|
Pr Set 6 |
Pr6_5Pr: TGCAGGAAAATCTACGAAACC ** Pr6_3Pr: CCTACGAAAGTCACTGACGGTTA |
N/A |
2,452-2,786(335 bp) |
2,402-2,736(335 bp) |
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsGenetics, Bioinformatics, and Biomedical Research
Description
CCDC40 is a coiled-coil domain-containing protein 40 that is essential for establishing the left-right axis and proper motile cilia function in the airway. Consistently, loss-of-function mutations in CCDC40 have been shown to cause defects in motile cilia function in humans, mice, zebrafish, and Xenopus tropicalis (Becker-Heck et al., 2011; Bhattacharya et al., 2015), leading to primary ciliary dyskinesia (PCD) in humans (DOID:9562; MIM:244400).
The emergence of gene-editing technologies, particularly Clustered Regularly Interspaced Short Palindromic Repeats/CRISPR-associated protein 9 (CRISPR/Cas9), has revolutionized genome manipulation methods. CRISPR/Cas9 has been established in both Xenopus tropicalis (Blitz et al., 2013; Nakayama et al., 2013) and Xenopus laevis (Wang et al., 2015), demonstrating their suitability as models for developmental genetics alongside classical embryology research. Due to its simpler diploid genome (Hellsten et al., 2010), X. tropicalis holds promise for modeling human genetic diseases (Hwang et al., 2019; Willsey et al., 2024), especially ciliopathies (Rao & Kulkarni, 2021), in contrast to the allotetraploid genome of X. laevis (Session et al., 2016).
During our research to investigate the pathogenicity of variants of unknown significance (VUS) in the ccdc40 gene (GeneID:100486955) affecting motile cilia function in X. tropicalis , we used CRISPR targeting and encountered significant challenges. Specifically, identifying a unique CRISPR/Cas9 target site without potential off-target effects was difficult (see (Blitz & Nakayama, 2022) for the strategy). Upon examination, we found that the previously published CRISPR/Cas9 target sequence, including its PAM site (Bhattacharya et al., 2015), appeared four times throughout the genome, raising concerns about off-target activity. Further complicating our analysis, the forward and reverse primers previously used for genotyping, designed to amplify an approximately 200 bp region containing the CRISPR/Cas9 target site, also matched four distinct genomic locations. Additional investigation using Xenbase (Fisher et al., 2023) confirmed these four genomic matches: two within the ccdc40 gene itself (referred to as the "former" ccdc40 , detailed further below), one in the gene LOC101731483 (GeneID:101731483), and one in LOC108648942 (GeneID:108648942) ( Figure 1A ). This suggests that the published genotyping results (Bhattacharya et al., 2015) must be mixed with data from four different genomic regions.
In X. laevis , despite its allotetraploid genome, the ccdc40 gene exists as a singleton found only in the S genome ( ccdc40.S [GeneID:108702701], XM_018238324.2; Xenbase, X. laevis 10.0). The predicted structure of ccdc40.S comprises 19 exons ( Figure 1B ). The genes of X. tropicalis LOC108648942 and LOC101731483 also contain 19 exons ( Figure 1B ), while the "former" ccdc40 gene has 22 exons. Notably, this "former" ccdc40 gene features six exons that are repeated. For example, especially "100% identity" between exon 2 and exon 16, exon 6 and exon 20, and exon 7 and exon 21 was observed. Moreover, the previously published CRISPR target site (Bhattacharya et al., 2015) appears twice within this gene—in exon 4 and exon 18—exhibiting 96.81% identity. Consequently, exons from 16 to 21 appear to be duplicated versions of exons 2 to 7, showing 96.81% to 100% identity between corresponding exons.
