Protein Extraction from Chlorella pyrenoidosa Using Bacillus spp. Isolated from Jeotgal: Strain isolation, Characterization, and Fermentation
Kyung-Jin Cho, Min-Ung Kim, Geum-Jae Jeong, Do Kyung Oh, Ju-Hong Kang, Da-Hyeon Yoon, Fazlurrahman Khan, Young-Mog Kim

TL;DR
Researchers isolated Bacillus strains from Korean fermented food to improve protein extraction from Chlorella pyrenoidosa, a microalga with a rigid cell wall.
Contribution
A novel microbial fermentation approach using Bacillus spp. for efficient protein extraction from Chlorella pyrenoidosa is proposed.
Findings
Eight Bacillus strains were selected for high enzymatic activity, including cellulases, proteases, and lipases.
Bacillus amyloliquefaciens F2 achieved the highest protein extraction yield of 35.45 ± 1.21%.
Four strains did not produce biogenic amines, indicating potential safety for food industry applications.
Abstract
Chlorella pyrenoidosa, a versatile microalga with a rich nutritional profile and functional components, has various applications. However, rigid cell walls pose challenges for the effective extraction of proteins. Microbial fermentation is a promising solution for large-scale production and industrial applications. This study aimed to isolate Bacillus spp. with high enzymatic activity from Jeotgal, a Korean traditional fermented food, and enhance protein extraction from C. pyrenoidosa using microbial fermentation with the isolated Bacillus spp. Twenty-two strains of Bacillus spp. were isolated, and eight Bacillus species were selected based on their ability to produce cellulases, proteases, and lipases. Microbial safety was further assessed by testing for biogenic amine production and hemolytic activity. All eight strains exhibited γ-hemolysis, with four strains not producing biogenic…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
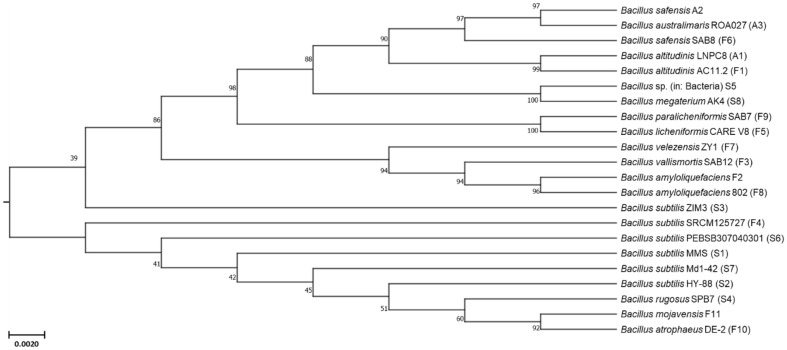 Figure 1
Figure 1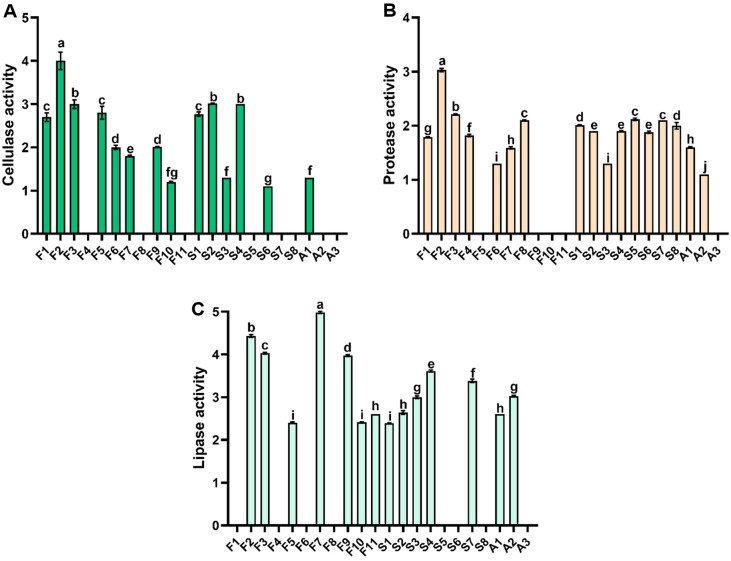 Figure 2
Figure 2Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsAlgal biology and biofuel production
Introduction
As the interest in sustainable food sources increases, marine biomass is emerging as a nutritious alternative to existing food materials. Marine biomass refers to all ocean organisms that can be utilized as resources. It has been reported that they are beneficial to the human body and emit less carbon dioxide [1, 2]. Chlorella pyrenoidosa (C. pyrenoidosa) is a green microalga containing various minerals (magnesium, calcium, iron, and potassium), vitamins, omega-3 fatty acids, and essential amino acids. In particular, C. pyrenoidosa has a high protein content and has attracted attention as a future food resource [3, 4]. However, it has a rigid cell wall, which makes the digestion and absorption of nutrients difficult. Physical methods such as ultrasonication [5], sonication [6], freeze-thawing [7], and high-pressure treatment [8], and chemical methods such as organic solvent extraction [9], salt precipitation [10], and enzyme treatment [11], have been used to extract proteins from C. pyrenoidosa. Microbial fermentation is carried out at low temperatures and pressures, offering advantages such as reduced ingredient denaturation, lower energy consumption and costs compared with other extraction methods, and minimal environmental impact [12]. Owing to these advantages, microbial fermentation has been used to extract plant proteins. Dai et al. (2022) effectively extracted proteins from soybean meal by fermentation with Bacillus licheniformis and Bacillus subtilis [13], resulting in an increase in the phenol and pyrazine contents. Xiao et al.(2009) extracted crude proteins from macroalgae via fermentation with Aspergillus niger and Candida utilis [14].
