Severe osteoarthritis in aged PANX3 knockout mice: implications for a novel primary osteoarthritis model
Brent Wakefield, Justin Tang, Julián Balanta-Melo, Jeffrey L Hutchinson, Rehanna Kanji, Geneva Herold, Brooke L O’Donnell, Courtney Brooks, Patti Kiser, Matthew W Grol, Cheryle A Séguin, Lilian I Plotkin, Frank Beier, Silvia Penuela

TL;DR
Deleting the PANX3 gene in mice leads to severe osteoarthritis in old age, suggesting a new model for studying the disease.
Contribution
A novel primary osteoarthritis model is proposed using aged PANX3 knockout mice.
Findings
Aged PANX3 knockout mice develop full-thickness cartilage lesions and severe synovitis.
Forced treadmill running did not worsen OA in PANX3 knockout mice.
PANX3 deletion negatively affects synovial joint health in aging.
Abstract
Osteoarthritis (OA) is a multifactorial disease associated with aging. As the molecular mechanisms underpinning the pathogenesis of this disease are unclear, there are no disease-modifying drugs to combat OA. Pannexin 3 (PANX3) has been shown to promote cartilage loss during posttraumatic OA. In contrast, the ablation of Panx3in male mice results in spontaneous full-thickness cartilage lesions at 24 mo of age. While protected from traumatic intervertebral disc (IVD) degeneration, Panx3KO mice show signs of IVD disease with altered disc mechanics. Whether the deleterious effects of ablating Panx3in aging are the result of accumulated mechanical damage is unknown. We used male and female WT and global Panx3KO C57Bl6 mice aged 18 mo of age. Mice were then randomized to sedentary (SED) or forced treadmill running (FEX) for 6 wk. Knee joint tissues, including the patellar tendon, quadriceps…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
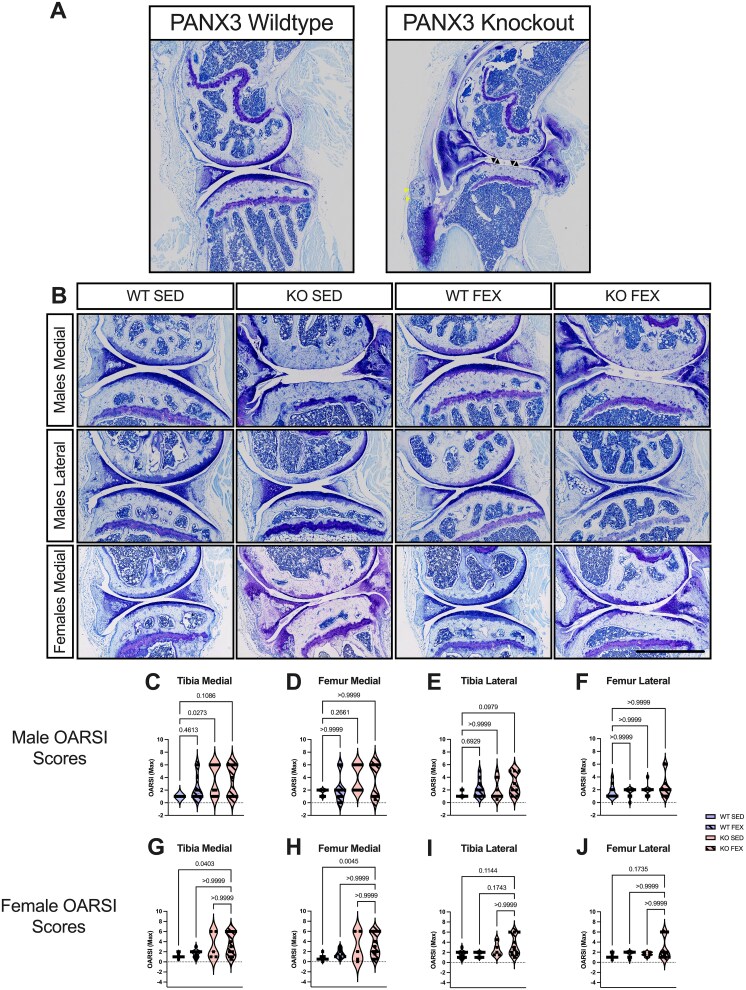 Figure 1
Figure 1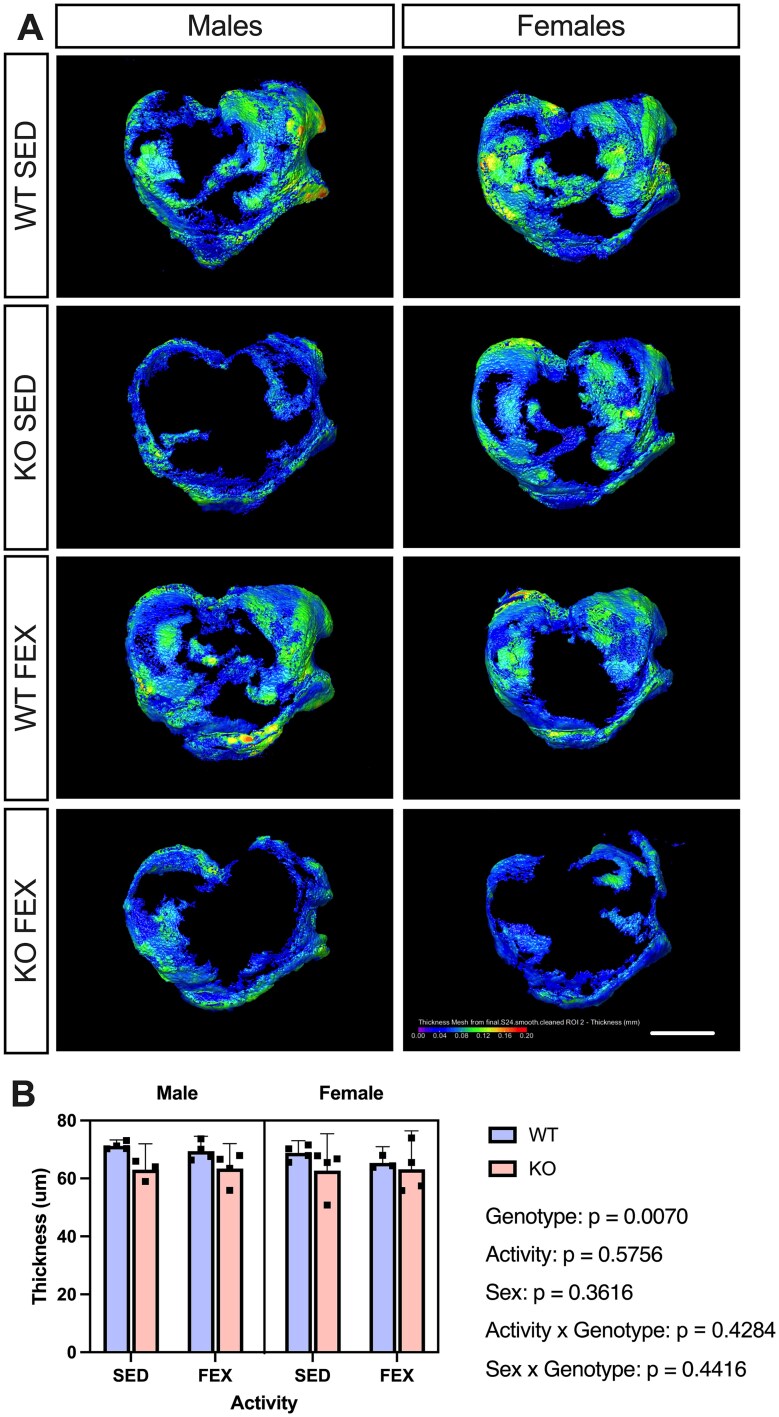 Figure 2
Figure 2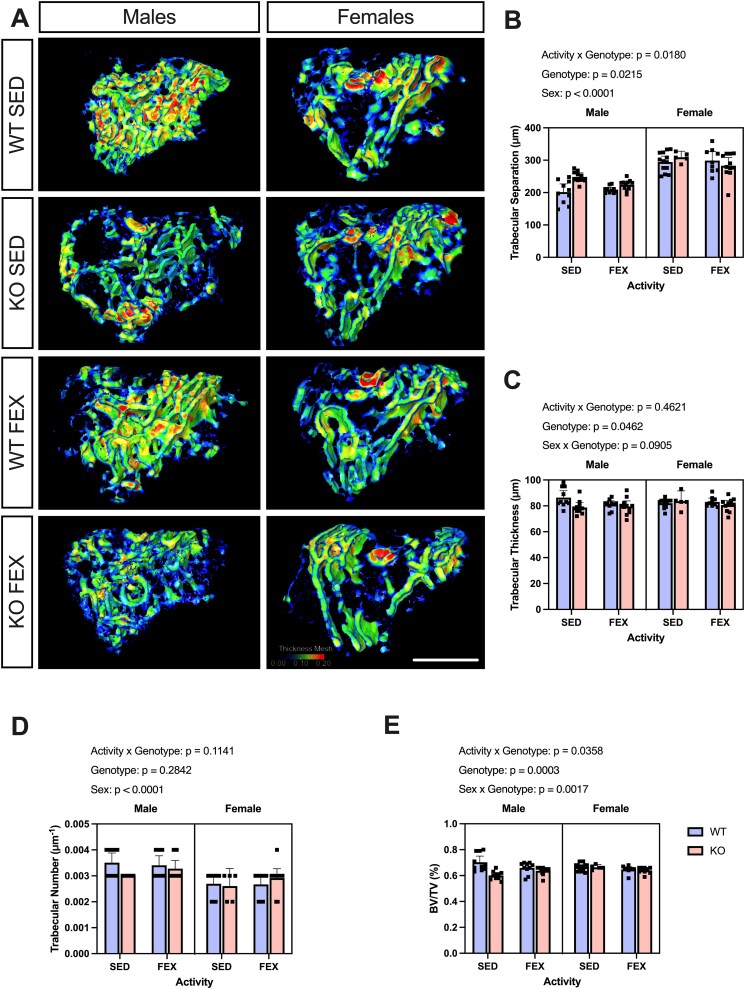 Figure 3
Figure 3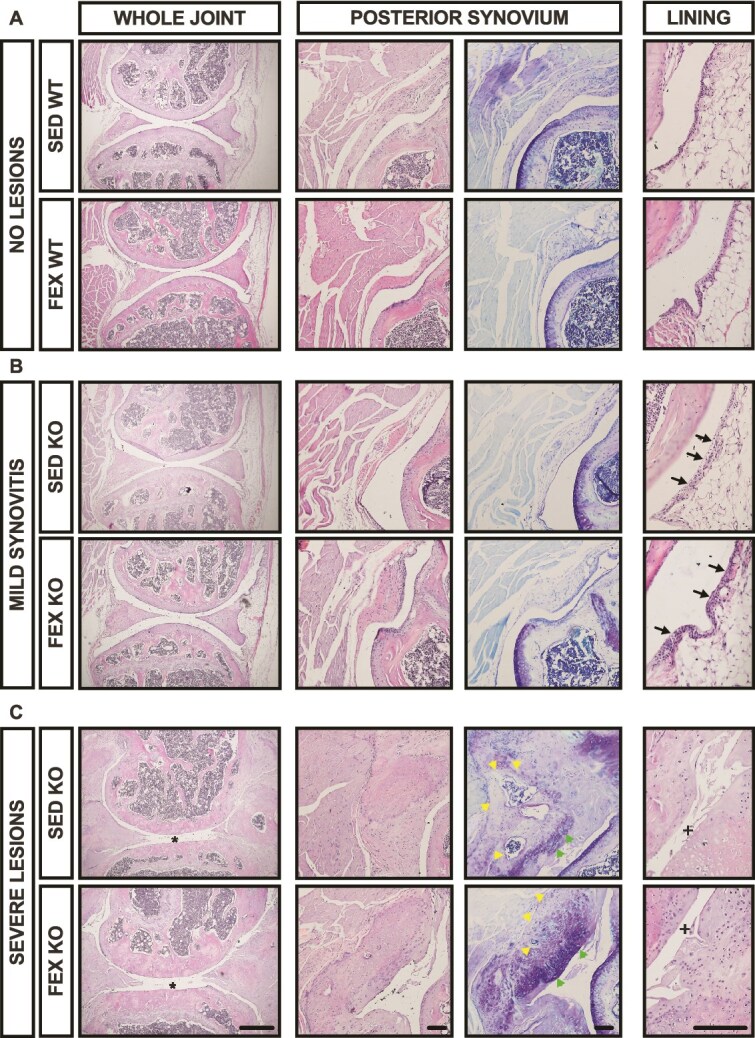 Figure 4