We decided to perform RT-PCR analysis to evaluate gene expression further and detect transcripts from these genes. Due to the high similarity between LOC108648942 and LOC101731483 (99.75% identity based on their predicted cDNAs, XM_031894494.1 and XM_031894493.1 from NCBI, (Sayers et al., 2024)), distinguishing between their transcripts by RT-PCR proved challenging. However, it was possible to design primers specific to the "former" ccdc40 (predicted) mRNA sequence (XM_031894196.1), which is 98.92 and 98.96% identical to XM_031894494.1 and XM_031894493.1, respectively. We selected three similar regions from the 5' to 3' ends of each predicted mRNA for amplification ( Figure 1C ). Our RT-PCR results ( Figure 1D ) indicate that transcripts from the primer regions matching LOC108648942 and LOC101731483 were detectable, while transcripts from the "former" ccdc40 gene were not detectable at developmental stage 28, a critical period when multiciliated cells (MCCs) containing motile cilia mature (Kulkarni et al., 2021; Kulkarni et al., 2018).
To further confirm the expression of LOC108648942 and LOC101731483, we performed a BLAST search in the NCBI database for Expressed Sequence Tags (ESTs) matching their predicted cDNAs. We identified multiple ESTs (e.g., CX941984, AL632867, CN086103, CX937961, DT449052, CX327554, EL834361) and successfully reconstructed a complete potential mRNA sequence corresponding to LOC108648942 and LOC101731483 from 5' to 3'. Although we also could identify some ESTs that overlapped with all three predicted cDNAs due to high similarity, we could not reconstruct a complete mRNA sequence corresponding to the "former" ccdc40 (XM_031894196.1). The ESTs we identified were derived from various tissues, including tadpole brains and spinal cords, adult spleen and testes, and whole embryos, suggesting widespread expression of LOC108648942 and LOC101731483.
The reconstructed mRNA encodes 1,012 amino acids, which is the same length as LOC108648942 and LOC101731483, corresponding to XP_031750354.1 and XP_031750353.1, with an amino acid identity of 99.90% ( Figure 1E ). In contrast, the "former" ccdc40 (XP_031750056.1) encodes 1,078 amino acids, displaying a slightly lower identity of 98.96% compared to the reconstructed sequence. When compared to Ccdc40.S from X. laevis (XP_018093813.1, 1,045 amino acids), LOC108648942 and LOC101731483 exhibit approximately 85% identity, while the "former" ccdc40 shows around 82% identity. Considering gene structures, mRNA expression data, and protein sequence similarities, we suggest that the "former" ccdc40 gene is likely not functional. Instead, we propose that LOC108648942 and LOC101731483 represent the authentic ccdc40 genes in the X. tropicalis genome, although further evidence is needed to confirm this conclusively.
When comparing LOC108648942 and LOC101731483 (XM_031894494.1/XM_031894493.1), several single-nucleotide polymorphisms (SNPs) were identified in the protein-coding regions and untranslated regions (UTRs). Some SNPs resulted in silent mutations, while others caused missense mutations. We focused on missense mutations to clearly distinguish the two genes. LOC108648942/XP_031750354.1 encodes the amino acids L81 and A957, whereas LOC101731483/XP_031750353.1 encodes P81 and V957 at the corresponding positions ( Figure 1E ). EST analysis further supported transcription from both genes; one EST (CX941984) contained a SNP corresponding to LOC101731483 in exon 3 encoding P81, and another EST (CX327554) corresponded to LOC108648942 in exon 19 encoding A957 ( Figure 1E, F). Given these observations and their closer similarity to X. laevis Ccdc40.S (that contains L81 and A990 at the corresponding sites), we propose renaming LOC108648942 as X. tropicalis ccdc40 and LOC101731483 as ccdc40.2 , following Xenopus gene nomenclature.
The Harland group (Grammer et al., 2005) reported a naturally occurring background mutant known as " grinch " in X. tropicalis Nigerian strain colonies from various laboratories, including the Grainger lab. These mutants exhibit significant ventral edema. Subsequently, the Khokha lab (del Viso et al., 2012) identified ciliary defects in this mutant, linking them to multiple mutations in the " ccdc40 " gene based on genome data v4.1 and v7.1. They predicted that the gene had 18 exons, in contrast to the 19 exons we identified based on genome version 10.0 ( Figure 1B ); however, they correctly predicted the encoded protein length to be 1,012 amino acids.
