Oomycete Diversity and Ecology in Declining Alder Stands in Switzerland
Goda Mizeriene, Vaidotas Lygis, Simone Prospero

TL;DR
This study explores oomycete diversity in alder tree stands in Switzerland, finding that soil is a key reservoir for these microorganisms, including potential pathogens.
Contribution
The study provides new insights into oomycete species distribution and their ecological roles in declining alder stands.
Findings
Soil samples had the highest oomycete isolation rate (82.5%) compared to water and bark.
Phytophthora was the most abundant genus, with 273 isolates identified.
Soil was found to host the most diverse oomycete community, including 16 species restricted to this substrate.
Abstract
In this study, we assessed the occurrence and diversity of four oomycete genera (Phytophthora, Phytopythium, Pythium, and Globisporangium) in 13 declining alder (Alnus glutinosa and A. incana) stands in Switzerland. For this, we sampled and analyzed soil from tree rhizosphere, water from streams and rivers along which the stands were located, and symptomatic alder bark. The overall isolation rate was 47.2%, with a total of 400 oomycete isolates recovered at all 13 sites. The highest incidence of oomycete isolates was in soil samples (baiting, 82.5% isolation rate), followed by water (baiting, 14.7%), and bark (direct isolation, 2.7%). Of all recovered oomycete isolates, 90.3% could be successfully assigned to a known species, for a total of 23 species identified, including both preferential saprotrophs and pathogens. Among all genera, Phytophthora was the most abundant with 273 isolates…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
 Figure 1
Figure 1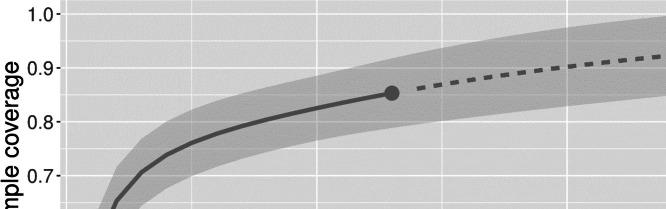 Figure 2
Figure 2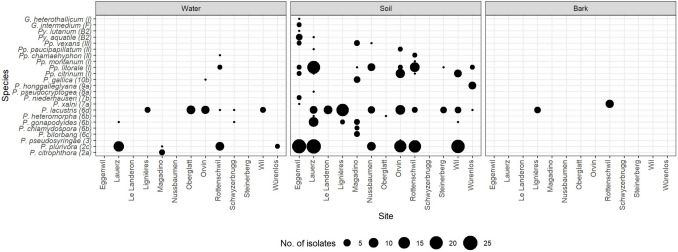 Figure 3
Figure 3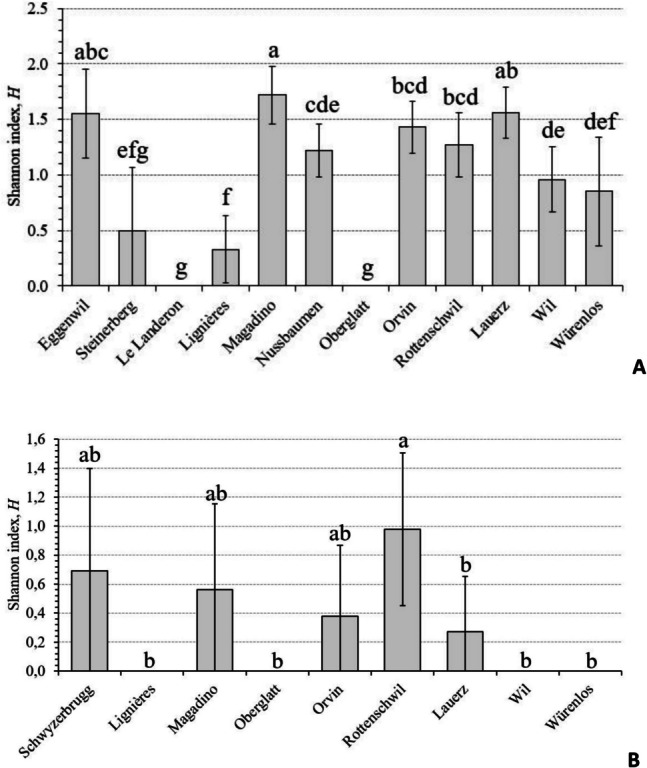 Figure 4
Figure 4- —Sciex-NMSch Fellowship
- —WSL - Swiss Federal Institute for Forest, Snow and Landscape Research
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsPlant Pathogens and Resistance · Plant Pathogens and Fungal Diseases · Yeasts and Rust Fungi Studies
Introduction
Oomycetes are filamentous, fungus-like microbial eukaryotes [1]. Thanks to their diverse lifestyles, pathogenicity, and host range, these microorganisms can be considered one of the most successful groups of eukaryotes, which is globally distributed in almost all ecosystems on Earth [2]. Although oomycetes are functionally heterogeneous [3], many of them are severe pathogens of agricultural crops and forest trees [4, 5], with a significant impact on the global food security [6]. Regarding tree health, the genera Phytophthora (P.), Pythium (Py.), and Phytopythium (Pp.) are particularly relevant.
