Oral Treatment of Lactoferrin Nanocapsules Modulates the Immune Response of Mice to a Cryptosporidiosis Infection
Ibrahim Aly, Waleed E. Elawamy, Hanan T. Hamza, Hany M. El-Wahsh, Ahmed EL-Bahiry, Amira Matter, Lamia I. Bakr

TL;DR
This study shows that lactoferrin nanocapsules, when combined with nanazoxid, can reduce the effects of a mouse infection caused by Cryptosporidium by modulating the immune response.
Contribution
The novel contribution is the evaluation of lactoferrin nanocapsules combined with nanazoxid as an effective oral treatment for cryptosporidiosis in mice.
Findings
Treatment with NZ plus LF-NC significantly reduced intestinal oocyst counts and pathological lesions.
Cytokines TGF-β, INF-γ, and IL-10 levels were decreased following therapy, indicating immune modulation.
Fecal and intestinal oocyst counts were notably reduced with NZ or NZ+LF-NC treatment.
Abstract
The current study was to evaluate the immunomodulatory impact of either lactoferrin (LF), lactoferrin nanocapsule (LF-NC), or lactoferrin conjugation with nanazoxid (NZ). Moreover, identifying drugs can effectively eliminate and successfully treat the infection with Cryptosporidium. Sixty male Swiss albino mice were divided into six groups, each of 10 mice. G1 served as the control, and G2 was inoculated with 104 oocysts. Mice were orally administered NZ (200 mg/kg b.wt.), LF (150 µg/kg b.wt.), or LF-NC (150 mg/kg b.wt./day) for a period of six days post-infection. The immunomodulatory potential and drug efficacy were assessed by fecal and intestinal oocyst counts, measuring antioxidant activities and cytokine levels in addition. Intestinal tissues were examined histopathologically. The cytokines TGF-β, INF-γ, and IL-10 were increased during infection and decreased following therapy.…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
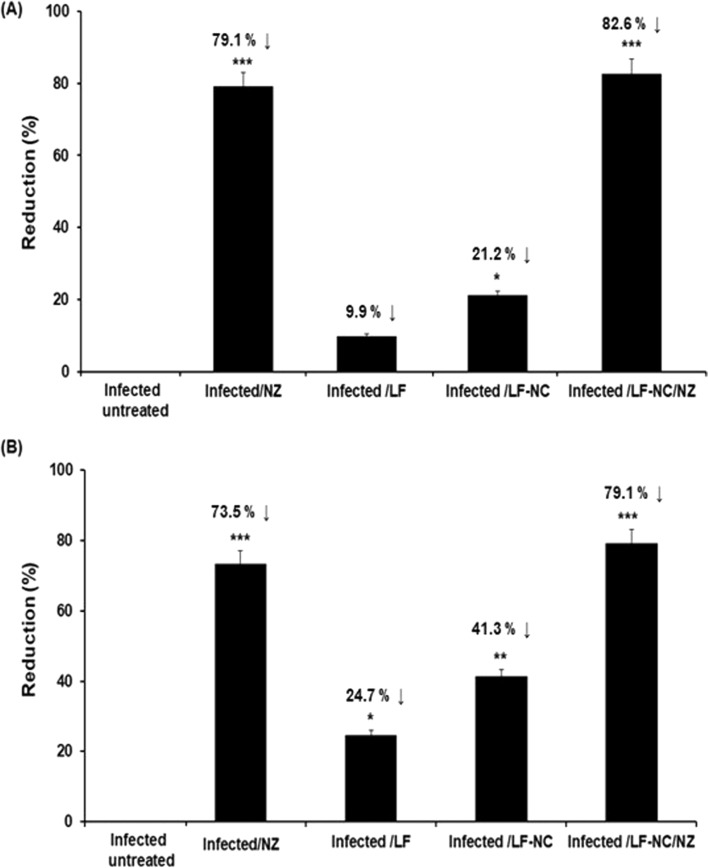 Figure 1