Bacillus species are among the most important industrial microorganisms that produce cellulases, proteases, lipases, and glucanase [15, 16]. In addition, Bacillus spp. have been reported to exhibit functional properties, such as antioxidant and probiotic activities [17?-19]. Many studies have been conducted to isolate Bacillus spp. from traditional Asian fermented foods such as Cheonggukjang, Natto, Tempeh, and Jeotgal [20???-24]. Bacillus spp. isolated from Jeotgal, a traditional Korean high-salinity fermented food, possess various enzymatic systems capable of breaking down complex biomolecule [25, 26]. This study aimed to isolate Bacillus spp. from Jeotgal exhibiting exceptional enzymatic activities and to assess their safety and antimicrobial activity. Additionally, we evaluated the potential of these Bacillus isolates as a biological resource for protein extraction from C. pyrenoidosa.
Material and Methods
Materials and Microorganisms
Three types of Jeotgal (flounder, squid, and anchovy) were purchased from the Jeju Dongmun Market in Korea. C. pyrenoidosa powders were obtained from Jeongwoodang Co., Ltd., (Republic of Korea). All other chemicals and reagents were of analytical grade and included the following: L-histidine, L-lysine, L-ornithine monohydrochloride (L-ornithine), L-arginine, L-tyrosine, and carboxymethyl cellulose (CMC) from Sigma-Aldrich (USA), sodium chloride (NaCl) and D-glucose from Junsei Chemical Co., Ltd., (Japan), and bromocresol purple, congo red, pyridoxal 5-phosphate monohydrate, and mineral oil from Duksan Pure Chemicals Co., Ltd., (Republic of Korea). Escherichia coli KCTC 2571, Bacillus cereus KCTC 1092, Staphylococcus aureus KCTC 3881, and Listeria monocytogenes KCTC 3569 were obtained from the Korean Collection for Type Culture (KCTC, Republic of Korea). Pseudomonas aeruginosa KCCM 11321 was obtained from the Korean Culture Center of Microorganism (KCCM, Republic of Korea). These bacteria were maintained in glycerol stocks at -80°C and used in the experiment
Isolation of Bacillus spp. from Jeotgal
The isolation of Bacillus spp. from Jeotgal was carried out using previously described procedures with some modifications [27]. Jeotgal (25 g) was combined with 225 ml of 0.1 M phosphate-buffered saline PBS (pH 7.2) and homogenized in a stomacher (BagMixer 400 W; Interscience, France) for 4 min. The suspension was serially diluted and inoculated onto all culture agar (ACA, Sigma-Aldrich) to isolate the bacteria. The inoculated ACA plates were incubated aerobically and anaerobically at 37 ± 1°C for 24 ± 2 h. Colonies were selected based on their morphological characteristics such as size, color, shape, and presence of bubbles. The colonies were streaked onto ACA plates and incubated under the same conditions. This process was repeated until a single colony was formed. The isolated strains were stored in a glycerol stock at -80°C and used in the experiment.