The genus Phytophthora includes more than 200 currently known species [7], many of them being plant pathogens [8] and representing a phytosanitary threat to forest ecosystems worldwide [9]. Pythium species are not only plant but also animal, algal, or fungal pathogens, as well as saprophytes. Currently, this genus counts more than 140 species, occurring in a wide range of both aquatic and terrestrial habitats [10]. Over the years, the genus Pythium has been reclassified, and more recently, based on the phylogeny and the morphology of the sporangium, it has been subdivided into the four genera Ovatisporangium, Globisporangium, Elongisporangium, and Pilasporangium [11]. The genus Phytopythium is relatively new and was previously assigned to Pythium sp. clade K [12]. Uzuhashi et al. [11] reclassified this genus as Ovatisporangium, and now Ovatisporangium and Phytopythium are considered synonyms [13]. This genus consists of 26 species [14] found in various habitats [15], mainly as necrotrophic generalists [1].
The introduction rate of pathogenic oomycetes into Europe’s natural ecosystems has increased dramatically over the last few decades [16] and continues to rise. Since the early 1990 s, a severe decline of alder (Alnus spp.) has been observed first in Britain and then across the entire continent [17]. The causal agent of this decline was found to be Phytophthora alni sensu lato, a previously unknown Phytophthora species [18]. Later, it was shown that P. alni s. l. consists of three species, namely the hybrid P. × alni and its two parental species P. × multiformis and P. uniformis [19]. These three species differ in morphology, genetic background, virulence, and geographic distribution [20].
In Switzerland, alder dieback has been observed since the early 2000 s, and P. alni s. l. was first officially isolated in 2008 from declining alders along the Reuss river [21]. In 2020, the isolates were re-analyzed and definitively attributed to P. × alni [22]. However, since then, no comprehensive study has been carried out to investigate the diversity of Phytophthora species in declining alder stands, although black (Alnus glutinosa) and grey (A. incana) alders are ecologically important species in riparian ecosystems such as river and stream banks, and lake shores. Recently, Schoebel et al. [23] presented results of a study that investigated Phytophthora community composition in Swiss watercourses over the period 2012–2016. A total of 11 Phytophthora species were detected, but no conclusions could be drawn linking the Phytophthora species found to declining alders.
In the present study, we aimed at answering the following two questions: (i) What is the incidence and diversity of the oomycete species belonging to the genera Phytophthora, Pythium, Phytopythium (Ovatisporangium), and Globisporangium in declining alder stands in Switzerland? (ii) How do incidence and diversity of the isolated oomycetes vary across sampling sites and substrates (i.e., symptomatic bark, rhizosphere soil, and water)?