Figure 1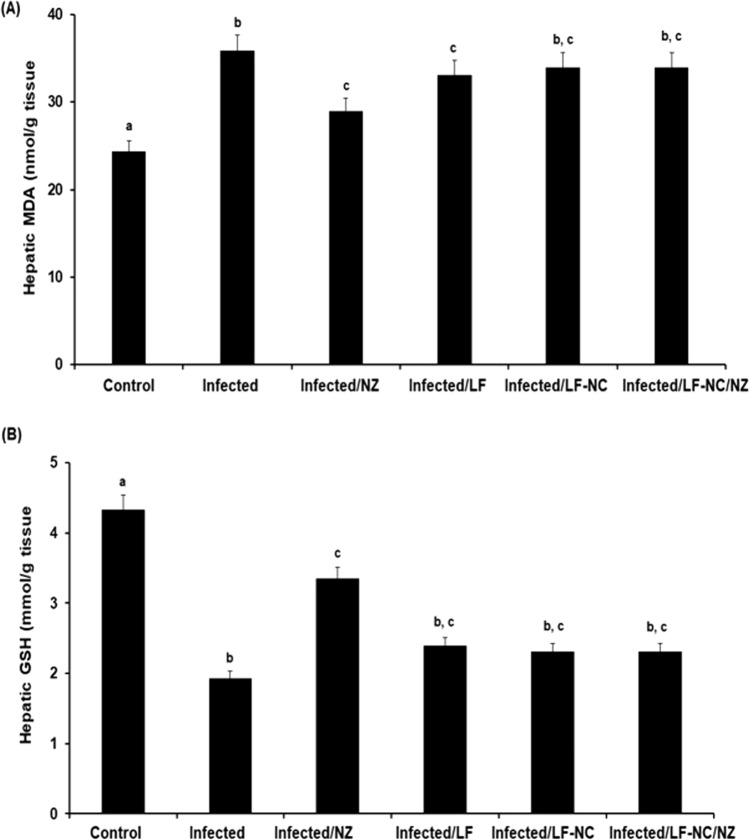 Figure 2
Figure 2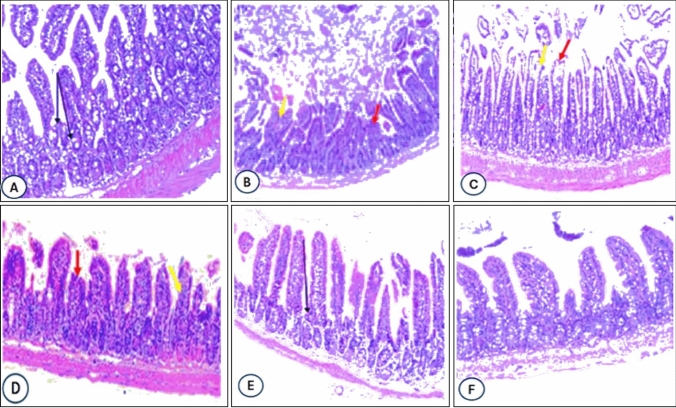 Figure 3
Figure 3- —Tanta University
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsParasitic Infections and Diagnostics · Amoebic Infections and Treatments · Parasites and Host Interactions
Introduction
Cryptosporidium is an important protozoan human and animal pathogen that is both waterborne and foodborne [8]. Cryptosporidium is a parasitic infection for the small intestine's epithelium that causes diarrhea and severe illness in immunocompetent and immunocompromised patients [18]. As well, infection with Cryptosporidium parvumis common in cattle, buffaloes, goats, sheep, horses, cats, human beings, and other vertebrates [24]. According to recent studies, Cryptosporidium spp. causes between 48,000 and 202,000 deaths and 7.6 million infections among young children in low-resource settings [20]. The oocysts of Cryptosporidium invade the apical intestinal epithelium, resulting in the development of new oocysts expelled by the host [20]. Newborns are susceptible to infection because their immune systems are immature and can become infected by ingesting small amounts of parasite oocysts. Cryptosporidium hominis and C. parvum species are infrequently detected in people; person-to-person transmission is most frequently identified in humans [16].
Until now, there has been no effective vaccine to prevent the disease, and treatment options are limited. Over 200 compounds have been assessed in humans and animals for their anti-cryptosporidial acts; no vaccine or even appropriate drug is yet to be present [21]. Nitazoxanide (NZ)is the only Food and Drug Administration-approved treatment for cryptosporidiosis [10]. NZ prevent parasitic anaerobic metabolism, which is crucial for energy production, which relieves clearance of parasitic infection [26].