DNA Extraction, PCR Amplification and Identification
DNA was extracted from a single colony using a DNA extraction kit (Bioneer Co., Ltd., Republic of Korea). Subsequently, polymerase chain reaction (PCR) was performed using the extracted DNA, AccuPower PCR premix (Bioneer), and universal primers (27F, 5’-AGAGTTTGATCTGGCTCAG-3’; 1492R, 5’-TACGGTTAC CTTGTTACGACTT-3’). The PCR protocol consisted of 30 cycles: denaturation at 94°C for 1.5 min, annealing at 55°C for 0.5 min, and extension at 72°C for 4 min in a PCR thermal cycler (Takara, Republic of Korea). The 16S rRNA sequences of the PCR products were analyzed for molecular biological identification. The sequences were edited and assembled into contigs using BioEdit Sequence Alignment Editor (Table S1). All contigs were compared against nucleotide sequences in the GenBank database (https://blast.ncbi.nlm.nih.gov/Bl-ast.cgi) using the Basic Local Alignment Search Tool (BLAST) search algorithm. Strains were identified by selecting those showing the highest sequence similarity (at least 99.80%) to the previously reported standard strains.
Enzyme Activity
The enzymatic activities of Bacillus spp. were measured for protease, cellulase, and lipase. All enzyme activities were determined by comparing the size of the clear zone formed after incubation with the colony or agar well size (Fig. S1A), using the following formula:
Enzyme activity (ratio) = Clearance zone size (mm) / Colony size (mm)
Protease Activity
The Protease activity was analyzed according to a previously described method [28]. Bacillus spp. was inoculated into ACA supplemented with 1% (w/v) skim milk (BD Difco, USA). Subsequently, the inoculated plates were incubated at 37°C for 24 h.
Cellulase Activity
Cellulase activity was analyzed using carboxymethyl cellulose (CMC) agar as described by Liang et al. (2014)[29]. CMC agar was prepared with the following medium composition: peptone 10 g/l, CMC 10 g/l, MgSO_4_ 0.2 g/l, NaCl 0.5 g/l, CaCl_2_ 0.1 g/l, Agar 15 g/l dissolved in distilled water. Then, the medium was sterilized by autoclaving at 121°C for 15 min. Bacillus spp. were inoculated onto CMC agar and incubated at 37°C for 48 h. Then, the plates were stained with 1% (w/v) Congo red solution for 15 min and destained with 1 M NaCl solution for 15 min
Lipase Activity
The lipase activity was assessed using a primary plate-screening assay [30]. Agar plates were prepared with the following medium composition: 20 g/l agar, 20 g/l Tween 80, and 0.1 g/l methyl red dissolved in distilled water. Agar wells with a diameter of 0.5 cm were made in each plate using a sterilized microtip. The strains were cultured in ACB at 37°C for 24 h and centrifuged at 13,000 g for 15 min (Hanil Supra R22; Republic of Korea). A 25 μl aliquot of the supernatant was inoculated into the agar wells and cultured at 37°C for 24 h.
Antibacterial Activity
The antibacterial activity of Bacillus spp. isolated from Jeotgal was evaluated against five pathogenic bacteria using a modified Kirby-Bauer assay [31]. Each pathogen and Bacillus spp. were inoculated separately into Muller Hinton broth (MHB, BD Difco) and cultured at 37°C for 24 h. The pathogen culture, initially at a concentration of 10^8^ CFU/ml, was diluted to 10^5^ CFU/ml using MHB and spread on Mueller Hinton agar (MHA, BD Difco). The pour plate method was employed to determine the concentration of viable cell count. Then, 100 μl of cultured Bacillus spp. was inoculated onto a paper disk (8 mm; ADVANTEC, Japan) and aseptically transferred to an MHA plate. The MHA plate was incubated at 37°C for 24 h, and the diameter of the inhibition zone was measured in mm. Antibacterial activity was determined by comparing the size of the clearance zone formed after incubation with the colony size (Fig. S1B) using the following formula:
Antibacterial activity (ratio) = Clearance zone size (mm) / Colony size (mm)
Safety Evaluation of Bacillus spp.
Hemolytic activity and biogenic amine productivity were analyzed to assess the microbial safety of Bacillus spp. isolated from Jeotgal. Hemolytic activity was determined by examining the clearance zone of ACA containing 5%sheep blood (v/v; KisanBio) [32]. The Bacillus spp. were plated on ACA and incubated at 37°C for 24 h. Biogenic amine productivity was analyzed for histamine, cadaverine, tyramine, and putrescine according to the method described by Yilmaz et al. (2022) [33]. The decarboxylase broth was prepared by adding pyridoxal 5-phosphate (5 mg/l) and 1% L-arginine, L-tyrosine, L-histidine, or L-lysine. Bacillus spp. were inoculated into each prepared broth and incubated anaerobically at 37°C for 24 h. The medium change from yellow to purple was determined as positive.