Materials and Methods
Study Sites
Field sampling was conducted at 13 sites in Switzerland between April and September 2015 (Table 1). The sites were selected with the help of the local forest service at locations (196–840 m a.s.l.) where declining alders (Alnus incana, A. glutinosa) had been observed. They either consisted of mostly disturbed (management, cattle, hiking) stands along a watercourse (stream or river) or within a marshland (for examples, see Fig. 1A and B). All sites were sampled only once, except for Rottenschwil and Orvin, which were sampled twice in different months (July and September) because of the presence of numerous declining trees. At each site, alder trees with symptoms of decline, such as bark lesions (bleeding cankers, tarry spots on the outer bark) along the stem or at the root collar, and crown thinning (Fig. 1C–E, [18]), were selected for sampling. Table 1. Information about the 13 sites sampled in this study and the number of samples collectedSiteCoordinates (WGS84)Waterbody name (type)Sampled Alnus speciesForest typeSamples (N)LongitudeLatitudeBarkSoilWaterEggenwil8.338447.36432Reuss (river)A. incanaRiparian770Lauerz8.5802547.03799Chlausenbach (stream)A. glutinosa, A. incanaRiparian881Le Landeron7.0500147.0499Vieille Thielle (duct)A. glutinosaRiparian111Lignières7.0780747.09019Unnamed (pond)A. glutinosaRiparian441Magadino8.869446.15581Ticino (river)A. incanaMarsh661Nussbaumen8.8223747.61628Nussbommersee (lake)A. incanaMarsh220Oberglatt8.5132347.48062Glatt (river)A. incanaMarsh221Orvin7.2024647.15602L’Orvine (river)A. glutinosa, A. incanaRiparian11112Rottenschwil8.37347.32264Reuss (river)A. glutinosa, A. incanaRiparian10102Schwyzerbrugg8.712347.15264Biber (river)A. incanaRiparian001Steinerberg8.5778947.04523Goldbach (stream)A. incanaRiparian221Wil8.5137747.61742Schwarzbach (stream)A. glutinosa, A. incanaRiparian221Würenlos8.3702247.43536Limmat (river)A. glutinosaRiparian111Total565613Fig. 1Typologies of sampling sites and disease symptoms. A Riparian stand along a river; B alder stand along a stream in a marsh; C old bleeding lesions on a stem of grey alder (Alnus incana); D fresh bleeding lesion (tarry spot) on a stem of black alder (Alnus glutinosa); E grey alders showing crown dieback symptoms (sparse crowns, small leaves) (A:© Swiss Forest Protection, WSL; B-E: © Phytopathology, WSL)
A total of 56 alder trees (42 A. incana and 14 A. glutinosa) were sampled (bark lesions and rhizosphere), ranging from 1 (Le Landeron and Würenlos) to 11 (Orvin) trees per site (Table 1). In Schwyzerbrugg, no trees were sampled because it was too difficult to access them. Thirty-six of the sampled alder trees (64.3%) had bleeding bark lesions on the main stem or at the root collar (11 A. glutinosa and 25 A. incana trees), 11 trees exhibited a thin crown with decline symptoms (one A. glutinosa and ten A. incana trees), and nine trees showed both crown decline and bleeding bark lesions (two A. glutinosa and seven A. incana trees). A total of 13 water samples were collected at 11 sites (two sites, Orvin and Rottenschwil, with two water samples).
Oomycete Sampling and Isolation
After removing the bark at the margin of a stem lesion with a sterile knife, from one single lesion 20–30 pieces of phloem tissue were taken using a sterilized Jamshidi needle (2 mm in diameter) and placed on a selective CARP+ medium [24] directly in the field. The Petri plates were then incubated in the laboratory at 22 °C in the dark and for 2 to 14 days examined for the presence of oomycete-like hyphae growing out of the samples. When such hyphae were observed, an agar plug from the margin of the colony was transferred with a sterile toothpick onto a new Petri plate containing carrot piece agar (CPA) medium [25] and incubated at 22 °C in the dark to obtain a pure culture.
Soil sampling was performed following the protocol of Tedersoo et al. [26]. Briefly, soil (in total about 1 kg per tree) was sampled along the four cardinal points around a symptomatic tree at 0.5–1.0 m distance from the root collar and at a depth of 5–20 cm using a spade. In the laboratory, soil was sieved through a sieve (mesh size 2 mm) and kept cool until further processing. If a waterbody (river, lake, or pond) with banks covered with A. glutinosa or A. incana trees was present at the sampling site, approximately 1.5 l of water with sediments has also been collected from that waterbody.
The presence of oomycetes in the collected soil and water samples was assessed using the baiting method proposed by Werres et al. [27]. Briefly, 3 days after sampling at the latest, 200 ml of the sampled soil was put in a plastic container (15 cm × 15 cm × 4 cm) and covered with 400 ml of distilled water. Thereafter, four young, healthy rhododendron leaves were put on the water surface as baits. Similarly, 400 ml of water with sediments has been poured into a plastic container and baited with four young, healthy rhododendron leaves. The containers were then incubated for 16-h light period at 20 °C and 8-h dark period at 15 °C for maximum 10 days. For the isolation of oomycetes, five tissue samples (ca 3 mm × 3 mm in size) were cut from each symptomatic leaf (surface-sterilized) around the formed necrotic spots and placed on Petri plates with CPA medium. The plates were incubated at 22 °C in the dark and pure cultures were obtained as described above for phloem tissue samples [27].