The identification of natural compounds with antiparasitic activity has been a pivotal aim of alternative therapies against several parasites and other diseases [29–31]. Lactoferrin (LF) is considered an attractive natural compound for its protective properties against a wide range of microbes, and it was found to inhibit the growth of cancerous tumors [27, 32]. LF inhibits the growth of protozoan parasites, such as Toxoplasma gondii, Plasmodium falciparum, and Trypanosoma cruzi [32]. Nanoparticles (NPs) attract more attention in applications for drug delivery and treatment [6]. Nanoparticles were industrialized into smart systems for controlled drug delivery and enhanced bioavailability, enhancing the pharmacokinetic profile of encapsulated therapeutic agents [13, 33]. The present study was conducted to evaluate the immunomodulatory effect of LF or LF-NC or their combination with NZ drugs against cryptosporidiosis in an immunosuppressed mouse model.
Material and Methods
Chemicals and Drugs
Nanazoxid was purchased from Medizen Pharmaceutical Industries for Utopia Pharmaceuticals; it was given at 200 mg/kg/day successively for seven successive days. Dexamethasone was purchased from Dexazone, Al Kahira pharmaceutical and chemical industry company. Carboxymethyl sephadex-C50 chromatography was purchased from FPLC (Bio-RAD, USA). Cytokines, including IL-10, INF-γ, and TGF-β ELISA kits, were purchased from QIAGEN Company. Biochemical kits were purchased from Bio-diagnostics Company (Giza, Egypt).
Collection of C. parvum oocysts
Patients attending outpatient clinics at Theodor Bilharz Research Institute (TBRI) Hospital. The accepted experimental group to collect the oocysts of C. parvum, they were with diarrhea or gastrointestinal problems. Fresh fecal samples were exposed to a direct smear examination, and modified Ziehl–Neelsen stain [1, 34]. Fecal specimens in three vials were received by the parasitology laboratory at TBRI. The vial contains 2.5% potassium dichromate, 10% formalin, or polyvinyl alcohol (PVA). Specimens submitted in PVA were processed and examined by modified Ziehl–Neelsen acid-fast stain.
Oocyst’s Preparation and Counting
The samples were examined and then stained with modified Ziehl–Neelsen acid fast stain (MZN), according to Garcia [15]. Briefly, stool samples were centrifuged at 500×g for 5 min, and the supernatant fluid was discarded. The sediment was washed twice in 1 ml of phosphate buffered saline (PBS) with centrifugation at 13,000×g for 2 min. Fecal debris was eliminated form samples after repeated washing and centrifugation.
Fifty µl of prepared cryptosporidium sample was counted under microscope. All mice groups were orally infected with C. parvum oocysts by means of esophageal tubes, excluding the control group, obtained from TBRI under the high-power field (HPF), and stained by Kinyoun’s acid-fast stain. Each animal was infected with about 10^4^ oocysts [12, 14].
Induction of Immunosuppression in Mice
The immune suppression in experimental rats was achieved by treatment with oral dexamethasone, a synthetic corticosteroid, by oral-gastric gavage for 14 consecutive days at a dosage of 0.25 mg/g/days prior to inoculation with Cryptosporidium oocysts [28].
Preparation of Lactoferrin
Cow milk colostrum samples were collected within 72 h after delivery (Faculty Dairy Farm of Behshahr, Mazandaran). After collection, the colostrum sample was placed in an ice flask and transported to the laboratory. The colostrum was centrifuged at 10,000 rpm for 20 min at 4 °C to separate the cream. The cream was discarded, while the skim (whey) was acidified to pH 4.6 using 2 N HCl and kept for 30 min at 40 °C. Acidic pH and heat caused the precipitation of casein. The acid whey was centrifuged at 10,000 rpm at 4 °C for 30 min, and the precipitate was discarded. The acid whey was neutralized to pH 6.8 with 2 N NaOH, and the neutralized whey was incubated at 65 °C for 30 min for pasteurization. Then the whey was precipitated using ammonium sulphate in two steps and centrifuged at 10,000 rpm at 4 °C for 30 min. After analyzing the proteins, we discard the sediment from the first stage. The second-stage sediment was filtered in phosphate-buffered saline (20 mM). Other proteins like β-lactoglobulin, α-lactalbumin, lactoperoxidase, and LF remain in the precipitation. LF was purified using carboxymethyl sephadex-C50 chromatography. Phosphate buffer (200 mM, pH 7.7) and a linear gradient of NaCl from 0.0 to 0.5 M were used, and LF was exited from 0.4 to 0.5 M of the NaCl linear gradient [23, 35].
