Temporal induction of the homeodomain transcription factor Nkx2-1 is sufficient to respecify foregut and hindgut endoderm to a pulmonary fate in Xenopus laevis
Brian A Hyatt, Erin Lundberg, Rachael Eye, Scott A Rankin, Aaron M Zorn

TL;DR
The study shows that the NKX2-1 transcription factor can reprogram gut cells in frog embryos to take on lung cell characteristics.
Contribution
The study demonstrates that temporal induction of NKX2-1 is sufficient to respecify endoderm to a pulmonary fate in Xenopus.
Findings
Human and Xenopus NKX2-1 induce pulmonary surfactant genes in foregut and hindgut endoderm.
NKX2-1 suppresses midgut and hindgut-specific gene expression.
A single pulmonary transcription factor can reprogram endoderm fate.
Abstract
The ability of transcription factors (TFs) to regulate cell fate decisions is paramount in developmental, homeostatic, and pathogenic contexts. The homeodomain TF NKX2-1 is an essential and evolutionarily conserved master regulator of pulmonary fate in vertebrates. In this study, we tested the spatial-temporal ability of Xenopus and Human NKX2-1 to respecify foregut and hindgut endoderm in developing Xenopus laevis embryos into a pulmonary fate, as indicated by expression of pulmonary surfactant genes sftpc and sftpb . Interestingly, we find that both Human and Xenopus NKX2-1 can induce the ectopic expression of pulmonary surfactant genes in foregut and hindgut endoderm over a wide range of developmental times, as well as suppress the expression of midgut and hindgut specific genes. These results suggest a single pulmonary TF can reprogram developing endoderm and specify pulmonary fate.…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
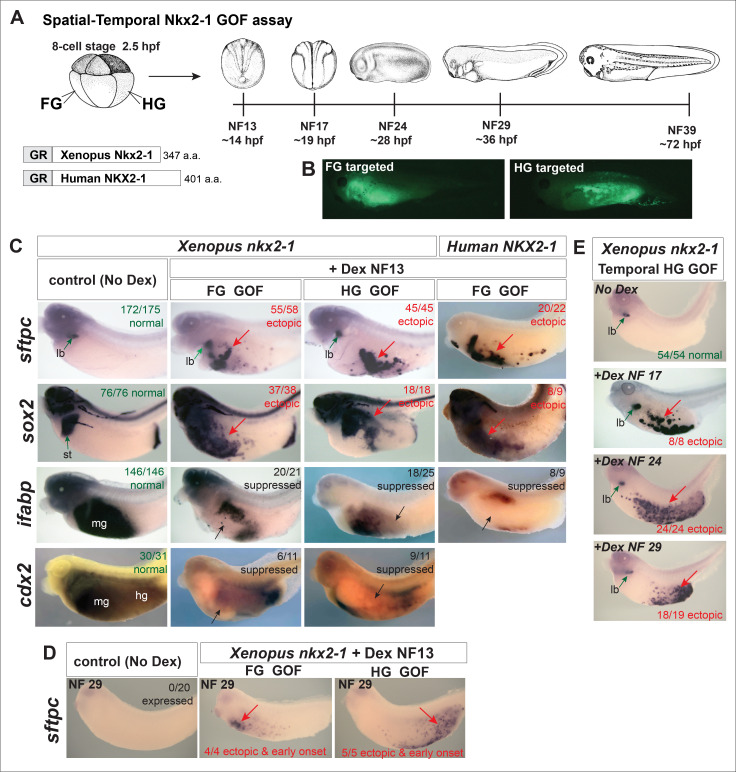 Figure 1
Figure 1Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsNeonatal Respiratory Health Research
Description
The amphibian
- Xenopus laevis* is a widely used vertebrate model organism with numerous advantages including external embryonic development, ease of experimental manipulations such as targeted microinjection, microdissections, and introduction of small biomolecules readily dissolved in culture media (Edwards and Zorn, 2021). The early Xenopus embryo has been extensively fate mapped, which permits targeted microinjection of reagents to specific progenitor populations of organs from all 3 germ layers in the embryo (Moody, 1987; Dale and Slack, 1987). While adult mammalian and amphibian lungs are morphologically very distinct, pulmonary fate induction in the developing foregut as well as early morphogenetic processes of trachea-esophageal septation and lung budding are evolutionarily conserved amongst vertebrates (Zorn and Wells, 2009; Rankin et al . , 2015; Rankin et al . , 2016; Edwards and Zorn, 2021; Rankin and Zorn 2022b).