DNA Extraction
To obtain pure mycelium for better DNA extraction, an agar plug removed from the margin of a growing pure culture was transferred to a new Petri plate containing liquid V8 medium [28]. The plates were then incubated for 5–7 days at 22 °C in the dark until they were ca. 80% covered by the culture. The mycelium was harvested on a filter paper by filtration with a Büchner funnel and a vacuum pump. The collected mycelium was washed off the filter paper with sterile distilled water and placed in a freezer at − 20 °C until the DNA extraction. After lyophilization, DNA was extracted using the DNeasy® Plant Mini kit or the DNeasy® 96 Plant Kit (Qiagen, 96 Hilden, Germany).
PCR and Sequencing
Isolates were identified to species by sequencing part of the ribosomal ITS region. PCR amplification was conducted in a 20-µl reaction volume containing final concentrations of 2 × Master mix (JumpStart™ RedTaq® Ready Mix (Sigma-Aldrich, Germany)), 0.625 µM of each ITS6 [29] and ITS4 [30] primers, and 1 µl template DNA. The target region was amplified by PCR using a Veriti™ Thermal Cycler (Applied Biosystems, Foster City, CA, USA). Amplification was performed with initial denaturation at 95 °C for 2 min, followed by 35 cycles of denaturation: 95 °C for 30 s, annealing: 55 °C for 30 s, extension: 72 °C for 2 min, and final extension: 72 °C for 10 min. The PCR products were purified using Illustra™ ExoProStar™ PCR and Sequence Reaction Clean-Up Kit (Sigma-Aldrich, Germany), according to the manufacturer’s instructions. For sequencing, BigDye™ Terminator v. 3.1 Cycle Sequencing Kit and BigDye™ x-Terminator Purification Kit (Applied Biosystems, Carlsbad, CA) were used according to manufacturer’s protocol. The sequencing was performed on an ABI 3130 or ABI 3730xl capillary sequencer (Applied Biosystems, USA). The obtained sequences were assembled and edited using the software CLC Main Workbench version 8.0 Beta 4 (Qiagen, Bioinformatics, Denmark). For species identification, sequences (~ 800 bp) were compared with publicly available sequences in the National Center for Biotechnology Information (NCBI; https://blast.ncbi.nlm.nih.gov/Blast.cgi) database with BLAST algorithm. Two sequences were considered to belong to the same species if they showed at least 99% similarity. Representative sequences of all identified species are available in the NCBI database (https://www.ncbi.nlm.nih.gov/) under the accession numbers PV082471 to PV082524 (for details, see Table S1).
Assessment of Oomycete Diversity
Individual-based rarefaction of Hill numbers (D) [31] was used to estimate oomycete diversity across various sites and substrates. The diversity indicators are characterized by the order q, which determines the sensitivity of the index to rare or abundant species. The diversity order q = 0 (^0^D) represents species richness and shows the diversity of all species. The diversity order of q = 1 (^1^D) displays “typical” species and functions as Shannon Diversity. Finally, the diversity order of q = 2 (^2^D) represents Simpson diversity and shows the diversity of dominant species [32]. Interpolated/extrapolated curves were produced with the iNEXT package (v.3.0.0) [33] in RStudio v. 4.2.3 (RStudio PBC, Boston, USA). To investigate whether uneven sample sizes from different sites affect total oomycete species diversity, we produced interpolated/extrapolated curves using iNEXT based on sample coverage (hereinafter SC) data. The iNEXT package estimateD function was used to calculate the diversity estimates for the minimum sample size between sites, by excluding uninformative sites. To determine whether diversity differences between sites were statistically significant, the 95% confidence intervals were compared. Since visually comparing all confidence intervals was difficult, the significance of the differences in species diversity in water and soil among sites was also determined by calculating and comparing Shannon diversity indices between sites using Hutcheson’s t-test [34].