Figure 4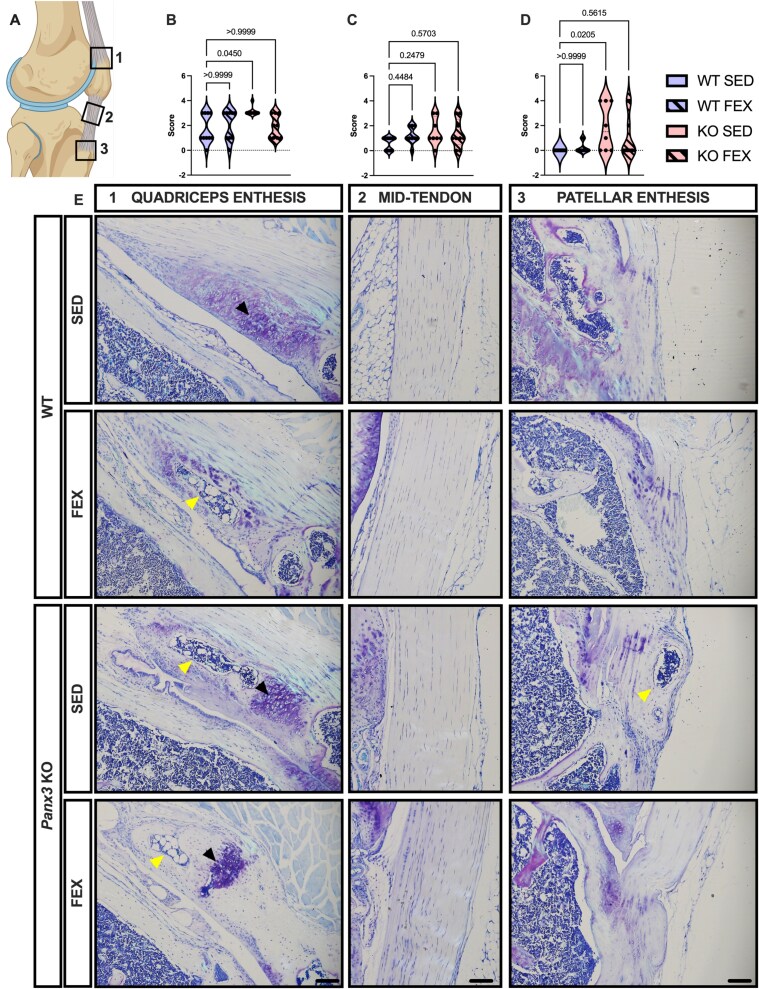 Figure 5
Figure 5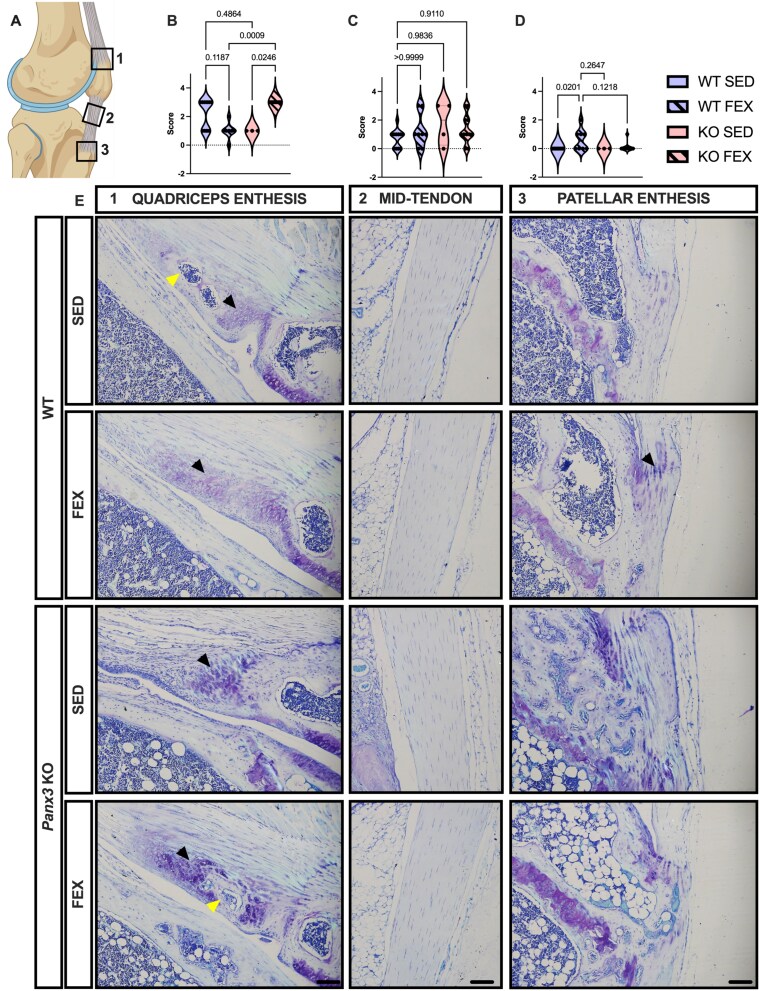 Figure 6
Figure 6|
|
|
|
| |
|---|---|---|---|---|
|
| – | |||
|
| 0.38, | – | ||
|
| 0.38, | 0.27, | – | |
|
| 0.58, | 0.32, | 0.19, | – |
- —Canadian Institute of Health Research operating
- —Natural Sciences and Engineering Research Council (NSERC)
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsOsteoarthritis Treatment and Mechanisms · Cytokine Signaling Pathways and Interactions · Cell Adhesion Molecules Research
Introduction
Globally, knee osteoarthritis (OA) affects millions—23% of the population over 40—causing disability that results in enormous personal and socioeconomic burdens.1While several risk factors exist for developing OA including sex, obesity, previous joint injury, joint shape and alignment, aging is the single greatest risk factor.2The molecular mechanisms that underpin this age-associated destruction of synovial joints are poorly understood.