Fermentation of C. pyrenoidosa for Protein Extraction
C. pyrenoidosa was suspended and fermented with the isolated Bacillus spp. to assess the enzymatic activity of each strain. Initially, 5% (w/v) C. pyrenoidosa medium was prepared by mixing 10 g of C. pyrenoidosa powder with 200 ml of distilled water. Subsequently, 1% (v/v) of each Bacillus strain, at a concentration of 10^7^ CFU/ml, was inoculated into the medium and cultured at 37°C with shaking at 150 rpm for 24 h. The pour plate method was employed to determine the concentration of Bacillus strains. A control experiment (without inoculation) was also conducted under identical conditions. The culture medium was then centrifuged at 10,000 g for 20 min to separate the supernatant, which was used to compare the protein extraction yields. The protein concentration was determined using the bicinchoninic acid (BCA) assay, employing the Pierce BCA Protein Assay Kit (Thermo Fisher Scientific, USA), as described earlier [34]. A standard curve was generated for protein quantification using bovine serum albumin. The protein yield from C. pyrenoidosa was calculated using the following equation:
Protein extraction yield (%) = P1/P0 × 100%
Where P1 is the protein content of the fermented C. pyrenoidosa extract (mg/ml) and P0 is the protein content of the uninoculated C. pyrenoidosa medium (mg/ml).
Statistical Analysis
All experiments were conducted in triplicates. The results were analyzed using SPSS 27 (Statistical Package for Social Science, USA) to calculate the mean and standard deviation of each test group and Duncan’s multiple range test. Statistically significant differences were observed at the 5% significance level (p < 0.05).
Results and Discussion
Identification of Isolated Bacillus spp.
The identification results for twenty-two Bacillus spp. isolated from the three types of Jeotgal are shown in Table 1. Eleven Bacillus strains were isolated from flounder Jeotgal, eight from squid Jeotgal, and three from anchovy Jeotgal. The 16s rRNA gene sequences of the twenty-two isolates were identified as follows: B. subtilis (six species), B. altitudinis (two species), B. amyloliquefaciens (two species), B. safensis (two species), B. megateruim (two species), B. licheniformis (one species), B. paralicheniformis (one species), B. rugosus (one species), B. atrophaeus (one species), B. mojavensis (one species), B. vallismortis (one species), B. velezensis (one species), B. australimaris (one species), and Bacillus sp. (unknown; one species), and were named F1-F11, S1-S8, and A1-A3. A phylogenetic tree was constructed to represent genetic relationships among the twenty-two isolates (Fig. 1). The phylogenetic tree was drawn to scale, and branch lengths were expressed in the same units as the evolutionary distance used to infer the phylogenetic tree [35]. Guan et al. (2011) analyzed bacterial communities in anchovy and shrimp Jeotgal and identified various Bacillus spp., suggesting that these bacteria play an important role in fermentation [36]. In addition, it has been reported that Bacillus spp. isolated from high-salinity Jeotgal exhibit high protease activity [37, 38]. Yin et al. (2024) found that an environment with 10% salinity enhanced the secretion of extracellular polymeric substances (EPS) from Bacillus spp. with notable expression of the DegS gene [39]. DegS is crucial for activating the expression of the aprE gene, which is involved in the production of alkaline proteases. Consequently, Bacillus spp. isolated from high-salt Jeotgal are well-suited for decomposing marine biomass, such as C. pyrenoidosa.
Selection and Evaluation of Enzyme Activities of Isolated Bacillus spp.
The enzymatic activities of Bacillus spp. isolated from Jeotgal are shown in Fig. 2. Among the twenty-two Bacillus strains, eighteen strains showed cellulase activity, fourteen strains showed protease activity, and fourteen strains showed lipase activity. All strains showed enzymatic activity for at least one substrate and eight strains (F2, F3, F7, S1, S2, S3, S4, and A1) showed enzymatic activity for all three substrates. In particular, strain F2, which was identified as B. amyloliquefaciens, showed the highest enzyme activity, with values of 4.00 ± 0.20 for protease, 3.03 ± 0.03 for cellulase, and 4.43 ± 0.03 for lipase. B. amyloliquefaciens is a representative cellulolytic bacterium known to have high cellulase activity and to produce various hydrolytic enzymes, such as lipase, α-amylase, and protease [40??-43]. In addition, B. amyloliquefaciens produces hydrolytic enzymes in the food industry due to its thermostable enzyme productivity [44]. Zhang et al. (2023) previously reported the use of Bacillus spp. as microbial starters to extract proteins from C. pyrenoidosa [45]. In addition, Kim et al. (2012) reported that the enzyme activities of Bacillus spp. varied depending on the source of isolation [46]. Thus, the eight Bacillus spp. isolated from Jeotgal, which possess various enzyme systems (including cellulase, protease, and lipase activities), could serve as microbial starters capable of breaking down complex biomolecules such as C. pyrenoidosa [25].