Results
Oomycete Incidence
A total of 400 oomycete (Phytophthora, Pythium, Phytopythium, and Globisporangium) isolates were recovered from the three different substrates (bark, soil, water). The overall isolation rate was 47.2%, with oomycetes recovered at all 13 sites. The highest incidence of oomycetes was obtained from soil samples (330 isolates, 82.5% isolation rate), followed by water samples (59 isolates, 14.7% isolation rate), and bark samples (11 isolates, 2.7% isolation rate). Out of the 400 oomycete isolates, 361 (90.3%) could be successfully assigned to a known species, for a total of 23 identified species (Table 2). The remaining 39 isolates could not be unequivocally assigned to a specific oomycete species and were thus removed from further analyses. Among all genera, Phytophthora was the most abundant with 273 isolates (75.6%), followed by Phytopythium (79 isolates), Pythium (six isolates), and Globisporangium (three isolates) (Table 2). Table 2. Incidence of identified (i.e., assigned to known species) oomycete taxa recovered from three substrate types (soil, water and bark) sampled in declining alder (Alnus glutinosa and A. incana) stands in Switzerland (for more information see Materials and Methods section)TaxaSoilWaterBarkSites^a^Isolates^b^SitesIsolatesSitesIsolatesP. citrophthora0-^c^13 (5.80)0-P. plurivora6113 (53.80)423 (44.20)0-P. pseudosyringae11 (0.48)0-0-P. bilorbang13 (1.43)0-0-P. chlamydospora12 (0.95)0-0-P. gonapodyides416 (7.60)22 (3.80)0-P. heteromorpha22 (0.95)0-0-P. lacustris959 (28.10)623 (44.25)14 (36.40)P. × alni11 (0.48)0-17 (63.60)P. niederhauseri12 (0.95)0-0-P. pseudocryptogea11 (0.48)0-0-P. honggalleglyana16 (2.86)0-0-P. gallica14 (1.90)11 (1.90)0-Genus* Phytophthora12210 (100)952 (100)211 (100)Pp. citrinum519 (25.00)0-0-Pp. litorale845 (59.21)12 (66.67)0-Pp. montanum11 (1.32)0-0-Pp. chamaehyphon12 (2.63)11 (33.33)0-Pp. paucipapillatum23 (3.95)0-0-Pp. vexans36 (7.89)0-0-Genus Phytopythium*876 (100)13 (100)0-Py. aquatile25 (83.33)0-0-Py. lutarium11 (16.67)0-0-Genus Pythium26 (100)0-0-G. heterothallicum11 (33.33)0-0-G. intermedium12 (66.67)0-0-Genus Globisporangium13 (100)0-0-Total (all genera)12295955211^a^Number of study sites, from which respective oomycete taxon has been isolated^b^Number of the recovered isolates and their frequency of occurrence within a specific genus (%, shown in brackets)^c^-, non-applicable
Oomycete Diversity Across Sampling Sites
The highest oomycete diversity was observed in Eggenwil (ten species), Lauerz (nine species), Magadino (eight species), and Orvin and Rottenschwil (seven species each) (Table 3). On the other hand, the sites Le Landeron, Schwyzerbrugg, Steinerberg, Lignières, and Oberglatt exhibited the lowest oomycete diversity with only one to two species isolated from each site (Table 3). Based on the incidence data generated from sampling units (i.e., same sample size across sites), the coverage-based rarefaction and extrapolation sampling curve indicates that the sampling effort in this study was sufficient to catch up to 85% of the expected oomycete diversity (Fig. 2). As stated above, Eggenwil had the highest observed species diversity, but based on rarefied and extrapolated estimates for 22 isolates, Magadino had the highest species richness (^0^D, 8.19), as well as the highest estimations for the most common (together with Eggenwil: ^1^D, 7.49 and 7.53, respectively) and most abundant (^2^D, 7.10) species (Table 3). Seven of the 13 sites sampled had SC scores greater than 0.90, which allows a reliable estimation of oomycete diversity (Table 3). Table 3. Observed and estimated diversity of identified (i.e., assigned to known species) oomycetes recovered from three substrate types (symptomatic bark, rhizosphere soil, and water) in declining alder (Alnus glutinosa and A. incana) stands in Switzerland (13 sampling sites; for more information see Table 1)Observed species diversityEstimated species diversity^a^Sampling siteNumber of isolatesNumber of speciesMethod^b^SC^c^Order Q^d^Diversity estimate of order q**qD (lower CL)^e^qD (upper CL)^f^Eggenwil4110R0.8107.536.039.0314.122.865.3822.631.733.54Lauerz789R0.9205.264.246.2913.723.004.4423.002.363.63Le Landeron81n.a.^g^n.an.an.an.an.aLignières272R0.9901.971.442.5011.290.941.6521.160.901.42Magadino208E0.9108.196.2510.1217.495.709.2827.105.119.08Nussbaumen204E1.0004.003.014.9913.432.764.1123.212.523.89Oberglatt92n.an.an.an.an.an.aOrvin497R0.9105.303.996.6113.943.114.7723.372.734.01Rottenschwil567R0.9305.424.426.4213.872.944.7923.112.233.98Schwyzerbrugg22n.an.an.an.an.an.aSteinerberg52n.an.an.an.an.an.aWil354R0.9703.633.014.2512.722.203.2322.271.722.82Würenlos114E1.0004.221.407.0413.621.755.4922.941.084.80^a^Calculated for a sample size of N = 22 isolates^b^R rarefaction, E extrapolation^c^Estimated sample coverage^d^Sensitivity of the index to rare or abundant species^e^The bootstrap lower confidence limits for expected richness (value of 0.95)^f^The bootstrap upper confidence limits for expected richness (value of 0.95)^g^n.a. not applicableFig. 2Oomycete sample completeness (sample coverage) based on the sampling design adopted in this study. The solid line represents sample coverage based on rarefaction, while the dashed line is based on extrapolation. The shading in grey indicates the 95% confidence intervals