Pannexin 3 (PANX3) is a channel-forming glycoprotein expressed in osteoblasts where it can act as a Ca^2+^release channel at the endoplasmic reticulum3and in chondrocytes where it can act as an adenosine triphosphate (ATP) release channel at the cell membrane.4PANX3 is also expressed in annulus fibrosus (AF) cells of the intervertebral disc (IVD)5^,^6as well as many human tissues.7In an early rat model of traumatic OA (anterior crucial ligament transection and partial medial meniscectomy), Panx3mRNA expression was upregulated in OA cartilage compared to control knees.8Additionally, in a 30-wk-old mouse model, PANX3 was upregulated in cartilage following destabilization of the medial meniscus (DMM) surgery to induce post-traumatic OA.9This increased expression of PANX3 in diseased cartilage suggests its mechanistic involvement in traumatic OA development in rodent models. In fact, Moon et al. performed the DMM surgery on global and chondrocyte-specific Panx3KO mice and found that these mice were strongly protected from OA compared to WT mice.9Similarly, in an IVD injury model, Panx3KO mice had fewer hypertrophic cells of the AF, and the AF structure was largely preserved compared to WT mice.6These two models suggest that the absence of PANX3 is protective in traumatic/injury-induced joint disease, however, its role in primary OA may be different as well as its expression in aged tissues. In humans, PANX3 is upregulated in OA cartilage tissue,9and noncoding intronic single nucleotide polymorphisms of PANX3are strongly associated with chronic low back pain.10These data suggest that PANX3 function in cartilage is conserved across rodents and humans and may be an important molecular player of OA, with potentially different mechanisms in post-traumatic OA versus age-related primary OA.
Previous studies have shown that aging influences the genetic expression patterns of joint tissues in response to stress/injury,11–13and therefore, aged models are required to better understand the pathobiology of age-associated OA. To this point, male Panx3KO mice at 18 or 24 mo of age showed accelerated cartilage erosion, subchondral sclerosis, and synovitis of the knee joint,14which was in contrast to the previously seen protective effects in the DMM model in adult mice.9An important difference between the two studies is that the aged mice were given a running wheel in their cage for environmental enrichment, which could have contributed to the different effects of Panx3KO on joint tissues. In the IVD, we have shown that uninjured IVDs are sensitive to aberrant biomechanical loading,6again highlighting the context-dependent function of PANX3 in joint health. To gain more insight into how mechanical loading affects joint wear and tear in conjunction with a genotypic deletion of Panx3, we recently examined a forced-exercise model on a 24-30-wk-old with a treadmill running protocol. Consistent with our prior findings, Panx3exhibited protective effects on the knee joints, as observed with Panx3KO mice exhibiting more pronounced superficial defects in tibial cartilage. Additionally, PANX3 mice displayed greater histological features of IVD degeneration (IDD) after forced treadmill running.15
In this study, we investigated how aging and excessive mechanical use, via forced treadmill running as applied in the 24-30-wk-old mice, influence joint pathology in Panx3KO mice. Panx3KO mice demonstrated a bimodal distribution in which roughly half of the animals, regardless of forced running, had full-thickness cartilage erosion down to the subchondral bone and expanded synovium containing ectopic calcification and fibrocartilage with scattered lymphocytes, while the other half had mild synovitis. In contrast, all WT mice had mild signs of superficial cartilage erosion and synovitis under either treatment. Male Panx3KO mice also displayed cartilage, bone, and bone marrow in the quadriceps enthesis reminiscent of enthesitis. The degree to which these mice had enthesitis or tendinopathy of the knee was correlated with OA scores. At the lumbar spine, Panx3KO mice IVDs were histologically similar to WT mice. These results suggest that aged Panx3KO mice, regardless of sex and activity, develop severe age-related knee joint pathology, including OA in aging.
Materials and methods
Mice
Animals used in this study were bred in-house and euthanized in accordance with the ethics guidelines of the Canadian Council for Animal Care. Animal use protocols were approved by the Council for Animal Care at Western University Canada (AUP 2019-069). C57BL/6N Mice were housed in standard shoe box-style caging, exposed to a 12-h light/dark cycle, and ate regular chow ad libitum. WT and Panx3KO mice were congenic.9DNA was collected from ear clippings of each mouse to determine the genotype using polymerase chain reaction as previously described.9^,^16At sacrifice, mouse knees and spines were collected and immediately processed for histological analysis.
Forced treadmill running
At 18 mo, mice were randomized to either a no-exercise group (sedentary, SED) or a forced treadmill running (forced-exercise, FEX) group. FEX groups ran on a treadmill (Columbus Instruments, Ohio) for 6 wk, 1 h a day for 5 d a wk, at a speed of 11 m/min and a 10° incline—an adapted protocol that has previously been used to induce OA in male C57BL/6 mice.17All mice were continuously monitored by an operator in the room to ensure the completion of the running protocol. Additionally, the mice were encouraged to run using a bottlebrush bristle and a shock grid at the end of the treadmill per the animal ethics protocol.