Preparation of Lactoferrin Nano-Capsule
The lactoferrin nano (LF-NC) was prepared using a double emulsion/solvent evaporation technique. The lipids were dissolved in dichloromethane, and the aqueous phase was prearranged by dissolving the LF in PBS containing 30% Kolliphor® P407. Sonication solutions at 15% amplitude for 2 min (pulse: 30–5 s) to obtain the first emulsion. The second emulsion was formed by mixing the first emulsion with a α-Tocopherol PEG solution, sonicating at 20% amplitude for 5 min (pulse: 30–5 s), and evaporating. The subsequent LF-NC was then supplemented with a maturation medium, filtered, and ultracentrifuged for 1 h. LF-NC final concentration was homogenized in Milli-Q® water, measured, and lyophilized. The supernatant was retrieved for further studies [4, 22].
Experimental Design
Sixty male Swiss Albino mice were randomly assigned to six groups (10 per group) as follows:
G1: uninfected normal group.
G2: Infected with C. parvum oocysts (10^4^/mouse).
G3: Infected with C. parvum oocysts and treated with NZ (200 mg/kg b.wt./day) seven days post infection and continued for 3 days.
G4: Infected with C. parvum oocysts and treated with LF (150 mg/kg b.wt./day) seven days post infection and continued for 3 days.
G5: Infected with C. parvum oocysts and treated with LF-NC (150 mg/kg b.wt./day) seven days post infection and continued for 3 days.
G6: Infected with C. parvum oocysts and treated with LF-NC and with NZ. Each was injected separately with same doses. Seven days post infection and continued for 3 days.
Stool Examination
Stool samples were daily collected after infection and examined by staining with modified MZN stain, and C. parvum oocysts count was calculated using oil immersion magnification at × 100 magnification [34]. Evaluation of the oocyst shedding of C. parvum species in the fecal samples was done by collection, cleaning, and homogenization in PBS. The mean shedding was estimated 12 days post-infection (12th dpi).
Duodenal Content Examination
Mice were sacrificed on the 12th day to collect the duodenal content, and to homogenize in PBS to estimate and calculate the number of * [C. parvum* oocyst shedding [34].
Histopathological Examination
The mice were sacrificed at 12 days post-infection [6], the pancreatic tissues were embedded in paraffin wax blocks, and stained at the pathology laboratory in TBRI's. Expert histopathologists assessed the pathological dissimilarities, vascular wall changes, and Langerhans islet changes pattern abnormalities [11, 13].
Evaluation of Antioxidant and Oxidant Markers
Following intestinal tissue homogenization in PBS and centrifugation at 5000 rpm for 30 min, the supernatants were used to measure the oxidative stress biomarker. Malondialdehyde (MDA), and glutathione (GSH) were measured [17].
Cytokine measurements
The cytokines were determined using ELISA kits for IL-10, INF-γ, and TGF-β, according to the manufacturer instructions. The blood was collected, and centrifuged to collect the serum, and the supernatant was stored at -80°C.
Statistical Methods
The findings presented in this study represent the average values obtained from three independent replicates. The data were reported as the mean value ± SDE. The comparison between groups was conducted using a one-way analysis of variance (ANOVA). To estimate the presence of a substantial disparity between means, a Turkey post hoc analysis was performed to compare several groups. In the context of statistical tests, a P value less than 0.05 was deemed to be statistically significant. The data and statistical analysis were conducted using SPSS version 25.
