The Schizosaccharomyces pombe nucleolar protein Nsk1 modulates rDNA silencing during interphase
Jun-Song Chen, Sarah M. Hanna, Alaina H. Willet, Kathleen L. Gould

TL;DR
This study explores the role of the nucleolar protein Nsk1 in rRNA silencing in Schizosaccharomyces pombe.
Contribution
The novel finding is that Nsk1 modulates rDNA silencing during interphase, a previously unknown nucleolar function.
Findings
Nsk1 participates in rRNA silencing during interphase.
Nsk1 does not affect rDNA segregation, transcription, or nucleolar organization.
Abstract
Schizosaccharomyces pombe Nsk1 acts at kinetochores during mitosis to prevent error-prone chromosome segregation and it is phosphoregulated by Cdk1 . The Clp1 / Cdc14 protein phosphatase, to which Nsk1 binds, reverses Cdk1-mediated phosphorylation of Nsk1 during anaphase. During interphase, Nsk1 localizes exclusively to the nucleolus and its function there is unknown. In this study, we examined whether Nsk1 shares functions in the nucleolus with other known Clp1 / Cdc14 phosphatase substrates that localize there. We found that Nsk1 participates in rRNA silencing but not rDNA segregation, rDNA transcription, or nucleolar organization.
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
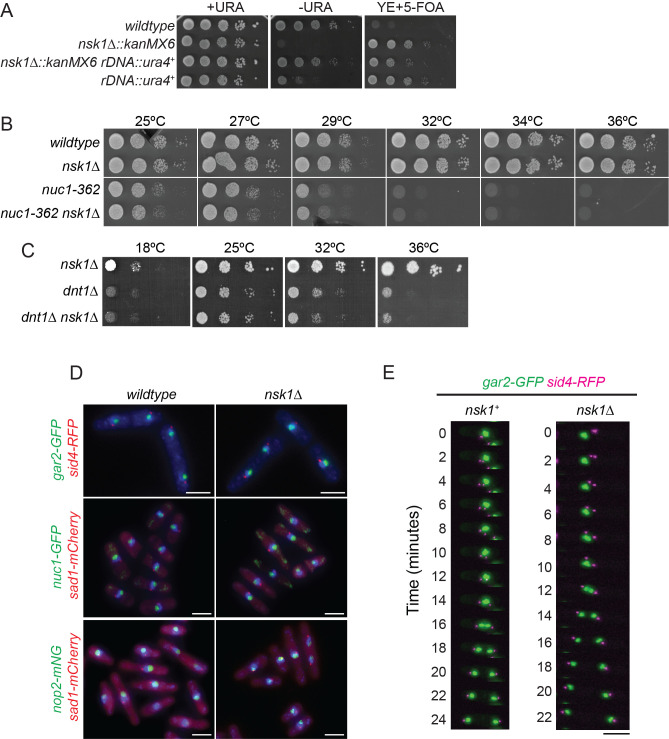 Figure 1
Figure 1|
|
|
|
|
972 |
|
Lab stock |
|
KGY8393 |
|
This study |
|
KGY4965 |
|
Jin et al., 2007 |
|
KGY7195 |
|
This study |
|
KGY8395 |
|
This study |
|
YDM2381 |
|
Dannel McCollum lab |
|
|
|
This study |
|
KGY10030-2 |
|
This study |
|
KGY965-2 |
|
This study |
|
KGY973-2 |
|
This study |
|
|
|
This study |
|
|
|
This study |
|
|
|
Hirano et al., 1989 |
|
|
|
This stud |
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsMicrotubule and mitosis dynamics · DNA Repair Mechanisms · Fungal and yeast genetics research
Description
Previously, we and others identified Schizosaccharomyces pombe Nsk1 as a substrate and binding partner of the Clp1 protein phosphatase (Buttrick et al., 2011; Chen et al., 2013; Chen et al., 2011). Although * clp1 ^+^ * is not essential, Clp1 dephosphorylates Cdk1 substrates and therefore plays a role in mitotic exit (reviewed in Clifford et al., 2008; Mocciaro and Schiebel, 2010; Stegmeier and Amon, 2004; Trautmann and McCollum, 2002). Nsk1 was found to localize to the kinetochore during mitosis where it acts to ensure accurate chromosome segregation and during interphase, Nsk1 localizes to the nucleolus (Buttrick et al., 2011; Chen et al., 2011).