Phytophthora
Phytophthora isolates were recovered at all 13 sites and belonged to 13 different species spanning seven different ITS clades (Fig. 3; [7]). On average, three Phytophthora species were detected in each site. The highest diversity was observed at Magadino with a total of six species, most of which belonged to the ITS clade 6 (Fig. 3). Twelve out of 13 detected Phytophthora species (all except P. citrophthora) were recovered from soil samples, five from water (including P. citrophthora), and two from bark lesions (P. × alni and P. lacustris).Fig. 3. Diversity of identified (i.e., assigned to known species) oomycetes recovered in Switzerland from water samples, rhizosphere soil samples, and bark lesions on symptomatic black alder (Alnus glutinosa) and grey alder (A. incana) trees. Numbers and letters in the parentheses after species names indicate the clade or group [7, 11, 13, 15] to which each species belongs. For full species names, please see Table 2
Phytopythium
The genus Phytopythium was found at nine of the 13 sites and included seven known species (Table 2, Fig. 3). Rottenschwil showed the highest Phytopythium species diversity, with a total of four different species detected. All six detected Phytopythium species were recovered from soil samples, two from water, and none from bark lesions.
Pythium and Globisporangium
Representatives of genera Pythium and Globisporangium were rarely detected at the sampled sites and were isolated exclusively from soil samples (Fig. 3).
Oomycete Diversity Across Different Substrate Types
Oomycete species diversity and incidence showed a significant variation within and among substrates (Fig. 3). Only one species—P. lacustris—was present and abundant in all substrates, while 16 species were restricted to a specific substrate, mainly soil.
The rhizosphere of symptomatic alder trees harbored the most diverse oomycete community, with a total of 22 known species (Table 2, Fig. 3). However, most of these species were rather rare and each was represented by less than ten isolates. Typical species present in the rhizosphere of declining alder trees were P. plurivora, P. lacustris, Pp. litorale, and Pp. citrinum (altogether they included 80.0% of the identified oomycete isolates recovered from soil). Based on 95% confidence intervals, significant differences were observed among some sites in oomycete diversity in the soil (Fig. S1). These differences were confirmed by a Hutcheson’s t-test comparing Shannon diversity indices (Fig. 4A).Fig. 4. Comparison of Shannon diversity indices among oomycete communities recovered from the rhizosphere soil of symptomatic alder trees (A) and water samples (B) taken in declining alder stands at 12 (soil) and 9 (water) positive sites (for more information, see Tables 1 and 3). Different letters above the bars indicate statistically significant differences at p < 0.05 based on a Hutcheson’s t-test. Error bars indicate values of confidence intervals
The aquatic oomycete community included a total of seven known species (five Phytophthora and two Phytopythium species) (Table 2, Fig. 3). The most common species in the water samples were P. lacustris and P. plurivora (23 isolates each; altogether they comprised 83.6% of the identified oomycete isolates recovered from water). Although the overall incidence of both species was the same, the first was found at six sites whereas the latter only at four sites. Notably, P. citrophthora was found only in water samples. Four Phytophthora species (P. lacustris, P. gallica, P. gonapodyides, and P. plurivora) and two Phytopythium species (Pp. chamaehyphon and Pp. litorale) were identified in both soil and water samples. The differences among sites in oomycete species diversity in water were not as marked as in the soil (Fig. S2, Fig. 4B), but the number of water samples analyzed was considerably lower than that of soil samples.
Finally, only two Phytophthora species were isolated from alder bark lesions, namely, P. × alni, the causal agent of alder decline, and P. lacustris (Fig. 3).
Discussion
Oomycete Abundance and Diversity
Oomycetes were found at all 13 sites sampled, with Phytophthora being the most abundant genus followed by Phytopythium. The other two investigated genera, Pythium and Globisporangium, were clearly less frequent. Among the three substrates analyzed, soil yielded the highest number of isolates and species, highlighting once again its importance as a reservoir for these microorganisms. Previous surveys of terrestrial and aquatic oomycete communities occasionally showed higher species diversity in water bodies compared to tree rhizosphere or forest soils (e.g., [35–37]). According to Català et al. [35], water bodies, and rivers in particular, seem to concentrate the inoculum of oomycetes of large areas, especially after rainy periods when the inoculum is discharged into larger bodies of water as runoff. On the other hand, the success of oomycete recovery from one or another substrate may depend on other factors, including the detection technique applied (e.g., [38]).