Results
Fecal Cryptosporidium Oocyst Sheddings
The present study evaluated the impact of LF and LF-NC on the shedding of C. parvum oocysts in infected mice in comparison with the reference drug NZ. Table 1 shows the number of C. parvum oocyst sheddings and the reduction percentages (R%) in the different studied groups. The mean C. parvum oocyst shedding was 247,000 ± 1921in the untreated infected group. Mice that were treated with LF had a non-significant reduction in C. parvum to 201,000 ± 1306(9.9%).Mice that were treated with LF-NC had the C. parvum oocyst shedding significantly reduced to 197,000 ± 1011 (21.2%) (p < 0.001) compared to the infected mice that were treated with NZ, where the C. parvum oocyst shedding was significantly reduced to 54,000 ± 989 (79.1%) (p < 0.001).A significant reduction in C. parvum oocyst shedding was observed in a group of mice treated with LC-NC loaded with NZ (82.6%) (p < 0.001).Table 1. The total count of fecal and duodenal C. parvum oocysts in the different studied groupGroupsNo. of Cryptosporidium oocystsFecalDuodenalInfected untreated247,000 ± 1921344,000 ± 3217Infected/NZ54,000 ± 98990,000 ± 1322Infected /LF201,000 ± 1306259,000 ± 1989Infected /LF-NC197,000 ± 1011202,000 ± 1431Infected /LF-NC/NZ43,000 ± 76272,000 ± 1081Data was expressed as mean ± S.D.NZ Nanazoxid, LF Lactoferrin, LF-NC Lactoferrin nanocapsuleReduction percentage (R%). Number of oocysts in infected untreated/number of oocysts in treated group
Duodenal Content Examination
The mean number of C. parvum oocysts from the collected intestinal contents in each group of mice was calculated using MZN stain. The oocysts were seen as pink, bright-rose ovoid, and round bodies with varying intensity grades against a bluish background. The number of intestinal C. parvum oocysts shedding and the R% in different studied groups are illustrated in Fig. 1. The mean C. parvum oocysts shedding was 344,000 ± 3217 in the infected, untreated group. Mice that were treated with LF showed a non-significant reduction in C. parvum oocysts shedding (259,000 ± 1989,24.7%). The mice that were treated with LC-NC showed a significant reduction in C. parvum oocysts to 2202,000 ± 1431 (41.3%) (p < 0.01) compared to treatment with NZ. The C. parvum oocyst shedding was significantly reduced to (91,000 ± 1322, 73.5% (p < 0.001). A significant reduction in C. parvum oocyst shedding was observed in a group of mice treated with LC-NC loaded with NZ (79.1%) (p < 0.001).Fig. 1. The mean number reduction of C. parvum oocysts of the collected intestinal contents mice groups using MZN stain. A % reduction in fecal count and B % reduction in Duodenal content
Cytokine Measurements
The level of IL-10 in the infectednontreated group was 49.8 ± 4.5 pg/ml compared to its level in the normal group 4.35 ± 0.012 pg/ml. The infected mice were treated with NZ, LF, or LC-NC and showed different means in concentration levels of IL-10. The mean value of IL-10 was reduced to 12.2 ± 0.57 pg/ml in the group of infected that was treated by NZ. A significant reduction in the IL-10 level was observed in the group that received combined treatment of LF-NC and NZ. The levels of TGF-β in all groups are demonstrated in Table 2. The level of TGF-β was highly significant (p < 0.001) in the infected group (993.7 ± 112 pg/ml) as compared to the normal control (225.3 ± 17pg/ml). TGF-β levels were decreased in all treated groups as compared to the infected group. The maximum reduction in the level of TGF-β was observed in the group of mice that were treated by LF-NC combined with the NZ drug; the level was 331 ± 54 pg/ml, and in the group of infected mice that were treated with the NZ drug, the level of TGF-β was 413.5 ± 36 pg/ml. The mean level of INF-γ in the normal control group was 114.5 ± 12.1 pg/ml, while in the infected group, an increment in the concentration was observed at293 ± 11.5 pg/ml. In general, pre- and post-infection, levels of INF-γ increment were higher when compared to all other groups. The levels of INF-γ in groups of mice that were treated with NZ were 165 ± 16.3 pg/ml and 132.3 ± 14.5pg/ml in the group of mice that were treated with LF-NC combined with the NZ drug (Table 2).Table 2IL-10, TGF- β, and INF-γ levels were examined in various groupsGroupsIL-10TGF- βINF-γControl4.35 ± 0.012225.3 ± 17114.5 ± 12.1Infected49.8 ± 4.5993.7 ± 112293 ± 11.5Infected/NZ12.2 ± 0.57413.5 ± 36165 ± 16.3Infected /LF39.1 ± 5.7802.9 ± 109213 ± 22.1Infected /LF-NC23.2 ± 3.8562.3 ± 84181 ± 28.1Infected /LF-NC/NZ9.4 ± 0.72331 ± 54132.3 ± 14.5