Here, we examined whether Nsk1 plays similar roles in the nucleolus to other S. pombe Clp1 and Saccharomyces cerevisiae Cdc14 binding partners that reside in the nucleolus during interphase. In S. cerevisiae , Net1 is a nucleolar protein that binds and anchors Cdc14 there during interphase (Shou et al., 1999; Traverso et al., 2001; Visintin et al., 1999). S. cerevisiae Tof2 is a second S. cerevisiae Cdc14 binding partner in the nucleolus (Geil et al., 2008) and Tof2 is related to Net1 (Waples et al., 2009). A second S. pombe nucleolar protein that binds Clp1 is Dnt1 (Jin et al., 2007). S. pombe Dnt1 is related to both S. cerevisiae Net1 and Tof2 (Jin et al., 2007).
We first tested whether rDNA silencing was perturbed in nsk1∆ cells because Dnt1 , Net1, and Tof2 are all involved in transcriptional silencing of rDNA (Geil et al., 2008; Huang et al., 2008; Jin et al., 2007; Straight et al., 1999). For this, we monitored for de-repression of the * ura4 ^+^ * reporter gene that had been integrated into the normally silenced rDNA repeats (Jin et al., 2007; Thon and Verhein-Hansen, 2000). Depression of the * ura4 ^+^ * reporter in this context leads to enhanced growth on plates lacking exogenous uracil and increased sensitivity to 5-fluoroorotic acid (5-FOA). By this assay, we found that rDNA silencing was partially relieved in nsk1∆ cells ( Figure 1A ) and to the same extent as in *dnt1∆ * cells (Jin et al., 2007). We next asked whether nsk1∆ interacted genetically with nuc1-632 , a mutant of the largest subunit of DNA-directed RNA polymerase 1, that is critical for the function and structural integrity of the nucleolus (Hirano et al., 1989). Although dnt1∆ showed a negative genetic interaction with nuc1-632 cells (Jin et al., 2007), we found no evidence of genetic interaction between nsk1∆ and nuc1-632 ( Figure 1B ). nsk1∆ also did not have any genetic interaction with dnt1∆ ( Figure 1C ).
We next examined whether other nucleolar proteins localized normally in nsk1∆ cells because in the absence of Dnt1 or S. cerevisiae Net1, several nucleolar proteins are delocalized implicating Dnt1 and Net1 in nucleolar organization (Jin et al., 2007; Straight et al., 1999). Gar2-GFP, a nucleolar marker and ortholog of human nucleolin (Gulli et al., 1995; Rutherford et al., 2024), localized normally within the nucleolus of nsk1∆ cells, as did two other nucleolar proteins, Nop2 , a rRNA methyltransferase involved in ribosome biogenesis (Rutherford et al., 2024), and Nuc1 (Hirano et al., 1989) ( Figure 1D ).
Tof2 is required for proper segregation of the rDNA during anaphase in S. cerevisiae (Geil et al., 2008). In S. pombe , the nucleolar DNA segregates after the bulk of the genomic DNA (Granot and Snyder, 1991; Strunnikov, 2005). Defects in rDNA structure lead to lagging rDNA during anaphase and asymmetries in rDNA segregation, easily visualized by monitoring Gar2-GFP (Win et al., 2005). Time-lapse imaging of 21 * nsk1 ^+^ * and 42 nsk1∆ cells showed that Gar2-GFP segregated equally and without lagging in all cells ( Figure 1E ).
We conclude from the combination of these experimental results that Nsk1 has no significant role in rDNA transcription, segregation of rDNA repeats during mitosis, or nucleolar organization but it does contribute to rDNA silencing during interphase. We speculate that this role may be related to its ill-defined function at centromeres during mitosis (Chen et al., 2011).
Methods
S. pombe strains used in this study were grown in yeast extract (YE) or Edinburgh minimal media (EMM) plus selective supplements (Forsburg and Rhind, 2006). Strain construction was accomplished through tetrad analysis using standard methods (Moreno et al., 1991). Tagged strains were generated by endogenously tagging the 3′ end of open reading frames with sequences encoding *mNG:hphMX6, mCherry:natMX6, RFP:kanMX6, * or
- GFP:kanMX6 * using pFA6 cassettes, as previously described (Bahler et al., 1998) and lithium acetate transformations (Keeney and Boeke, 1994). G418 (100 mg/mL; Sigma-Aldrich), Hygromycin B (50 mg/mL; Thermo Fisher), or nourseothiricin (100 µg/ml; Sigma-Aldrich) was used for selection of
- kanMX6, hphMX6, * or *natMX6 * cells, respectively. Tagged strains were confirmed by whole-cell PCR. All fusion proteins were expressed from their native promoters at their chromosomal loci.