Evaluation of the Antibacterial Activity of Eight of Bacillus spp. Possessing Various Enzymatic Activity
During fermentation, the antibacterial properties of Bacillus spp. are crucial for inhibiting the growth of harmful pathogens, thereby enhancing food safety and ensuring consistent fermentation quality [47]. Table 2 presents the antibacterial activities of the eight Bacillus strains against the five types of pathogens. All the selected Bacillus spp., except S1, demonstrated antibacterial activity against L. monocytogenes. Notably, strains F2, F7, and S4 exhibited antibacterial effects against S. aureus, with F2 exhibiting the strongest activity. Some Bacillus spp. produce antibacterial substances [48]. Interestingly, only strain F2 showed antibacterial activity against the gram-negative bacterium E. coli. B. amyloliquefaciens F2 demonstrated the broadest and most potent antibacterial activity, effectively inhibiting E. coli, S. aureus, L. monocytogenes, and P. aeruginosa. This result aligns with those of previous studies, which indicated that B. amyloliquefaciens generally exhibit superior antibacterial activity against pathogens compared to other Bacillus species [49, 50].
Microbial Safety Analysis of Bacillus spp.
The results of the microbial safety assessment of the eight Bacillus isolates are presented in Table 3. Hemolysis refers to the destruction of red blood cells by substances produced by microorganisms that can lead to abnormal red blood cell function [51]. All eight Bacillus isolates were identified as γ-type (data non-shown), indicating no hemolytic activity [52].
Analysis of the biogenic amine productivity of Bacillus spp. showed that F3 and F7 produced putrescine, whereas S1 produced cadaverine and tyramine (Table 3). S3 produced putrescine, cadaverine, and tyramine. In contrast, the strains F2, S2, S4, and A1 did not produce biogenic amines. Moon et al. (2015) reported that B. subtilis and B. licheniformis isolated from Cheonggukjang, a Korean fermented food, do not produce biogenic amines [38]. Lee et al. (2017) also reported that three strains of B. amyloliquefaciens isolated from soy sauce did not produce biogenic amines [53]. Biogenic amines can be found in fermented foods such as dairy and soybean products and can be harmful to humans when consumed in large amounts [54]. Therefore, the strains F2, F2, S4, and A1, which do not produce biogenic amines, not only help maintain the quality of fermented foods, but also exhibit low pathogenicity risk, and can be easily applied to commercial fermentation processes.
Protein Extraction of C. pyrenoidosa for Utilizing Food Materials
Table 4 presents the protein extraction yields from C. pyrenoidosa using the eight Bacillus spp. as fermentation starters. Strain F2 showed the highest protein extraction yield of 34.45 ± 1.21%, and F3 showed the second highest yield of 21.96±0.78% (p < 0.05). Next, S2 showed 16.46 ± 1.02%; S4, 13.93±0.6%; F7, 13.67±0.7%; S1, 7.78±0.29%; S3, 6.99 ± 1.24%; and A1, 5.66 ± 0.01%, in that order. The high protein extraction yields for F2 and F3 are attributed to their superior cellulase and protease activities (Fig. 2). Protein extraction yields from C. pyrenoidosa fermentation by three strains of B. subtilis (S1, S2, and S3) ranged from 6.99 ± 1.24% to 16.46 ± 1.02%. These differences were attributed to variations in enzyme activity, despite all the strains belonging to the same species. The C. pyrenoidosa cell wall is composed of polysaccharides and glycoproteins [55], making it susceptible to decomposition by extracellular enzymes, such as cellulase and proteases, produced during fermentation. This is supported by results indicating that strain F2, which possessed the highest cellulase, protease, and lipase activities, effectively extracted proteins from C. pyrenoidosa. In addition, our previous study confirmed that the protein extraction yield from C. pyrenoidosa was approximately 35% when using proteases derived from Bacillus spp. [55]. This result is similar to the C. pyrenoidosa protein extraction yield using strain F2, suggesting that microbial fermentation can effectively decompose C. pyrenoidosa cell walls. Safi et al. (2014) reported that the protein extraction yield of Chlorella vulgaris was 33.20% when alkaline treatment was performed for 2 h at pH 12 [56], and Wang and Jang (2012) reported that the protein extraction yield of C. pyrenoidosa was 22.90% after freeze-thawed and sonicated [57]. This suggests that Bacillus spp. treatment is an efficient method for C. pyrenoidosa protein extraction. Consequently, microbial fermentation, such as B. amyloliquefaciens F2, has the potential to expand the commercial utilization of marine protein sources, such as C. pyrenoidosa. In future, it will be necessary to study the optimal enzyme activity conditions for B. amyloliquefaciens F2 protein properties and scale-up to provide a basic technology for alternative protein industrial applications.