Evaluation of Oxidative Stress and Antioxidant Markers
The level of MDA in normal control was 21.8 ± 0.678 mmol/g, while it was 45 ± 4.86 mmol/g in the infected control group as depicted in Fig. 2. Moreover, MDA levels decreased in the treated groups in comparison to the infected groups. A significant reduction in the MDA level was reported in the group that was treated with NZ and LF-NC. In addition, GSH level in the normal control group was 4.33 ± 0.179 mmol/g, while a significant decrement was shown in the infected group (1.93 ± 0.115 mmol/g; p < 0.001). The most significant level of GSH was observed in the group of mice treated with NZ and in the group treated with LF-NC (p < 0.001).Fig. 2A The level of Hepatic Malonaldehyde (MDA) and B glutathione (GSH) in experimental groups to measure the oxidative stress and antioxidant markers
Histopathological Examination
Inflammatory changes were detected in * [C. parvum*-infected groups, with a significant dysplastic change were observed in both infected and untreated groups (Fig. 3B). High-grade dysplasia was accompanying to the number of endogenous developmental stages of the parasite, with a higher mean number of oocysts in the infected, and untreated control group (Fig. 3B). Low-grade dysplastic changes were observed in the infected mice and treated with lactoferrin (Fig. 3C), while no dysplasia and no frank carcinoma developed in the groups treated with NZ or LF-NC (Fig. 3D–F).Fig. 3. Representative intestinal stained with Hematoxylin and Eosin in various groups. Control uninfected group (A), infected control (B), infected treated with NZ (C), infected treated with LF (D), infected treated with LF-NC (E), infected treated with LF-NC + NZ (F)
Discussion
In the current work, the therapeutic efficacy of nanazoxid (NZ), lactoferrin (LF), LF-NC, and LF-NC/NZ in treating immunocompromised mice experimentally infected with C. parvum oocysts was evaluated. Among the indicators used for defining the treatment efficacy of the used drugs was C. parvum oocyst shedding and subsequent counts in both stool and intestinal contents and calculating their percent reduction. * [C. parvum* oocyst numbers shed in stools and intestinal contents and their R% among the immunocompromised groups were assessed. It was noticed that * [C. parvum*-infected mice treated with NZ-loaded LF-NC showed the highest percentages of reduction in oocyst shedding (82.6%), followed by a group of mice that were treated with NZ alone (79.1%), in contrast to their equivalent infected untreated group. These findings were consistent with previous data [25]. The highest oocyst shedding was observed in infected and untreated immunosuppressed groups, reaching values of 344,000–3217/g of duodenal contents. This was in parallel with the study conducted by others [5], who disclosed that in severe immunodeficient mice, even when injected with a low parasite load, the parasite excretion increased, reaching mean oocyst numbers of more than 10,000/g feces on the 45th dpi. Correspondingly, another group found that the defeat of the immune system caused by dexamethasone made C. parvum organism clearance very problematic, increasing the duration and severity of infection in immunosuppressed mice [36]. However, when NZ was solely administered, it showed 79.1% and 73.5% reduction rates for C. parvum oocyst in stool and intestinal contents, respectively, with statistical significance as compared to the control-infected nontreated group. This agreed with earlier researchers, who established the success of NZ in both immunocompetent and immunocompromised groups [2].