The homeodomain TF Nkx2-1 is a master regulator of pulmonary cell fate (Kimura et al . , 1996; Minoo et al . , 1999), maintaining adult lung function (Boggaram, 2009) and transcriptionally activating lung specific genes Sftpc and Sftpb (Kelly et al . , 1996; Bohinski et al . , 1994). In addition, the expression of Nkx2-1 is used to identify lung specific tissue formed in pluripotent stem cell protocols (Longmire et al . , 2012; Rankin et al . , 2018) and classify lung cell tumor types (Camolotto et al . , 2018).
We generated temporally inducible expression constructs for both X.laevis nkx2-1 and Human NKX2-1 , fusing the ligand binding domain of the Human nuclear glucocorticoid receptor (GR) to the N-terminus of each; such GR fusions to TFs are widely used in Xenopus to control temporal activity of TFs, as nuclear translocation can be induced by exposure to the synthetic GR ligand Dexamethasone (Dex) which can be added to the embryo culture buffer at any stage of interest (Kolm and Sive, 1995; Sinner et al . , 2004; Oropeza and Horb, 2012). In these experiments, messenger RNA (mRNA) encoding the GR-TF fusion is transcribed in vitro and then microinjected into the early cleavage stage embryo where it is translated *in vivo; * the GR-TF fusion protein is sequestered in the cytoplasm until embryos are transferred into culture media containing Dex, at which point the fusion protein translocates to the nucleus and exerts activity. We note that while the full length Xenopus Nkx2-1 protein (347 amino acids in length; NCBI Reference Sequence: NP_001079093.1) is shorter than full length Human NKX2-1 protein (401 amino acids; NCBI Reference Sequence: NP_001073136.1), overall the 2 proteins are 70% identical at the amino acid level, with the DNA binding homedomains (human amino acids 194-247) being 100% identical ( extended data fig.1 ).
In order to determine if NKX2-1 has the ability to reprogram foregut (FG) and hindgut (HG) endoderm into a pulmonary fate, we employed a spatial-temporal gain of function (GOF) assay in Xenopus ( Fig.1A,B ). mRNA encoding Xenopus or Human GR-NKX2-1, along with *GFP * mRNA as a lineage trace, was microinjected at the 8-cell stage into dorsal-vegetal (targeting FG) or ventral-vegetal (targeting HG) blastomeres; examination of GFP fluorescence during subsequent development confirmed proper targeting ( Fig.1B ). Embryos were transferred into media containing Dex at a number of progressive developmental stages ( Fig.1A ), cultured to NF39, and assayed by in situ hybridization ( Fig.1C,D ). Dex induction of GR-NKX2-1 prior to gastrulation NF10.5 resulted in arrested development / failure of gastrulation. Dex induction of both Xenopus and Human NKX2-1 after gastrulation at NF 13 resulted in the ectopic expression of the pulmonary surfactant genes sftpc ( Fig.1C ) and sftpb (identical to sftpc ) in either FG or HG endoderm ( Fig.1C ). Interestingly, the foregut/esophagus/stomach TF *sox2 * was also ectopically induced, whereas the midgut endoderm marker ifabp and mid-hindgut endoderm TF cdx2 ( ** Fig. 1C ** ) were both suppressed by Xenopus Nkx2-1and Human NKX2-1. This suggests that NKX2-1 can respecify early post-gastrula foregut and hindgut endoderm into a pulmonary fate. As nkx2-1 is also endogenously expressed in pharyngeal thyroid endoderm and regulates thyroid development (Small et al . , 2000; Hollemann and Pieler 2000; Kurmann et al . , 2015; Dame et al . , 2017), we examined markers of pharyngeal endoderm, foxe1 , and thyroid endoderm, pax2 , however these were unchanged by Nkx2-1 overexpression. We also investigated the temporal onset of ectopic sftpc expression when Nkx2-1 was Dex activated at NF13; sftpc was detected at NF29 but not earlier ( Fig.1D ); this NF29 expression is earlier than the endogenous onset of sftpc which normally occurs at NF37/38 in the lung bud (Hyatt et al . , 2007; Rankin and Zorn, 2022a), and demonstrates a precocious induction in addition to the ectopic location. It is intriguing why there is a delay between induction of Nkx2-1 at NF13 and activation of sftpc (NF29), and this warrants further investigation, perhaps suggesting that alternative transcriptional programs must first be shut off prior to a sftpc pulmonary fate being induced.