Conclusion
In conclusion, we successfully isolated and characterized a total of twenty-two Bacillus spp. from several Jeotgal samples that are considered to have significant biotechnological potential. isolated Bacillus spp. from flounder, squid, and anchovy Jeotgal were evaluated for their roles in fermentation. These Bacillus isolates were functionally identified to have effective protease, lipase, and cellulase activities. These can be employed as agents to enhance biomass degradation necessary to produce bioactive compounds and improve food processing. The antibacterial properties of isolated Bacillus strains have been recognized as potential antimicrobial agents that can control the growth of microbial pathogens, thereby ensuring the safety of food during fermentation. In particular, F2, S2, S4, and A1, which do not produce biogenic amines, were confirmed to be safe for commercial fermentation applications. Additionally, this study demonstrates that using Bacillus spp. in fermentation improves protein extraction yields from C. pyrenoidosa and indicates the effectiveness of microbial fermentation with Bacillus strains in improving protein extraction from microalgae. Notably, B. amyloliquefaciens F2, isolated in this study, exhibited a C. pyrenoidosa protein extraction yield of 34.45 ± 1.21%, highlighting its potential to enhance conventional microalgal protein extraction methods. Finally, this study supports the commercial viability of using Bacillus spp. to extract proteins from marine biomass. Future studies should optimize the growth conditions and enzyme production, particularly for B. amyloliquefaciens F2. Furthermore, it is essential to explore the scaling up of protein extraction from microalgal species, such as C. pyrenoidosa, for commercial applications
Supplemental Materials
Supplementary data for this paper are available on-line only at http://jmb.or.kr.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Gao K Mc Kinley KR 1994 Use of macroalgae for marine biomass production and CO 2 remediation: a review J. Appl. Phycol.6456010.1007/BF 02185904 · doi ↗
- 2Larsen R Eilertsen K-E Elvevoll EO 2011 Health benefits of marine foods and ingredients Biotechnol. Adv.2950851810.1016/j.biotechadv.2011.05.01721683131 · doi ↗ · pubmed ↗
- 3TokuşogluÖ Üunal M 2003 Biomass nutrient profiles of three microalgae: Spirulina platensis, Chlorella vulgaris, and Isochrisis galbana J. Food Sci.681144114810.1111/j.1365-2621.2003.tb 09615.x · doi ↗
- 4Lee SY Show PL Ling TC Chang JS 2017 Single-step disruption and protein recovery from Chlorella vulgaris using ultrasonication and ionic liquid buffer aqueous solutions as extractive solvents Biochem. Eng. J.124263510.1016/j.bej.2017.04.009 · doi ↗
- 5Hildebrand G Poojary MM O'Donnell C Lund MN Garcia-Vaquero M Tiwari BK 2020 Ultrasound-assisted processing of Chlorella vulgaris for enhanced protein extraction J. Appl. Phycol.321709171810.1007/s 10811-020-02105-4 · doi ↗
- 6Kulkarni S Nikolov Z 2018 Process for selective extraction of pigments and functional proteins from Chlorella vulgaris Algal Res.3518519310.1016/j.algal.2018.08.024 · doi ↗
- 7Katsimichas A Karveli I Dimopoulos G Giannakourou M Taoukis P 2023 Kinetics of high pressure homogenization assisted protein extraction from Chlorella pyrenoidosa Innov. Food Sci. Emerg. Technol.8810343810.1016/j.ifset.2023.103438 · doi ↗
- 8Grossmann L Ebert S Hinrichs J Weiss J 2018 Effect of precipitation, lyophilization, and organic solvent extraction on preparation of protein-rich powders from the microalgae Chlorella protothecoides Algal Res.2926627610.1016/j.algal.2017.11.019 · doi ↗