Complete defect in PA-PLA1α secretion function leading to autosomal recessive woolly hair and hypotrichosis: insights from a novel compound heterozygous LIPH variant study in a Chinese pedigree
Xinyue Zhang, Kexin Guo, Jiawei Liu, Xueting Yang, Rui Zhang, Rongrong Wang, Donglai Ma, Xue Zhang

TL;DR
A Chinese patient with woolly hair and hypotrichosis was found to have a novel LIPH gene variant that disrupts protein secretion, causing the disease.
Contribution
A novel compound heterozygous LIPH variant was identified and shown to cause complete loss of PA-PLA1α secretion function.
Findings
The novel LIPH variant c.530T>G nearly abolishes PA-PLA1α secretion.
Arginine substitutions in LIPH variants are linked to impaired protein secretion.
The study confirms an autosomal recessive inheritance pattern for the disease.
Abstract
Autosomal recessive woolly hair/hypotrichosis (ARWH) is a rare inherited hair disease. In this study, we report a 31-year-old Chinese female with the characteristic clinical features of woolly hair and hypotrichosis. Through whole-exome sequencing (WES), we identified a novel missense variant (NM_139248.3: c.530T>G: p.Leu177Arg) and a previously reported missense variant (c.742C>A: p.His248Asn) of LIPH in the patient. TA cloning demonstrated that these variants were located on different alleles, supporting an autosomal recessive inheritance pattern. In silico tools predicted the novel variant to be disease-causing, likely reducing the stability of PA-PLA1α, the protein encoded by LIPH. PA-PLA1α, a member of the AB hydrolase superfamily and the lipase family, functions as a secreted protein to perform its hydrolytic and catalytic activities. Through a secretion assay, we observed that…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
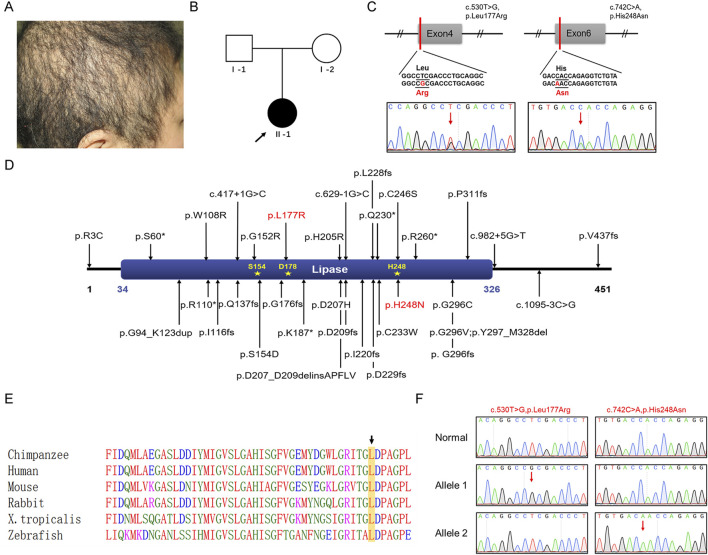 FIGURE 1
FIGURE 1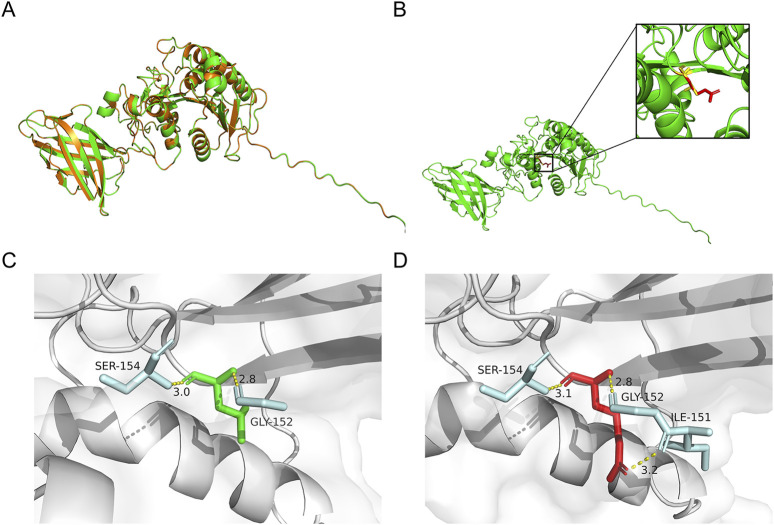 FIGURE 2
FIGURE 2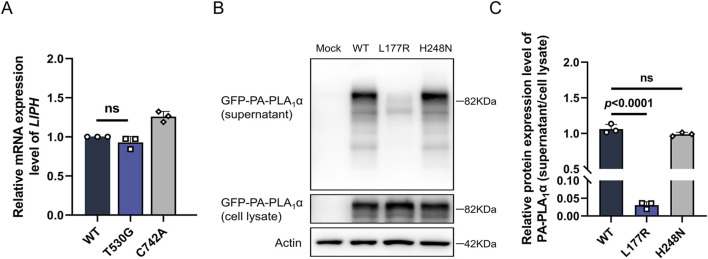 FIGURE 3
FIGURE 3| Patients | Years |
| Amino acid changes | LIPH domain affected | Phenotypes | References |
|---|---|---|---|---|---|---|
| 1 | 2014 | c.742C>A | p.His248Asn | Lipase domain | Hypotrichosis | Liu LH et al. |
| 2 | 2017 | c.614A>G | p.His205Arg | Lipase domain | Woolly hair, hypotrichosis | Chang XD et al. |
| c.742C>A | p.His248Asn | |||||
| 3 | 2017 | c.736T>A | p.Cys246Ser | Lipase domain | Woolly hair, hypotrichosis | |
| c.742C>A | p.His248Asn | |||||
| 4 | 2017 | c.736T>A | p.Cys246Ser | Lipase domain | Woolly hair, hypotrichosis | |
| c.742C>A | p.His248Asn | |||||
| 5 | 2017 | c.454T>A | p.Gly152Arg | Lipase domain | Woolly hair, hypotrichosis | |
| c.742C>A | p.His248Asn | |||||
| 6 | 2020 | c.686delAins18 | p.Asp229fs37* | Lipase domain | Woolly hair, hypotrichosis | Lv H et al. |