Micro-CT (μCT) imaging and analysis
Knee joints were fixed by immersion in 10% neutral buffered formalin at room temperature for 24 h, prior to storage in 70% ethanol. On the day of scanning, all unstained samples were kept submerged in 70% ethanol and imaged with an 1172 SkyScan μCT at 4.83 μmresolution using a source voltage of 59 kV, a source current of 167 μA, and an Al 0.5 mm energy filter. A rotation range of 180° with a rotation step of 0.5° and frame averaging of two was applied. Positioning of the samples during scanning was standardized, with the distal femur superior and the proximal tibia inferior in the scan view. Subsequent cartilage staining with phosphotungstic acid (PTA) dissolved in 70% ethanol was then conducted on a subset of knee samples and imaged with the same parameters and protocol as stated above. Afterwards, all scans were reconstructed in NRecon software (v.1.7.0.4, Bruker micro-CT) using a smoothing width of three, a beam-hardening correction of 100%, and a ring artifact correction of 20. The nomenclature used follows the recommendation of the American Society for Bone and Mineral Research.18
Pre-segmentation of the cartilage was conducted through Dragonfly (v.2022.1, Object Research Systems) and final segmentation was accomplished with the Biomedisa application (v.23.08.1, Australian National University CTLab) to measure cartilage thickness. Additionally, the subchondral bone was evaluated through Bone Analysis Wizard in Dragonfly to obtain average trabecular separation (Tb.Sp), average trabecular thickness (Tb.Th), trabecular number (Tb.N), and bone volume fraction (BV/TV). The optimal segmentation thresholds were automatically selected through the Otsu algorithm built into the Dragonfly software.
Histopathological assessment of the knee joint
At the experimental endpoint, a separate set of knee joints was fixed in 4% paraformaldehyde at room temperature for 24 h on a shaker and then decalcified in 5% EDTA for 12 d at room temperature. Knees were processed and embedded in the sagittal plane in paraffin, and 6-μm-thick sections were cut from front to back. Sections were stained with toluidine blue. Three sections from the medial and lateral compartments were scored by two blinded reviewers using the OA Research Society International (OARSI) recommendations for histological assessments of OA in the mouse.19The average max score of each sample was then used for statistical analysis.
Synovial tissue pathological assessment
Considering the severity and bimodal distribution of the phenotype in these animals, we opted to take a more descriptive approach when describing the synovial changes that were occurring within these animals versus a semi-quantitative analysis. A pathologist with experience describing joint disease in animal models investigated the occurrence of specific pathological features observed in the synovium of the animals. The pathologist was blinded to all genotypes, sex, and activity. One H&E-stained section of the medial load-bearing region per animal was selected for analysis.
Histopathological assessment of the lumbar intervertebral discs
Lumbar spines were fixed for 24 h with 4% paraformaldehyde, followed by 7 d of decalcification with Shandon’s TBD-2 (Thermo Fisher Scientific) at room temperature. Tissues were embedded in paraffin and sectioned in the sagittal plane at a thickness of 5 μm. Mid-sagittal sections were deparaffinized and rehydrated as previously described20and stained using 0.1% Safranin-O/0.05% Fast Green. Sections were imaged on a Leica DM1000 microscope with Leica Application Suite (Leica Microsystems: DEU). To evaluate IDD, spine sections were scored by two observers blinded to age, exercise, sex, and genotype using a previously established histopathological scoring system for mouse IVDs.21To report on degeneration across the lumbar spine, scores for individual lumbar IVDs (L2-L6) were averaged, and the total score was plotted for each individual mouse.
Enthesis and patellar tendon analysis
Mid-tendon sections were stained with toluidine blue and scored for distal quadriceps tendon enthesitis, distal patellar tendon enthesitis and patellar tendinopathy. Enthesitis was scored using the following parameters: 0 = normal; 1 = ectopic cartilage; 2 = ectopic cartilage/bone; 3 = ectopic cartilage/bone with a marrow cavity; 4 = ectopic bone with a marrow. Tendinopathy was scored using the following system: 0 = normal; 1 = increased cellularity; 2 = cell rounding/clustering; 3 = chondrogenesis (ie, proteoglycan-rich matrix, hypertrophy); 4 = ectopic cartilage/bone.
Statistics
The Department of Epidemiology and Biostatistics at Western University was consulted to determine the appropriate statistical analysis. Data are presented as stated in the respective figure. Prism 9 (GraphPad Software Inc.) Version 9.4.1 (458) was used to run all statistical tests, including one-way ANOVA or two-way and three-way ANOVA for comparison. For OARSI scores, histopathological scores of the IVD, enthesitis and tendinopathy scores, males and females were analyzed separately within their respective genotypes and activity groups. A Kruskal–Wallis test was used with an uncorrected Dunn’s test for multiple comparisons to determine statistical differences among the groups. Correlations were performed using Pearson’s r. All applicable data met assumptions for homoscedasticity or normality of residuals. Based on the recommendations of the editorial entitled: Moving to a World Beyond “p < .05,”22the data is referred to in terms of weak (p < .05), moderate (p < .01), or strong (p < .001) statistical evidence. Please see Tables S1–S5.
Results
Panx3KO mice develop full-thickness cartilage erosion of the tibia and femur surface in aging
We first assessed whether Panx3KO mice develop histopathological OA compared to WT mice in aging, and whether forced treadmill running would influence these outcomes. Toluidine blue-stained, paraffin-embedded sections from the knee joints of 18-mo-old WT and Panx3KO mice under SED or FEX conditions was analyzed (Figure 1A). We immediately noticed obvious osteoarthritic characteristics in the Panx3KO samples, including erosion of the cartilage and boney deposits in the enthesis (Figure 1A; Figures S3and S4). Consistent with our previous findings, closer histological assessment of the male mice observed full-thickness cartilage lesions in half of the Panx3KO mice in the medial tibial and femoral surfaces but not in any of the WT mice [13.15 mean rank difference, p = .0273; 10.33, p = .2661] (Figure 1B–D). In the lateral compartment, there was no statistical evidence for differences between SED WT and SED Panx3KO mice [4.011, p > .9999; 3.743, p > .9999] (Figure 1B, E, and F). Regarding the effect of forced treadmill running, there was no statistical evidence that the addition of forced treadmill running influenced the cartilage structure in both knee compartments of WT mice for the tibia [8.200, p = .4613; 7.311, p = .6929] and the femur [0.5333, p > .9999, 0.7556, p > .9999]. In the lateral compartment, there was no statistical evidence that FEX Panx3KO mice had worse OARSI scores compared to SED WT mice, as 3 mice had full-thickness lesions of the tibia [11.80, p = .0979] (Figure 1E); and there was no statistical evidence for differences in femoral OARSI scores among the groups [5.419, p > .9999] (Figure 1F). Taken together, this suggests that some male Panx3KO mice develop histological features of severe OA only in the medial compartment of the tibia (approximately 50% phenotypic penetrance).