Notably, although the phenotype observed by Grammer et al. (2005) suggested simple Mendelian inheritance (a single recessive allele resulting in a 25% mutant phenotype), the mutant cDNAs analyzed by del Viso et al. (2012) revealed multiple mutations (Figure 4b in del Viso et al., 2012), indicating the presence of more than one mutant allele. Given our findings presented here, some of their results might be explained by the existence of two separate ccdc40 genes. Therefore, a detailed re-examination of the grinch mutation locus would be valuable in the future, explicitly considering the two-gene scenario. Additionally, we noted that the mutant cDNAs and predicted proteins described by del Viso et al. (2012; Supplementary Figure 5) exhibit amino acids representative of both genes (L81 from LOC108648942/ ccdc40 and P81 from LOC101731483/ ccdc40.2 ), again supporting transcriptional activity of both genes.
The loss of the ccdc40 gene from the L genome of X. laevis and the genomic complexity (three similar genes and naturally occurring mutations such as grinch ) in the ccdc40 -containing region of X. tropicalis may indicate that this genomic region is prone to mutation in Xenopus species. It would thus be insightful to investigate this region across other Xenopus species and additional X. tropicalis strains beyond the Nigerian strain.
In summary, the existence of two *ccdc40 * genes ( ccdc40 and ccdc40.2 ) in X. tropicalis complicates genotyping and CRISPR targeting. Therefore, we suggest that X. laevis , which has only one ccdc40.S gene, offers a simpler and more suitable genetic model for studying Ccdc40 function.
Methods
Frogs
*X. tropicalis * colony is from our in-house breeding stocks at the University of Virginia, which are not highly inbred but originated from the same ancestors as the Nigerian inbred line used for genome sequencing (Hellsten et al., 2010). Frogs ( Xenopus tropicalis ) were bred and housed in a vivarium using protocols (ACUC# 4295, 1472) approved by the University of Virginia Institutional Animal Care and Use Committee (IACUC). Embryos needed for experiments were generated using in vitro fertilization as described before (Nakayama & Grainger, 2023; Rao et al., 2025). Briefly, the testes from male frogs were crushed in 1x MBS (pH 7.4-7.5) with 0.1-0.2% BSA and added to eggs obtained from the female frogs. After 1-3 minutes of incubation, freshly made 0.1x MBS (pH 7.4-7.5) was added, and the eggs were incubated for 10 more minutes till contraction of the animal pole of the eggs was visible. The jelly coat was removed using 2-3% cysteine in 0.1 x MBS solution (pH 7.8-8.0) for 3-5 minutes (until the jelly coat was removed).
RNA extraction and cDNA synthesis (for RT-PCR)
Five embryos at stage 28 (staged according to (Nieuwkoop, 1994)) were pooled and subjected to total RNA extraction using Quick-RNA ^TM^ Miniprep Kit (Zymo Research), following the manufacturer's instructions. 5 ng of the resultant total RNA was used to make cDNA using Verso cDNA Synthesis Kit (Thermo Scientific™), following the manufacturer's instructions
PCR
PCR reactions used EmeraldAmp® GT PCR Master Mix (Takara Bio), following the manufacturer's instructions. Each 12.5 μl reaction contained 0.5 μl cDNA and specific primer sets (Figure 1). For PCR, after denaturing the PCR mixture at 94°C for 5 min, the condition of one PCR cycle was as follows: denaturing at 94°C for 20 sec; annealing (see below for temperature) for 20 sec; extension at 68°C for 12 sec. Touchdown PCR (Korbie & Mattick, 2008) was performed, namely, by gradually reducing the annealing temperature (initially set at 65°C) in 0.5°C increments per cycle for 13 cycles, followed by additional cycles at a constant annealing temperature of 58°C. The number of additional cycles required for each primer set was determined for each primer set (see Table 1) by trial and error, i.e., 26 cycles for Pr Set1 and 2; 19 cycles for Pr Set 3 and 4; 17 cycles for Pr Set5 and 6. Subsequently, 5 μl of the PCR product for each primer set was loaded for electrophoresis using 2% agarose gel by the standard method.