Endoderm regional identities are progressively determined during Xenopus development (Horb and Slack, 2001; Shifley et al . , 2012), and while post-gastrula endoderm is known to still be plastic in potential (McLin et al . , 2007; Zhang et al . , 2013; Rankin et al . , 2018), hindgut fate is specified during gastrulation (Rankin et al . , 2018). We thus tested if Nkx2-1 was sufficient to induce pulmonary fate in progressively older endoderm ( Fig.1E ). Intriguingly, inducing Xenopus Nkx2-1 activity via Dex treatment at NF 17, NF24, and NF29 all still resulted in ectopic activation of sftpc in the HG ( Fig.1E ). Similar results were obtained inducing Nkx2-1 in the FG at these older times, suggesting Nkx2-1 has the ability to respecify HG and FG endoderm that is undergoing differentiation/commitment. Future experiments will continue to test progressively older Nkx2-1 induction times.
The ability of individual or combinations of TFs to reprogram one adult or fully differentiated somatic cell type directly into another has garnered great interest for tissue engineering applications aimed to create cell types to treat diseases or investigate mechanisms of regeneration and injury repair. Xenopus has proven a useful model organism to screen the ability of TFs to respecify / reprogram cell fates (Horb et al . , 2003; Afelik et al . , 2006; Afouda et al . , 2018; Oropeza and Horb, 2012; Grand et al . , 2023). Our results suggest a single pulmonary TF can respecify developing endoderm: Nkx2-1 can induce pulmonary specific *sftp * expression and suppress alternative fates (midgut, hindgut). Future studies may test if Nkx2-1 is sufficient to induce pulmonary fate in non-endoderm cells (mesoderm and ectoderm). Preliminary targeted injections and analyses suggest Nkx2-1 cannot induce sftpc in ectoderm cells, and thus multiple pulmonary or endoderm-specific TFs may be required to convert non-endoderm lineages into pulmonary fate. In addition, our work provides a comparative platform for future studies investigating how mutations in Human NKX2-1 may affect its transcriptional activity.
Methods
Xenopus laevis care
All experiments involving Xenopus were carried out using approved Institutional Animal Care and Use Committee (IACUC) protocols from Bethel University (160302A and R220301) or Cincinnati Children’s Hospital Medical Center (IACUC2022-0026). Xenopus laevis were purchased from Xenopus I (Dexter, MI) or Nasco (Fort Atkinson, WI). Induction of egg laying was performed via hormonal injections (Pregnant Mare Gonadotropin (PMG), 50 units 2 days prior and human Chorionic Gonadotropin (hCG), 800 units one day prior to egg laying). Eggs were fertilized with an in vitro sperm suspension and kept in a salt solution of one-third (R/3) Marc’s Modified Ringers (MMR) containing 100 ug/ml gentamycin. 8-cell stage embryos with regular cleavage patterns and clear pigment differences were placed in a 4-6% Ficoll solution in R/3 for microinjection. Embryos were staged according to Nieuwkoop and Faber (1994).