| c.736T>A | p.Cys246Ser | |||||
| 7 | 2022 | c.736T>A | p.Cys246Ser | Lipase domain | Woolly hair, hypotrichosis | Qu B et al. |
| c.742C>A | p.His248Asn | |||||
| 8 | 2025 | c.530T>G | p.Leu177Arg | Lipase domain | Woolly hair, hypotrichosis | This study |
| c.742C>A | p.His248Asn |
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsHair Growth and Disorders · Skin and Cellular Biology Research · Wnt/β-catenin signaling in development and cancer
1 Introduction
Autosomal recessive woolly hair/hypotrichosis (ARWH) is a nonsyndromic hereditary hair disease first defined by Hutchinson in 1974 (Hutchinson et al., 1974). Disease onset typically occurs soon after birth and is clinically characterized by coarse, sparse, dry, and tightly curled hair, which may also involve beard and axillary hair (Kazantseva et al., 2006). Compared with other family members, patients usually have blonde or lighter hair (Horev et al., 2009). ARWH exhibits significant genetic heterogeneity, with causative variants identified in several genes. Based on these genes, ARWH is classified into three types: ARWH1 (OMIM #278150) caused by variants in LPAR6, ARWH2 (OMIM #604379) caused by variants in LIPH, and ARWH3 (OMIM #616760) caused by variants in KRT25. Additionally, homozygous nonsense variants in C3orf52, which interacts with LIPH, have also been detected in individuals with ARWH (Kazantseva et al., 2006; Pasternack et al., 2008; Shimomura et al., 2008; Ansar et al., 2015; Malki et al., 2020). Understanding the specific roles of these genes and the consequences of their variants is essential for elucidating the pathology of ARWH.
Among the causative genes of ARWH, the LIPH gene has received particular attention. Situated on chromosome 3q27-q28, LIPH spans approximately 45 kb and contains 10 exons (Jin et al., 2002). It encodes the enzyme lipase member H (phosphatidic acid-preferring phospholipase A1α, PA-PLA_1_α). This secreted enzyme works extracellularly, hydrolyzing phosphatidic acid (PA) to produce lysophosphatidic acid (LPA), which is an essential molecule for the growth of hair follicles (Horev et al., 2009). Consequently, the functional disruption of PA-PLA_1_α has been recognized as a key mechanism in ARWH2 pathogenesis. This disruption leads to deficient LPA production, which severely impairs normal hair development and results in the characteristic ARWH phenotype. Existing studies on pathogenic LIPH variants have predominantly focused on those impairing the hydrolytic activity of PA-PLA_1_α, whereas cases involving secretion defects are comparatively rare and remain less explored as a pathogenic mechanism. This leaves a gap in our understanding of ARWH pathogenesis, particularly secretion defects, which requires further study.