Male and female PANX3 KO mice develop full-thickness cartilage erosion in the tibia and femur in aging. Representative whole joint sagittal images stained with toluidine blue were taken at 10× magnification (scale bar = 1000 μm) at the medial and lateral compartments (A-B). (A) Full-view comparison of a PANX3 WT versus KO joint. (B) Representative images of cartilage from male and female PANX3 WT and KO mice under SED and forced treadmill running (FEX) conditions, as indicated. (C-J) violin plots delineating the distribution/grouping of OARSI max scores for male medial tibia (C) and femur (D), male lateral tibia (E) and femur (F), female medial tibia (G) and femur (H), and female lateral tibia (I) and femur (J). WT SED (N = 10 males, N = 12 females), WT FEX (N = 10 males, N = 9 females), KO SED (N = 10 males, N = 5 females), KO FEX (N = 9 males, N = 11 females). For statistical comparisons among the groups, a Kruskal–Wallis test followed by a Dunn’s multiple comparisons test was performed. Internal arrows point to full-thickness cartilage loss, peripheral (yellow) arrows point to boney deposits in the enthesis.
Next, we performed the same histological analysis of female knees from aged WT and Panx3KO mice under SED and FEX conditions. Note, female mice had not been included in our earlier aging study.14Like in males, several female Panx3KO mice developed full-thickness cartilage erosion in the medial compartment (6 mice in total), while no WT mice developed such erosion [11.79, p = .0403; 14.95, p = .0045] (Figure 1B, G, and H). Forced treadmill running seemed to have little to no effect on cartilage structure in the medial compartment [1.700, p > .9999; 5.255, p > .9999] (Figure 1B, G, and H). In the lateral compartment, while some SED Panx3 KO mice showed signs of cartilage erosion, full-thickness erosion was only observed in some Panx3KO mice that were forced to run (N = 3), but there was no statistical inference to suggest differences between these groups (Figure 1I and J). This data suggests that, like male Panx3KO mice, female mice exhibit a bimodal distribution, with a subset of mice developing full-thickness cartilage erosion, and the addition of forced treadmill running may influence the development of cartilage loss of the tibia in the lateral compartment with approximately 50% phenotypic penetrance.
Given we saw full-thickness lesions of the tibia, we conducted further μCT analysis on PTA-stained cartilage samples with the contralateral knee of the WT and Panx3KO. In support of our histopathological observations, all Panx3KO mice demonstrated moderate statistical evidence for reduced tibial cartilage thickness in comparison to WT mice [F (1, 22) = 8.83, p = .0070], regardless of sex and activity (Figure 2A and B). This data suggests that both Panx3KO male and female mice experience cartilage erosion throughout the whole cartilage of the tibia even without forced exercise, as no statistical effect was observed between activity groups. In comparison to our previous work,23may suggest the development of cartilage loss of the tibia is due to a combination of both natural aging and genotype.
PANX3 KO mice have reduced cartilage thickness when examined through micro-CT. Representative micro-CT images of knee cartilage samples stained with phosphotungstic acid from a male (left panel) and female (right panel) PANX3 WT and KO mice subjected to SED or forced treadmill running (FEX) conditions, as indicated (A). Images are shown from the superior view (scale bar = 1 mm). (B) Corresponding micro-CT cartilage thickness measurements of treatment groups WT SED (N = 4 males, N = 4 females), WT FEX (N = 4 males, N = 3 females), KO SED (N = 3 males, N = 4 females), KO FEX (N = 4 males, N = 4 females). For statistical comparisons among the groups, a three-way ANOVA with sex × genotype × activity as factors followed by Tukey’s multiple comparisons test was performed. All data are shown as means ± CI.
Panx3KO mice exhibit reduced Tb.Th and secondary ossification center volume in the proximal tibia
Building on the established findings regarding the pivotal role that PANX3 plays in the regulation of osteoblast differentiation, mechanically induced bone modeling, and bone formation,3^,^24^,^25our prior observations in 24-30-wk-old mice revealed reductions in the BV/TV of the proximal tibia.23Considering these insights and the significant articular cartilage degeneration we are witnessing in the current study, we next wanted to identify any pathological changes in the secondary ossification center of the proximal tibia through μCT imaging (Figure 3A). Panx3KO mice were observed to have weak statistical evidence of an increase in average Tb.Sp [F (1, 71) = 5.53, p = .0215], which is representative of cavities in the bone marrow (Figure 3A and B), and a complementary decrease in average Tb.Th [F (1, 71) = 4.11, p = .0462] (Figure 3A and C). Additionally, there was weak statistical evidence for a two-way interaction of activity x genotype [F (1, 71) = 5.87, p = .0180], where the presence of forced treadmill running seemed to reduce the genotypic increase in average Tb.Sp of the male Panx3KO mice and completely reverse its affects in the female mice (Figure 3B). However, there was no evidence of any genotypic or activity changes to Tb.N [F (1, 71) = 2.56, p = .1141] (Figure 3D). Despite this, further analysis of the tibial secondary ossification center revealed strong evidence for a genotypic effect on BV/TV [F (1, 71) = 14.1, p = .0003], with Panx3KO mice has reduced bone volume compared to WT mice (Figure 3E). There was also weak statistical evidence for an interaction between activity and genotype [F (1, 71) = 4.58, p = .0358], and moderate statistical evidence for a sex x genotype interaction [F (1, 71) = 10.6, p = .0017]. More specifically, a SED lifestyle seems to exacerbate the bone volume reduction observed in Panx3KO mice, whereas this bone volume decrease also appears to be absent in females compared to males (Figure 3E). Altogether, this data suggests that a Panx3deletion leads to a reduction in trabecular bone within the secondary ossification center of the proximal tibia and the presence of forced exercise may mitigate the extent of this alteration.