For rDNA silencing assays, cells were grown to log phase in YE at 32°C, a 10-fold serial dilution series starting with 50,000 cells were spotted on MAL (minimum media supplemented with 225 mg/L adenine and 75 mg/L leucine), YE, and YE containing 1.5 g/L of 5-Fluroorotic acid (5-FOA; Toronto Research Chemicals, North York, Ontario, Canada) agar plates.
Ethanol fixation and DAPI staining were performed as previously described (Roberts-Galbraith et al., 2009). Fixed-cell images in Figure 1D were acquired with a Zeiss Axio Observer inverted epifluorescence microscope with Zeiss 63X oil (1.46 NA) and captured using Zeiss ZEN 3.0 (Blue edition) software and Axiocam 503 monochrome camera (Zeiss). Exposure times ranged from 100 to 250 ms.
Live-cell time-lapse imaging in Figure 1E was performed on a Leica Thunder Imager system including a DMi8 inverted microscope, a 63X plan apo oil objective (1.40 NA), a Leica K8 sCMOS camera, standard excitation and emission filters and an LED light source. Images were captured using Leica Application Suite X (LAS X) software. A CellASIC ONIX microfluidics perfusion system (Millipore Sigma) was used, and cells were loaded into Y04C plates for 10 s at 8 psi. YE liquid medium flowed through the chamber at 5 psi throughout imaging. Z-series optical sections were taken at 0.5 µm spacing and images were acquired every 2 min.
Reagents
** *S. pombe * strains used in this study **
**: **
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Bahler J, Wu JQ, Longtine MS, Shah NG, Mc Kenzie A, Steever AB, et al., Pringle JR. 1998. Heterologous modules for efficient and versatile PCR-based gene targeting in Schizosaccharomyces pombe. Yeast. 14: 943-51. 1579.10.1002/(SICI)1097-0061(199807)14:10<943::AID-YEA 292>3.0.CO;2-Y 9717240 · doi ↗ · pubmed ↗
- 2Buttrick GJ Meadows JC Lancaster TC Vanoosthuyse V Shepperd LA Hoe KL Kim DU Park HO Hardwick KG Millar JB 2011930 Nsk 1 ensures accurate chromosome segregation by promoting association of kinetochores to spindle poles during anaphase B.Mol Biol Cell 22231059-15244486450210.1091/mbc.E 11-07-060821965289 PMC 3226469 · doi ↗ · pubmed ↗
- 3Chen JS Broadus MR Mc Lean JR Feoktistova A Ren L Gould KL 201317 Comprehensive proteomics analysis reveals new substrates and regulators of the fission yeast clp 1/cdc 14 phosphatase.Mol Cell Proteomics 1251535-94761074108610.1074/mcp.M 112.02592423297348 PMC 3650322 · doi ↗ · pubmed ↗
- 4Chen JS Lu LX Ohi MD Creamer KM English C Partridge JF Ohi R Gould KL 2011117 Cdk 1 phosphorylation of the kinetochore protein Nsk 1 prevents error-prone chromosome segregation.J Cell Biol 19540021-952558359310.1083/jcb.20110507422065639 PMC 3257533 · doi ↗ · pubmed ↗
- 5Clifford DM Chen CT Roberts RH Feoktistova A Wolfe BA Chen JS Mc Collum D Gould KL 200861 The role of Cdc 14 phosphatases in the control of cell division.Biochem Soc Trans 36Pt 30300-512743643810.1042/BST 036043618481975 · doi ↗ · pubmed ↗
- 6Forsburg SL Rhind N 200621 Basic methods for fission yeast.Yeast 2330749-503X 17318310.1002/yea.134716498704 PMC 5074380 · doi ↗ · pubmed ↗
- 7Geil C Schwab M Seufert W 200878 A nucleolus-localized activator of Cdc 14 phosphatase supports r DNA segregation in yeast mitosis.Curr Biol 18130960-98221001100510.1016/j.cub.2008.06.02518595708 · doi ↗ · pubmed ↗
- 8Granot D Snyder M 1991 Segregation of the nucleolus during mitosis in budding and fission yeast.Cell Motil Cytoskeleton 2010886-1544475410.1002/cm.9702001061661641 · doi ↗ · pubmed ↗