Here, we report a Chinese patient with clinical symptoms of woolly hair and hypotrichosis since birth. In this patient, we identified a novel variant (NM_139248.3: c.530T>G, p.Leu177Arg) and a reported variant (c.742C>A, p.His248Asn) in LIPH. Through genetic and functional analyses, we confirmed that this novel variant disrupts the development of hair follicles by completely abolishing PA-PLA_1_α secretion function, ultimately resulting in ARWH. Notably, this study found that arginine substitutions may lead to defective PA-PLA_1_α secretion, providing evidence for a previously unrecognized pathogenic mechanism. Finally, we systematically reviewed reported pathogenic variants of ARWH and their corresponding phenotypes across populations, revealing that LIPH variants are the predominant cause of ARWH in Chinese patients. This highlights the importance of prioritizing LIPH gene variants in the clinical diagnosis of Chinese patients with ARWH-like symptoms.
2 Methods
2.1 Clinical data collection and ethical compliance
This study enrolled patients who presented with distinct phenotypes of short, thin, and curly hair. The patient provided written informed consent prior to participation for the collection of her case details, images, and peripheral blood samples. The study received approval (Approval No. 2022170) from Peking Union Medical College’s Institutional Review Board.
2.2 Whole-exome sequencing (WES) and Sanger sequencing
Genomic DNA was extracted from the patient’s peripheral blood. The sequencing library was prepared. Target regions were sequenced with paired-end reads of 150 bp and 100× raw read coverage. The human reference genome (GRCh37/hg19) was used to align the produced raw reads. The variants were annotated with ANNOVAR and screened through public databases, including the dbSNP database, the 1000 Genomes Project, and gnomAD. Candidate pathogenic variants identified via WES were subsequently verified using Sanger sequencing. Primers were designed by Primer-BLAST online software. The results of the sequencing were examined by SnapGene software and then further validated.
2.3 Bioinformatics analysis
REVEL, CADD, and Mutation Assessor were used to predict pathogenicity. Evolutionary conservation analysis was performed using T-Coffee for multiple sequence alignments. Functionally important domains and sites within proteins were identified by referring to the InterPro database. The change in the stability of the variant protein sequence or structure was predicted using MUpro and iStable. The effect of the variant on protein structure was assessed using HOPE. AlphaFold was employed to construct a three-dimensional (3D) protein model, and PyMOL was used to visualize the protein structures of both the wild-type (WT) and variant.
2.4 RNA extraction, cDNA synthesis, and TA cloning construction
Using the conventional RNA extraction method, the RNA was isolated from the patient’s peripheral blood. Subsequently, the RNA was reverse-transcribed into cDNA according to the protocol. The sequence of the LIPH variants (c.530T>G and c.742C>A) was amplified from the above cDNA using the primer pair LIPH-RTPCR-F: 5′- CAGATGTTGGCAGAAGGAGC -3′ and LIPH-RTPCR-R: 5′- GCTGACACACTGCCATC -3′. Following the normal procedure, RT-PCR products were cloned into the pMD18-T vector. Sequencing was performed on 50 clones chosen at random.
2.5 Cell culture, plasmid construction, and transfection
HEK293T cells were cultured in complete Dulbecco’s Modified Eagle Medium (DMEM) at 37°C under a 5% CO_2_ atmosphere. WT, as well as the c.530T>G and c.742C>A variants of LIPH, were linked into pEGFP-N vectors. HEK293T cells were transfected with Lipofectamine 3000 (Invitrogen, United States).
2.6 Quantitative reverse transcription-PCR (qPCR)
RNAs from HEK293T cells transfected with pEGFP-N vectors were reverse transcribed into cDNAs. qPCRs were performed using SYBR Green^®^ Master Mix (Yeasen, China). The primers for LIPH and the internal control ACTB used for qPCR were LIPH-qPCR-F: 5′- GTGCTTGTCAAGATCAGACGC-3′ and LIPH-qPCR-R: 5′- CGCAGGTCAGGTTTTTCCTTG-3′; ACTB-qPCR-F: 5′-CATGTACGTTGCTATCCAGGC-3′; ACTB-qPCR-R: 5′-CTCCTTAATGTCACGCACGAT-3′. Data were shown as the mean ± SD.