We found a higher oomycete species diversity, in particular of Phytophthora species, in the rhizosphere of declining alder stands compared to other studies carried out in Austria [39], Italy [40], Poland [41], and Turkey [42]. Bregant et al. [43] isolated a similar number (12) of Phytophthora species from the rhizosphere of declining A. glutinosa trees in Portugal, but only five of them were common to our study. Riit et al. [44] used a metagenomic approach for detection of oomycetes in the rhizosphere soil of declining alder trees across the Fennoscandian and Baltic countries and detected DNA of ten Phytophthora species and 28 Pythium species. These differences could be partially due to detection methods applied, but also to the influence of abiotic and biotic factors like sampling season, geographic location, and vegetation type (e.g., [45, 46]).
The overall diversity of Phytophthora species in our water samples was similar to that reported in surveys conducted in other European countries (e.g., [39, 47, 48]), and in Australia [49]. Of the other oomycete genera, only two species of Phytopythium were isolated at low frequencies from water samples during the present study. This is quite surprising as most taxa of the families Pythiaceae and Peronosporaceae are dependent on aquatic environments and are usually abundantly recovered from natural water bodies (e.g., [48, 50, 51]). This low success of oomycete recovering from water samples may be explained (inter alia) by the fact that we used an ex situ baiting method, while an in situ baiting would probably have allowed capturing more species [38].
Bark lesions on Alnus species yielded the lowest number and diversity of oomycetes—we were able to isolate only two Phytophthora species. A lower incidence and diversity of Phytophthora species in the alder bark than in the soil was also reported in other studies (e.g., [52]) and may be due to several factors, including a suboptimal isolation method (in our case, direct plating of bark samples in the field) or occasional sampling of old lesions in which Phytophthora might not be active anymore.
Insights into the Ecology of the Isolated Oomycete Species
The Genus Phytophthora
Phytophthora plurivora, P. lacustris, and P. gonapodyides were the most abundant species in the rhizosphere soil and water samples, all of which are widespread in natural and agricultural ecosystems. Phytophthora plurivora is an aggressive soil-borne plant pathogen with a broad host range and worldwide distribution, often associated with declining forest trees (e.g., [53]). The high incidence of P. plurivora in declining alder stands in Switzerland is in agreement with the results of previous studies conducted in Europe (e.g., [41–43, 54, 55]).
Although the clade 6 member P. lacustris is usually regarded as a saprotroph or opportunistic plant pathogen, which is common in riparian ecosystems in Europe and North America [56–58], there is a growing concern about its involvement in the etiology of tree diseases. O’Hanlon et al. [59] speculated that this species may be responsible for black alder decline in Northern Ireland. The pathogen was also found infecting this alder species in Portugal [60], as well as causing diseases in several other plant hosts in Europe [57]. In our study, P. lacustris, together with P. × alni, was the only oomycete species isolated from alder bark lesions. Given the predominance of P. lacustris in Swiss watercourses [23], its exact role in causing alder decline in Switzerland should be further investigated.
The third most frequently isolated Phytophthora species in our study, P. gonapodyides, also belongs to ITS clade 6 and has been traditionally regarded as a weak parasite with saprophytic abilities, usually present in aquatic environments [61]. In Europe, this species was already associated with declining broadleaved trees [53]. However, although in North America P. gonapodyides is considered an important species involved in the etiology of native alder species dieback [56], its role in alder decline in Europe is still unclear.
All the other ten Phytophthora species identified in our study were restricted to one or two sites and mostly present only in soil. Among them, seven species were already known to occur in Switzerland [23], whereas for P. heteromorpha, P. niederhauseri, and P. pseudocryptogea, this is the first report for the country. Phytophthora heteromorpha was first described in Italy from riparian habitats and in inoculation experiments proved to be pathogenic on A. incana [62]. Phytophthora niederhauseri is a highly pathogenic polyphagous species associated with ornamentals, fruit trees, and native plants, distributed worldwide, including Europe [63, 64]. Finally, P. pseudocryptogea is a species within the P. cryptogea species complex that was officially described in 2015 [65]. In Turkey, P. pseudocryptogea was recovered from the rhizosphere of declining oaks [66], whereas in Canada, the species was reported to cause root rot on western white pine (Pinus monticola) in seed orchards [67]. In Italy, it was isolated from declining alder trees [68], which indicates its potential to be pathogenic on trees.