Serum levels of IFN-y, TNF-α, and IL-10 were increased in infected and untreated groups. This finding was agreed upon by colleagues, who found that protection against this parasite has been associated with the production of IFN-, TNF-α, and IL-10 [9, 25]. Similarly, IL-10 was an important factor in an efficient anti-* [C. parvum* immune response [19]. The level of hepatic MDA in immunocompromised groups increased in infected groups as compared to normal controls. Hepatic MDA levels were found to be decreased in all treated groups as compared to the infected group. A significant reduction was observed in the group treated with LF-NC + NZ. Our data, in agreement with others, regarding the GSH designation for the liver of immunocompromised groups showed the level of GSH in the normal control group of mice was 4.33 ± 0.179; the infected group showed a decrease to 1.93 ± 0.115 mmol/g. The level of GSH in all treated groups was increased in the group that was treated with LF-NC + NZ [7].
The current study observed that the administration with NZ and LF improves mucosal damage in the infected group. Moreover, inhibit C. parvum colonization and multiplication via diminishing anti-cryptosporidial effects. Daily intake of NZ prevents C. parvum intestine tissue lesions in immunodeficient mice [3]. In the present study, the liver could also be considered the site of accumulation of the nanoparticles in the mice; in this context, a toxicological study was performed on this organ.
Conclusion
The current data demonstrated that NZ-loaded LF-NC, due to its non-toxic nature and minor anti-Cyclosporidium action, had the highest significant effectiveness in all subgroups. It is a potential drug that requires continuing efforts to treat cryptosporidiosis in both immunocompetent and immunocompromised humans.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Abubakar I, Aliyu S, Arumugam C, Usman N, Hunter PJ Bjocp (2007) Treatment of cryptosporidiosis in immunocompromised individuals: systematic review and meta-analysis. Brit J Clinic Pharmacol 3(4):387–39310.1111/j.1365-2125.2007.02873.x PMC 220323417335543 · doi ↗ · pubmed ↗
- 2Amadi B, Mwiya M, Sianongo S, Payne L, Watuka A, Katubulushi M, Kelly PJ Bid (2009) High dose prolonged treatment with nitazoxanide is not effective for cryptosporidiosis in HIV positive Zambian children: a randomised controlled trial. J BMC Infect Dis 9:1–710.1186/1471-2334-9-195PMC 279487419954529 · doi ↗ · pubmed ↗
- 3Benamrouz S, Guyot K, Gazzola S, Mouray A, Chassat T, Delaire B, Chabé M, Gosset P, Viscogliosi E, Dei-Cas EJPO (2012) Cryptosporidium parvum infection in SCID mice infected with only one oocyst: q PCR assessment of parasite replication in tissues and development of digestive cancer. J P Lo S One 7(12):e 51232.10.1371/journal.pone.0051232 PMC 352177323272093 · doi ↗ · pubmed ↗
- 4Boivin GP, Bottomley MA, Grobe NJ Jot A Af L As (2016) Responses of male C 57BL/6N mice to observing the euthanasia of other mice. J Am Assoc Lab Anim Sci 55(4):406–411PMC 494361027423146 · pubmed ↗
- 5Chandramathi S, Suresh K, Shuba S, Mahmood A, Kuppusamy UJP (2010) High levels of oxidative stress in rats infected with Blastocystis hominis. J Parasitol 137(4):605–61110.1017/S 003118200999135119961647 · doi ↗ · pubmed ↗
- 6Checkley W, White AC, Jaganath D, Arrowood MJ, Chalmers RM, Chen X-M, Fayer R, Griffiths JK, Guerrant RL, Hedstrom LJTLID (2015) A review of the global burden, novel diagnostics, therapeutics, and vaccine targets for cryptosporidium. Lancet Infect Dis 15(1):85–9410.1016/S 1473-3099(14)70772-8PMC 440112125278220 · doi ↗ · pubmed ↗
- 7Dayao DA, Sheoran A, Carvalho A, Xu H, Beamer G, Widmer G, Tzipori SJ Ijfp (2020) An immunocompetent rat model of infection with Cryptosporidium hominis and Cryptosporidium parvum. Int J Parasitol 50(1):19–2210.1016/j.ijpara.2019.10.00231759945 · doi ↗ · pubmed ↗
- 8Feldman AT, Wolfe D (2014) Tissue processing and hematoxylin and eosin staining. Methods Mol Biol 1180:31–4310.1007/978-1-4939-1050-2_325015141 · doi ↗ · pubmed ↗