2.7 Preparation of supernatants and cell lysates
After transfection with pEGFP-N vectors for 24 h, the complete medium for HEK293T cells was replaced with serum-free medium. Following a 24-h incubation, the media were harvested and precipitated using methanol and chloroform. The precipitated protein pellet was redissolved in 40 μL of 1× SDS sample buffer with protease inhibitors (Roche, Switzerland) and phosphatase inhibitors (Roche, Switzerland) and boiled for 10 min. The remaining HEK293T cells were washed with DPBS. Then, 150 μL of 1× SDS sample buffer, as mentioned above, was added directly to the cell pellet and boiled for 10 min.
2.8 Western blotting
Western blotting was performed using a conventional method. Protein extracts from the supernatants and cell lysates were separated by SDS-PAGE. The primary antibodies used for probing the proteins were anti-β-Actin antibody (Cell Signaling Technology, 3700, United States) and anti-LIPH antibody (Proteintech, 16602-1-AP, China). The results were detected using chemiluminescence with species-specific HRP-conjugated secondary antibodies.
3 Results
3.1 Clinical features
The patient was a 31-year-old Chinese female who presented with short, thin, curly, and coarse hair that had been prone to breakage since birth (Figure 1A). She exhibited no abnormalities in intelligence or overall health and had no significant medical or family history of skin or limb deformities. Her parents were not consanguineous (Figure 1B) and declined further genetic testing.
Clinical characteristics of a Chinese patient with ARWH and identification and analysis of the LIPH variants. (A) Clinical symptoms of the patient with short, thin, curly, and coarse hair. (B) Family pedigree: The black circle indicates the affected individual, and the arrow indicates the proband. (C) Sanger sequencing confirmed the c.530T>G and c.742C>A variants in LIPH in the patient. The two variant bases are marked by red arrows. (D) The positions of all reported pathogenic variants associated with ARWH are shown. The LIPH protein contained one crucial domain, the lipase (34–326) domain, and three active sites, Ser154, Asp178, and His248. Red denotes variants found in this work. (E) Leu177 was evolutionarily conserved across different species. Leucine is marked with a black arrow and framed in a yellow box. (F) TA cloning and sequencing indicated that the variants occurred in trans in the patient. The two variant bases are marked by red arrows.
3.2 Genomic sequencing and in silico analysis
WES was performed to identify pathogenic variants associated with the patient’s symptoms. Ultimately, two missense variants were identified in LIPH (NM_139248.3): c.530T>G (p.Leu177Arg) and c.742C>A (p.His248Asn). These variants were verified by Sanger sequencing (Figure 1C). The p.His248Asn variant was a known pathogenic variant associated with woolly hair/hypotrichosis. The p.Leu177Arg variant, on the other hand, was novel and had not been found in the dbSNP database, HGMD, or in-house databases (all reported pathogenic variants were shown in Figure 1D). Referring to the InterPro database, this novel variant residue is situated in a crucial domain known as lipase (IPR013818), very close to the active site (Asp178) (Figure 1D). This evidence suggests it has the potential to affect protein function by modifying the area surrounding the site. The leucine residue at amino acid position 177 is highly conserved across species (Figure 1E). The variant was predicted to be pathogenic by in silico analysis tools (REVEL, 0.983 damaging; Mutation Taster, 1 disease-causing; CADD, 29.7 pathogenic). The classification of the variant was “likely pathogenic” (PM1+PM2_supporting + PP3_strong) according to the ACMG/AMP 2015 guideline. In addition, no variants associated with woolly hair or hypotrichosis were found.
3.3 Located variants by TA cloning
The unavailability of peripheral blood samples from the patient’s parents precluded us from determining the origin of these variants. To verify whether each of the two variants was inherited separately from the father and the mother, TA cloning was employed. PCR was used to amplify an LIPH gene fragment encompassing the c.530T>G and c.742C>A variant sites from the patient’s genomic DNA. The PCR products were then cloned into the pMD18-T vector, and a total of 50 clones were randomly selected for subsequent Sanger sequencing. As shown in Figure 1F, the two variants occurred in trans (i.e., on different alleles), confirming that the patient had woolly hair and hypotrichosis with an autosomal recessive inheritance pattern.