Generation of inducible expression constructs
Xenopus laevis and Human GR-NKX2-1 fusion constructs were produced by PCR amplifying the respective nkx2-1 genes with primers containing EcoRV and *SpeI * recognition sequences. Amplicons were cloned downstream and in frame with the Human GR sequence and 5’ capped mRNA for microinjection was made from linearized plasmid using mMessage mMachine T7 transcription kit (ThermoFisher AM1344).
*In situ * hybridization and photo documentation
Whole mount in situ hybridization (WM-ISH) was performed on MEMFA fixed embryos as described (Sive et al . , 2000). Anti-sense DIG labeled probes for WM-ISH were created from linearized plasmids containing the cDNA for each of the following genes: sftpc , sftpb (Hyatt et al . , 2007), sox2 (Ishii Y et al . , 1998; Rankin et al . , 2015), ifabp (Chalmers and Slack, 1998; Shi and Hayes, 1994), cdx2 (Chalmers et al . , 2000; Rankin et al . , 2018), foxe1 (El-Hodiri et al . , 2005), and pax2 (Wang et al . , 2011), using T7, T3, or SP6 polymerases and the Roche DIG RNA labeling mix (Millipore Sigma 11277073910) according to the manufacturer’s instructions. Following WM-ISH, embryos were bleached under fluorescent light in a 1% H 2 O 2 , 5% Formamide and 0.5X SSC solution followed by refixing in MEMFA. Embryos were transferred to 1X PBS in a 1% 1X PBS agarose dish for scoring and photo documentation on a Ziess Stemi 2000-C stereoscope with a mounted camera.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Afelik S Chen Y Pieler T 200661 Combined ectopic expression of Pdx 1 and Ptf 1a/p 48 results in the stable conversion of posterior endoderm into endocrine and exocrine pancreatic tissue.Genes Dev 20110890-93691441144610.1101/gad.37870616751182 PMC 1475757 · doi ↗ · pubmed ↗
- 2Afouda BA Lynch AT de Paiva Alves E Hoppler S 2017128 Genome-wide transcriptomics analysis identifies sox 7 and sox 18 as specifically regulated by gata 4 in cardiomyogenesis.Dev Biol 43410012-160610812010.1016/j.ydbio.2017.11.01729229250 PMC 5814753 · doi ↗ · pubmed ↗
- 3Boggaram V 200911 Thyroid transcription factor-1 (TTF-1/Nkx 2.1/TITF 1) gene regulation in the lung.Clin Sci (Lond)11610143-5221273510.1042/CS 2008006819037882 · doi ↗ · pubmed ↗
- 4Bohinski RJ Di Lauro R Whitsett JA 199491 The lung-specific surfactant protein B gene promoter is a target for thyroid transcription factor 1 and hepatocyte nuclear factor 3, indicating common factors for organ-specific gene expression along the foregut axis.Mol Cell Biol 1490270-73065671568110.1128/mcb.14.9.5671-5681.19948065304 PMC 359092 · doi ↗ · pubmed ↗
- 5Camolotto SA Pattabiraman S Mosbruger TL Jones A Belova VK Orstad G Streiff M Salmond L Stubben C Kaestner KH Snyder EL 20181126 Fox A 1 and Fox A 2 drive gastric differentiation and suppress squamous identity in NKX 2-1-negative lung cancer.Elife 710.7554/e Life.3857930475207 PMC 6303105 · doi ↗ · pubmed ↗
- 6Chalmers AD Slack JM 199881 Development of the gut in Xenopus laevis.Dev Dyn 21241058-838850952110.1002/(SICI)1097-0177(199808)212:4<509::AID-AJA 4>3.0.CO;2-L 9707324 · doi ↗ · pubmed ↗
- 7Chalmers AD Slack JM Beck CW 200081 Regional gene expression in the epithelia of the Xenopus tadpole gut.Mech Dev 9610925-477312512810.1016/s 0925-4773(00)00379-810940633 · doi ↗ · pubmed ↗
- 8Dale L Slack JM 198741 Fate map for the 32-cell stage of Xenopus laevis.Development 9940950-199152755110.1242/dev.99.4.5273665770 · doi ↗ · pubmed ↗