A PANX3 deficiency alters subchondral bone characteristics in aged mice. Representative micro-CT 3D renderings of male (left panel) or female (right panel) mouse knee subchondral bone samples from indicated genotype and treatment group (A). Scale bar = 1 mm. (B-E) bar graphs displaying the average Tb.Sp (B), average trabecular bone thickness (C), average Tb.N (D), and subchondral BV/TV (E). WT SED (N = 10 males, N = 13 females), WT FEX (N = 10 males, N = 9 females), KO SED (N = 10 males, N = 5 females), KO FEX (N = 11 males, N = 11 females). For statistical comparisons among the groups, a three-way ANOVA with sex × genotype × activity as factors followed by Tukey’s multiple comparisons test was performed. All data are shown as means ± CI.
Panx3KO mice have comparable body weights to WT mice under SED and forced treadmill running conditions within their respective sex
Having previously published large reductions in body weights of Panx3KO mice compared to WT mice in adulthood,24we next wanted to investigate whether the observed changes in articular cartilage and trabecular bone were attributable to alterations in mechanical loading that stemmed from differences in body weight between the genotypes. Hence, we also investigated the effect of deleting Panx3on body weight in aged animals (Figure S1) and found that those genotypic differences were diminished with age. There was no statistical evidence for differences in body weight between genotypes in both males [F (1, 36) = 0.2273, p = .6364] (Figure S1A) and female [F (1, 32) = 3.207, p = .0828] (Figure S1B) mice. While there was no statistical evidence for body weight differences between activity groups in males [F (1, 36) = 0.09, p = .7574], there was weak statistical evidence for female mice to have lower body weights when forced to run on a treadmill for 6 wk compared to SED female mice [F (1, 32) = 5.37, p = .0270]. This data suggests aged WT and Panx3KO mice have similar body weights under SED and FEX conditions, thereby suggesting body weight differences are unlikely to be a contributing factor for the observed genotypic disparities in the joint tissue.
Panx3KO mice develop mild to severe synovitis under both SED and forced treadmill running conditions
Considering the bimodal distribution seen in our KO mice, and upon observation of the severe synovial changes in the mice with full-thickness erosions, we chose to take a descriptive approach for the synovial analysis. Synovial tissue was assessed by a blinded pathologist with extensive experience describing animal model synovial tissue. H&E-stained sections in the medial load-bearing zone were assessed. Three distinct joint morphological phenotypes were observed across the groups: (1) no lesions (Figure 4A); (2) mild acute synovitis (Figure 4B); and (3) severe diffuse synovial fibrosis with ectopic ossification (Figure 4C). All samples with mild to severe synovitis were from Panx3KO mice, except for one WT sample. Synovium of Panx3KO mice with intact cartilage consisted of acute lymphocytic synovitis, where the synovium was expanded by lymphocytes and a few macrophages (Figure 4B, mild synovitis). Full-thickness cartilage erosion in the Panx3KO mice coincided with severe diffuse synovial fibrosis, ulceration, and ectopic ossification (Figure 4C, severe synovitis). These mice had locally extensive to complete effacement of the synovium by collagen, fibrocartilage, and in some cases, bone interrupted by areas of acellular basophilic material. These findings suggest that aged Panx3KO mice develop mild-to-severe synovitis of the knee joint, which coincides with the severity of cartilage erosion.
Panx3KO mice develop mild to severe synovitis of the knee in aging. Sagittal sections of the medial compartment were stained with H&E and assessed by a blinded pathologist. Slides were assessed for signs of immune cell infiltration of the synovial lining (black arrows), indicating mild synovitis, which was a phenotype of Panx3KO mice (one WT mouse was characterized to have mild synovitis). Ectopic ossification (left, yellow arrows) and fibrocartilage (right, green arrows) were observed exclusively in Panx3KO mice and were characterized as severe lesions. * denotes cartilage erosion. + denoting loss of synovial lining. Whole joint images (left) are 4× magnification. Scale bar = 500 μm. The posterior synovium (middle) is 10× magnification. Scale bar = 100 μm. The lining (right) images are 20× magnification. Scale bar = 100 μm.
Panx3KO mouse IVDs age normally in males and females, even when forced to treadmill run
To investigate the role of PANX3 in age-associated IDD, and whether forced treadmill running in aging influences’ disc health, we next analyzed the IVDs for histological changes. WT and Panx3KO mice were aged to 18 mo and lumbar spines were analyzed histologically as described in the Methods section. Under both SED and FEX conditions, male (Figure S2A and B) and female (Figure S2C and D) Panx3KO IVDs appeared normal relative to WT controls with no differences in histopathological features of degeneration in the nucleus pulposus and the AF at any disc height (Figure S2A and C) or when averaged across the lumbar spine (Figure S2B and D). This data suggests that the IVDs of both male and female Panx3KO mice, age and respond to forced treadmill running similarly to their WT counterparts as no effect was observed under the applied conditions.
Male Panx3KO mice display ectopic cartilage and bone deposits containing marrow in the enthesis of the quadriceps and patellar tendons
Considering the enthesis originates from fibrocartilage cells that are highly responsive to mechanical loading,25we next analyzed the patellar tendon for signs of tendinopathy and enthesitis at the distal patella and quadriceps tendons (Figures 5and 6A). In male mice, there was weak statistical evidence [12.26, p = .0450] suggesting SED Panx3KO mice develop enthesitis of the quadriceps tendon enthesis, consisting of cartilage and bone deposits, often with a marrow cavity (Figure 5B). Throughout the patellar tendon, there was no statistical evidence for histopathological cellular changes in any of the groups [p = .1473] (Figure 5C). Like the quadriceps enthesis, the distal patellar tendon enthesis of SED Panx3KO mice showed signs of enthesitis, including deposits of cartilage and bone, often with a marrow cavity, compared to SED WT mice [11.31, p = .0205] (Figure 5D). In female mice, we performed the same semi-quantitative analysis of the enthesis and patellar tendon (Figure 6A). At the quadriceps enthesis, there was weak statistical evidence [14.50, p = .0246] that Panx3KO mice develop enthesitis with forced treadmill running (Figure 6B), with all the mice showing ectopic cartilage and bone, often with marrow formation, within the quadriceps tendon enthesis (Figure 6E). Throughout the patellar tendon, there was no statistical evidence [p = .4007] for histopathological changes to cellular shape or distribution between any of the groups (Figure 6C). At the patellar tendon enthesis, there was weak statistical evidence [8.063, p = .0201] that WT mice develop enthesitis with forced treadmill running, with no statistical evidence of histological differences in Panx3KO mice (Figure 6D). This data suggests that Panx3KO mice develop enthesitis of the quadriceps and patellar tendon entheses during aging, while forced treadmill running in female WT mice produced enthesitis at the distal patellar tendon enthesis. We next ran a Pearson’s correlation between medial OA scores and the enthesitis and tendinopathy scores to determine if there was a relationship between which mice develop OA and those that develop enthesis and tendon pathology. There was strong statistical evidence suggesting that higher enthesitis and tendinopathy scores coincided with higher OA scores (Table 1).