3.4 Structural bioinformatics analysis of the variant impact
The possible impact of the p.Leu177Arg variant on the secondary and tertiary structures of PA-PLA_1_α was further investigated. The p.Leu177Arg variant was predicted to decrease the stability of the PA-PLA_1_α based on results from MUpro and iStable software. According to the HOPE database, the p.Leu177Arg variant led to an increase in amino acid size, a shift in charge from neutral to positive, and a loss of hydrophobic interactions within the core. These alterations may affect intramolecular interactions and subsequently influence the function of PA-PLA_1_α.
The 3D structures of the WT and variant proteins were predicted to evaluate the influence of the novel p.Leu177Arg variant on the structure of the PA-PLA_1_α, based on an AlphaFold model (AF-Q8WWY8-F1-v4). The structures were visualized using PyMOL (Figure 2A). The substitution regions were highlighted in the protein model (Figure 2B). In the WT, Leu177 formed hydrogen bonds with Gly152 and Ser154 (Figure 2C). In the variant, the initial hydrogen bond distance between Leu177 and Ser154 increased from 3.0 Å to 3.1 Å, while an additional hydrogen bond formed between Leu177 and Ile151 (Figure 2D). The above-mentioned alterations likely reduce the stability and impact the intermolecular interactions of the protein.
Predicted effects of variant on protein structure. (A) The superimposed view of the PA-PLA1α in its wild-type (WT) and p.Leu177Arg variant. (B) Predicted structures depicted the changes in PA-PLA1α after the Leu177Arg. Red and yellow structures separately indicate WT and the p.Leu177Arg variant. (C,D) The changes in hydrogen bonds between pre- and post-variant amino acids. Amino acids at position 177 of the WT and variant are shown in green and red, respectively. The amino acids involved in the interaction are highlighted in blue. The hydrogen bond is represented by the yellow dotted line.
3.5 Severe secretion defect of PA-PLA1α due to novel LIPH variant in ARWH
PA-PLA_1_α functions as a secreted protein to perform its hydrolytic and catalytic activities. By transfecting the corresponding variants into HEK293T cells, we observed that the novel missense variant p.Leu177Arg did not affect mRNA expression or intracellular protein synthesis and expression (Figures 3A,B). However, the secretion assay revealed that the novel missense variant almost abolished the secretion of PA-PLA_1_α compared to the control (p < 0.0001), which would have resulted in a complete impairment of the hydrolysis function (Figures 3B,C). The p.His248Asn variant has been shown to cause a complete loss of PA-PLA_1_α hydrolytic activity, despite not affecting PA-PLA_1_α secretion (Shinkuma et al., 2010). In this study, we found that the novel variant directly led to a severe secretion defect of PA-PLA_1_α, resulting in impaired hydrolytic activity. Therefore, the novel compound heterozygous LIPH variants led to defective PA-PLA_1_α function, which is the underlying cause of the disease.
Functional analysis of the LIPH variants. (A) The relative mRNA expression level of LIPH in HEK293T cells transfected with the corresponding constructs was measured by qPCR. (B,C) Expression and secretion of WT, Leu177Arg, and His248Asn variant PA-PLA1α transfected in HEK293T cells were investigated using Western blotting. The top panel shows the expression levels of PA-PLA1α obtained from the constructs in the supernatant; the second panel demonstrates the expression levels of PA-PLA1α in the cell lysate. β-Actin expression was used for normalization.
4 Discussion
A typical presentation of ARWH is marked by the occurrence of rough, dry, and curly spiral hair at birth (Akiyama, 2021; Hayashi and Shimomura, 2022). Hair growth is generally slow and sparse in affected individuals, with hairs that tend to break easily, resembling sheep’s wool, and varying degrees of sparsity, rarely exceeding a few inches in length (Shimomura, 2012). The majority of ARWH patients experience moderate-to-severe hypotrichosis, which varies among the individuals and families (Shimomura et al., 2009a; Takeichi et al., 2017). The worst situations result in the loss of all scalp hair (Shimomura et al., 2009b). Histopathological examination of the scalp reveals abnormal hair follicles and shafts. A few patients also exhibit thinning or lack of eyebrows, eyelashes, and beards, as well as armpit and body hair (Shimomura et al., 2009a).