** In silico assay **
Homology searches for short sequences like CRISPR target sites or primer were conducted using GGGenome (https://gggenome.dbcls.jp/). Gene structure and BLAST searches were performed with Xenbase (http://www.xenbase.org/, RRID:SCR_003280) and NCBI databases (https://www.ncbi.nlm.nih.gov/). PCR primers were designed using Primer3 (v. 0.4.0) (https://bioinfo.ut.ee/primer3-0.4.0/).
Reagents
Table 1. RT-PCR primers
**: **
*, commonly used for Pr Set1 and 2; **, commonly used for Pr Set5 and 6.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Becker-Heck A Zohn IE Okabe N Pollock A Lenhart KB Sullivan-Brown J Mc Sheene J Loges NT Olbrich H Haeffner K Fliegauf M Horvath J Reinhardt R Nielsen KG Marthin JK Baktai G Anderson KV Geisler R Niswander L Omran H Burdine RD 2010125 The coiled-coil domain containing protein CCDC 40 is essential for motile cilia function and left-right axis formation.Nat Genet 4311061-4036798410.1038/ng.72721131974 PMC 3132183 · doi ↗ · pubmed ↗
- 2Bhattacharya D Marfo CA Li D Lane M Khokha MK 2015114 CRISPR/Cas 9: An inexpensive, efficient loss of function tool to screen human disease genes in Xenopus.Dev Biol 40820012-160619620410.1016/j.ydbio.2015.11.00326546975 PMC 4684459 · doi ↗ · pubmed ↗
- 3Blitz IL Biesinger J Xie X Cho KW 20131121 Biallelic genome modification in F(0) Xenopus tropicalis embryos using the CRISPR/Cas system.Genesis 51121526-954X 82783410.1002/dvg.2271924123579 PMC 4039559 · doi ↗ · pubmed ↗
- 4Blitz IL Nakayama T 202231 CRISPR-Cas 9 Mutagenesis in Xenopus tropicalis for Phenotypic Analyses in the F(0) Generation and Beyond.Cold Spring Harb Protoc 2022310.1101/pdb.prot 10697134244352 · doi ↗ · pubmed ↗
- 5del Viso F Bhattacharya D Kong Y Gilchrist MJ Khokha MK 20121121 Exon capture and bulk segregant analysis: rapid discovery of causative mutations using high-throughput sequencing.BMC Genomics 1364964910.1186/1471-2164-13-64923171430 PMC 3526394 · doi ↗ · pubmed ↗
- 6Fisher M James-Zorn C Ponferrada V Bell AJ Sundararaj N Segerdell E Chaturvedi P Bayyari N Chu S Pells T Lotay V Agalakov S Wang DZ Arshinoff BI Foley S Karimi K Vize PD Zorn AM 202354 Xenbase: key features and resources of the Xenopus model organism knowledgebase.Genetics 22410016-673110.1093/genetics/iyad 01836755307 PMC 10158840 · doi ↗ · pubmed ↗
- 7Grammer TC Khokha MK Lane MA Lam K Harland RM 200531 Identification of mutants in inbred Xenopus tropicalis.Mech Dev 12230925-477326327210.1016/j.mod.2004.11.00315763207 · doi ↗ · pubmed ↗
- 8Hellsten U Harland RM Gilchrist MJ Hendrix D Jurka J Kapitonov V Ovcharenko I Putnam NH Shu S Taher L Blitz IL Blumberg B Dichmann DS Dubchak I Amaya E Detter JC Fletcher R Gerhard DS Goodstein D Graves T Grigoriev IV Grimwood J Kawashima T Lindquist E Lucas SM Mead PE Mitros T Ogino H Ohta Y Poliakov AV Pollet N Robert J Salamov A Sater AK Schmutz J Terry A Vize PD Warren WC Wells D Wills A Wilson RK Zimmerman LB Zorn AM Grainger R Grammer T Khokha MK Richardson PM Rokhsar DS 2010430 The genome of the Western clawed frog Xenopus tropicalis.Sc · doi ↗ · pubmed ↗