The Genera Phytopythium, Pythium, and Globisporangium
The rhizosphere soil of declining alders was found to host a diverse assemblage of species of the genera Phytopythium, Pythium, and Globisporangium, which, to our knowledge, had never been previously investigated in natural ecosystems in Switzerland. These genera are known to include numerous plant pathogens mainly of tree seedlings and herbaceous plants [11, 69]. In this study, two Phytopythium species, namely Pp. litorale and Pp. citrinum, were quite commonly isolated from rhizosphere soil of declining alders*.* Derviş et al. [70] speculated that Pp. litorale may be the causal agent of the severe decline of oriental plane in Turkey, while Polat et al. [71] associated it to a kiwifruit dieback. In several European countries, the USA, and Vietnam, Pp. litorale was found in watercourses [37, 39, 51, 59]. Phytopythium citrinum is known as a common inhabitant of aquatic and riparian ecosystems in Europe and North America [47, 51]. Also, it was isolated from the rhizosphere of declining black alder and pedunculate oak trees in Poland [55, 72]. In a recent study by Christova [47], both Pp. citrinum and Pp. litorale showed moderate to high potential to infect several woody plant species, as well as some perennial and herbaceous plants. For this reason, both organisms were determined as pathogens with a wide host range. The other five Phytopythium species detected in this study were less frequent and limited to a few sites. While Pp. vexans is widespread worldwide and shows pathogenicity towards economically important woody hosts to which it causes root rot, damping off, crown rot, stem rot, or patch canker (e.g., [64], and references therein), the published information on the ecology of Pp. montanum, Pp. chamaehyphon, and Pp. paucipapillatum is rather scarce and does not refer to Alnus species. Similarly, neither of the two Pythium species found in the present study, Py. aquatile and Py. lutarium, and neither of the two Globisporangium species, G. heterothallicum and G. intermedium, were associated with alder previously.
Conclusions
Confirming previous studies in Europe and North America, our analyses revealed a diverse and abundant community of oomycetes in declining alder stands in Switzerland, in particular in the rhizosphere of symptomatic trees. The species recovered ranged from known saprotrophs or opportunistic plant pathogens to aggressive pathogens. Noteworthy, only two Phytophthora species were isolated from bark lesions on alders, namely, P. × alni, the known causal agent of alder decline, and P. lacustris. Although this might be partially explained by the fact that not all sampled bark lesions were still active, it could also suggest that the observed alder decline might be due to pathogens acting only in the root system and/or abiotic stress factors. Future studies are needed to build up understanding of the ecological role of all oomycete species recovered in such ecosystems as well as their possible interactions with alder and a changing environment. Understanding how oomycete communities are assembled in stands (e.g. based on functional traits, [73]) with different health status would also help to further clarify their role in forest ecosystems.
Supplementary Information
Below is the link to the electronic supplementary material.Supplementary file1 Table S1 Oomycete isolates recovered from declining alder (Alnus glutinosa and A. incana) stands in Switzerland with their respective substrate of origin and sequence GenBank accession numbers. Fig. S1 Oomycete diversity in the rhizosphere soil of symptomatic alder trees at 12 different sites sampled during the present study (for more information see Tables 1 and 3) in Switzerland. The comparison is shown by individual abundance-based rarefaction (solid lines) and extrapolation curves (dashed lines). Each panel is based on the first three Hill numbers [32, 33]: ^0^D (left panel), ^1^D (middle panel) and ^2^D (right panel). The color-shaded regions show the 95% confidence intervals. Fig. S2 Oomycete diversity in water samples taken in the declining alder stands at 9 different sites in Switzerland (for more information see Tables 1 and 3). The comparison is shown by individual abundance-based rarefaction (solid lines) and extrapolation curves (dashed lines). Each panel is based on the first three Hill numbers [32, 33]: ^0^D (left panel), ^1^D (middle panel) and ^2^D (right panel). The color-shaded regions show the 95% confidence intervals. (DOCX 195 KB)
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Marçais B (2018) Phytophthora alni species complex (alder Phytophthora). Wallingford, CABI Compendium 40948. 10.1079/cabicompendium.40948
- 2Tkaczyk M, Szmidla H, Sikora K (2023) Occurrence of Phytophthora species in the rhizosphere of dying black alder Alnus glutinosa in central and northeastern Poland. Sylwan 167:452−460. 10.26202/sylwan.2023040