According to Online Mendelian Inheritance in Man (OMIM), ARWH caused by LIPH variants is classified as ARWH2. To date, 34 pathogenic variants of LIPH associated with ARWH2 have been reported. The majority of ARWH2 families reside in Japan, Pakistan, and the Volga-Ural area of Russia, showing clear ethnic and geographic differences and a considerably high prevalence of their respective founder variants (Kazantseva et al., 2006; Shimomura et al., 2009a; Tanahashi et al., 2014; Hayashi and Shimomura, 2022). In the Japanese population specifically, the founder variants p.Cys246Ser or p.His248Asn have been reported in all ARWH2 families (Akiyama, 2021). Interestingly, according to data provided by HGMD, at least one of these two founder variants was consistently identified in all Chinese ARWH2 patients in this and previous studies (Table 1), providing factual evidence to support the hypothesis of genetic similarity between ARWH in Chinese and Japanese populations.
PA-PLA_1_α, predominantly expressed in hair follicles, the anterior cortex, the cuticle of the hair shaft, and the Huxley layer of the inner root sheath (IRS), exerts its hydrolytic and catalytic activities as a secreted protein. Recent genetic studies on human hair disorders have demonstrated the essential role of LIPH-LPA-LPAR6 signaling in hair shaft development (Minakawa et al., 2024). It is proposed that the 2-acyl-LPA that PA-PLA_1_α generates in the IRS activates LPAR6 through either paracrine or autocrine mechanisms (Inoue et al., 2011). This activation induces the release of membrane-bound pro-TGFα through TACE-dependent mechanisms. Soluble TGFα then binds to epidermal growth factor receptor (EGFR) on IRS cells, activating EGFR and promoting IRS development for proper hair shaft formation. The p.His248Asn variant has been reported to cause a complete loss of PA-PLA_1_α hydrolytic activity and failure to activate LPAR6, despite not affecting PA-PLA_1_α secretion (Shinkuma et al., 2010). In this study, we found that the novel p.Leu177Arg variant directly led to a severe secretion defect of PA-PLA_1_α, resulting in impaired hydrolytic activity. Notably, such a severe secretion defect has only been observed in the p.Gly152Arg and p.His205Arg variants, with this the third instance (Chang et al., 2017). These findings suggest that variants resulting in the substitution of normal amino acids with arginine (Arg) may specifically impair PA-PLA_1_α secretion, as observed in p.Leu177Arg, p.Gly152Arg, and p.His205Arg. In contrast, other missense variants located near these three arginine-substitution sites, such as p.Cys246Ser, p.His248Asn, and even p.Ser154Ala—which is only one residue away from p.Gly152Arg—do not impact PA-PLA_1_α secretion (Shinkuma et al., 2010; Chang et al., 2017).
Arginine is particularly noteworthy due to its unique combination of physico-chemical properties, which distinguish it from most other amino acids. First, the side chain of arginine contains a guanidinium group, which endows it with a strong positive charge, making it the most positively charged residue among the 20 amino acids. This highly charged nature is unparalleled by other amino acids and allows arginine to participate in extensive ionic interactions and hydrogen bonding. However, in the context of protein folding, such strong electrostatic interactions can disrupt normal protein folding by affecting the stability of local structural elements (Harms et al., 2011; Perdriau et al., 2025). Second, arginine has one of the largest side chains among the 20 amino acids, and its bulkiness often introduces significant steric hindrance. This can lead to structural distortions, particularly when arginine is substituted in tightly packed regions (Gupta and Uversky, 2024). When investigating the impact of the p.Leu177Arg, p.His205Arg, and p.Gly152Arg variants, it becomes evident that these unique properties of arginine play a significant role. After arginine substitutions (p.Gly152Arg and p.His205Arg variants), the substituted arginine interacted with more nearby residues by hydrogen bonds (Supplementary Figures S1A–D), whereas other missense variants such as p.Ser154Ala and p.His248Asn tended to reduce interactions with other residues (Supplementary Figures S1E–H). Furthermore, arginine substitutions significantly increased residue size due to its bulky side chain. An increased number of interacting residues may work together with the bigger residue itself to form a larger group that disrupts the native conformation of the protein, causing steric hindrance and a loss of flexibility required for proper folding, ultimately resulting in endoplasmic reticulum (ER) retention and impaired secretion. The consistent observation across multiple variants suggests a potential mechanistic link between arginine substitutions and secretion defects. This observation provides important insights into the molecular mechanisms underlying protein secretion defects. Further research is required to confirm these findings.
Additionally, in this study, we conducted a literature review and comprehensively compiled the reported pathogenic variants of ARWH along with their corresponding phenotypes and populations (Supplementary Table S1). Our findings indicated that LIPH variants are the predominant cause of ARWH in Chinese patients, with no significant differences in phenotypic severity observed across different variants. In contrast, only one case has been associated with a KRT25 variant, and no reported cases have been linked to LPAR6 variants. Therefore, when clinically diagnosing Chinese patients who present with woolly hair and hypotrichosis, particular attention should be given to genetic screening for potential LIPH variants.
Despite the promising results of this study, several limitations should be acknowledged. First, while genetic and functional evidence strongly supports the pathogenicity of the LIPH variant, several predicted loss-of-function variants have been observed in public databases, such as GnomAD. However, in contrast to the other variants, which primarily affect the hydrolytic activity, the novel variant p.Leu177Arg identified here directly disrupts protein expression, leading to a complete loss of secretion, highlighting a less common pathogenic mechanism. Second, our finding is based on a single patient. Future investigations involving larger cohorts would be essential to validate these findings and better clarify the genotype-phenotype correlations of LIPH variants.
In conclusion, this study reported a patient with ARWH and confirmed the clinical diagnosis through in vitro experiments, demonstrating that the novel LIPH c.530T>G (p.Leu177Arg) variant affected protein function by impairing PA-PLA_1_α secretion. This direct effect on protein secretion is extremely rare in previous studies of ARWH and may be related to arginine substitutions, providing new insights into the pathogenic mechanism of ARWH. Our findings expand the spectrum of LIPH variants associated with ARWH of Chinese origin and have the potential to influence the formulation of new diagnostic and treatment approaches for this condition.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Akiyama M. (2021). Isolated autosomal recessive woolly hair/hypotrichosis: genetics, pathogenesis and therapies. J. Eur. Acad. Dermatology Venereol. JEADV 35 (9), 1788–1796. 10.1111/jdv.17350 33988877 · doi ↗ · pubmed ↗
- 2Ansar M.Raza S. I.Lee K.Irfanullah Shahi S.Acharya A. (2015). A homozygous missense variant in type I keratin KRT 25 causes autosomal recessive woolly hair. J. Med. Genet. 52 (10), 676–680. 10.1136/jmedgenet-2015-103255 26160856 PMC 5090258 · doi ↗ · pubmed ↗
- 3Chang X.-D.Gu Y.-J.Dai S.Chen X.-R.Zhang C.-L.Zhao H.-S. (2017). Novel mutations in the lipase H gene lead to secretion defects of LIPH in Chinese patients with autosomal recessive woolly hair/hypotrichosis (ARWH/HT). Mutagenesis 32 (6), 599–606. 10.1093/mutage/gex 043 29346610 · doi ↗ · pubmed ↗
- 4Gupta M. N.Uversky V. N. (2024). Biological importance of arginine: a comprehensive review of the roles in structure, disorder, and functionality of peptides and proteins. Int. J. Biol. Macromol. 257 (Pt 1), 128646. 10.1016/j.ijbiomac.2023.128646 38061507 · doi ↗ · pubmed ↗
- 5Harms M. J.Schlessman J. L.Sue G. R.García-Moreno B. (2011). Arginine residues at internal positions in a protein are always charged. Proc. Natl. Acad. Sci. U. S. A. 108 (47), 18954–18959. 10.1073/pnas.1104808108 22080604 PMC 3223443 · doi ↗ · pubmed ↗
- 6Hayashi R.Shimomura Y. (2022). Update of recent findings in genetic hair disorders. J. Dermatology 49 (1), 55–67. 10.1111/1346-8138.16204 34676598 · doi ↗ · pubmed ↗
- 7Horev L.Tosti A.Rosen I.Hershko K.Vincenzi C.Nanova K. (2009). Mutations in lipase H cause autosomal recessive hypotrichosis simplex with woolly hair. J. Am. Acad. Dermatology 61 (5), 813–818. 10.1016/j.jaad.2009.04.020 19766349 · doi ↗ · pubmed ↗
- 8Hutchinson P. E.Cairns R. J.Wells R. S. (1974). Woolly hair. Clinical and general aspects. Trans. St. John’s Hosp. Dermatological Soc. 60 (2), 160–177.4467886 · pubmed ↗