Sedentary male Panx3KO mice show signs of patellar and quadriceps enthesitis. Male WT and Panx3KO knee sections were stained with toluidine blue, sectioned in the sagittal plane and scored for quadriceps enthesitis (A1), patellar tendinopathy (A2), and patellar enthesitis (A3). Sedentary, and forced treadmill running (FEX). Violin plots showing distribution/grouping of histological scores for quadriceps enthesitis (B), tendinopathy (C), and patellar enthesitis (D). Representative toluidine blue sagittal sections (E). 10× magnification. Scale bar = 100 μm. Black arrows point to cartilage, yellow arrows point to bone and marrow. WT SED (N = 8), WT FEX (N = 10), KO SED (N = 12), and KO FEX (N = 11). For statistical comparisons among the groups, a Kruskal–Wallis test was performed.
Female Panx3KO mice develop quadriceps enthesitis with forced treadmill running. Female WT and Panx3KO knee sections were stained with toluidine blue, sectioned in the sagittal plane and scored for quadriceps enthesitis (A1), patellar tendinopathy (A2), and patellar enthesitis (A3). Sedentary, and forced treadmill running (FEX). Violin plots showing distribution/grouping of histological scores for quadriceps enthesitis (B), tendinopathy (C), and patellar enthesitis (D). Representative toluidine blue sagittal sections (E). 10× magnification. Scale bar = 100 μm. Black arrows point to cartilage, yellow arrows point to bone and marrow. WT SED (N = 14), WT FEX (N = 8), KO SED (N = 4), and KO FEX (N = 11). For statistical comparisons among the groups, a Kruskal–Wallis test was performed.
Discussion
In this study, we found that 18-mo-old male and female Panx3KO mice demonstrated severe OA of the knee and WT control mice demonstrated milder OA, while IVDs seem to be histologically comparable between genotypic groups. The addition of forced mechanical loading through treadmill running does not exacerbate this phenotype in the medial compartment, while a subset of FEX Panx3KO mice had full-thickness lesions in the lateral compartment, suggesting a compartment-specific effect of forced treadmill running in Panx3KO mice. The presence of forced exercise also attenuated the increase in average Tb.Sp and decrease in BV/TV of the proximal tibial secondary ossification center for Panx3KO mice in comparison to WT. This observation may result from increased bone remodeling triggered by mechanical stress, where the bone adapts to increased loading by minimizing the space between the trabeculae to strengthen its framework. However, due to the male Panx3KO mice already having a more osteopenic phenotype, and hence a greater Tb.Sp and lower BV/TV, this interaction manifests as less pronounced changes in the separation or bone volume between the genotypes. It is notable that in the histology we witnessed the presence of blood entering the joint cavity, which may be an indication that minor trauma or stresses on the bone can result in a fracture that causes bleeding into the joint space. Additionally, a fracture may also contribute to the development of OA overtime by altering the biomechanics and stability of the joint. Additionally, Panx3KO mice develop mild to severe synovitis consisting of lymphocyte infiltration, ectopic fibrocartilage and calcification of the knee joint. Male Panx3KO mice also appeared to have histological features of quadriceps and patellar tendon enthesitis under SED conditions, whereas female Panx3KO mice developed quadriceps tendon enthesitis with forced treadmill running. Within lumbar spine IVDs, both male and female Panx3KO mice had similar histopathological features compared to the WT controls. Even with the stress of forced treadmill running, there was no statistical evidence for histopathological differences among the groups, suggesting running later in life is not detrimental to disc structure in either genotype.
Full-thickness cartilage erosion is not a normal histological feature of knee joints in aged mice.26In this study, the full-thickness cartilage loss observed in our animals was accompanied by erosion and fibrillation of adjacent cartilage surfaces. Additionally, using μ-CT contrast imaging of cartilage in the contralateral knee, we found that these mice had reduced thickness in the Panx3KO mice. However, it is notable that the representative images show full cartilage loss in both the WT and KO mice, albeit more in mice with the latter genotype. But given that none of the WT mice had full-thickness cartilage loss in the histological analysis, this supports that the ulcerations we see may be artifacts from processing (ie, ineffective PTA-staining of degenerative areas) thereby making it hard to determine if there are the presence of premortem full-cartilage lesions. All this is not to suggest that the cartilage of these Panx3KO mice is not degenerating, but rather histological and μCT representation may not accurately depict the in vivo state of the cartilage tissue premortem.
In addition to cartilage erosion, we saw mild to severe synovitis in the Panx3KO animal knees, which included extensive fibrocartilage and calcification deposition within the synovium. Given this phenotype was not detected in WT mice, we suspect that it may be Panx3KO specific. Whether PANX3 is expressed in synovial tissue has not been determined; however, in silico data suggests PANX3should be expressed in human synovial fibroblasts.27Additionally, considering the severe inflammatory phenotype, we are unaware of any indication that macrophages or lymphocytes express *Panx3.*Future studies should investigate the periarticular expression and function of Panx3in joint tissues.
Considering the evidence showing PANX3’s role in cartilage, it is possible that the synovial phenotype is initiated by cartilage degradation and subsequent synovitis. Chronic release of damage-associated molecular patterns or other catabolic signals (eg, cytokines) from degrading cartilage into the synovial fluid space may activate synovial lining macrophages.28In our previous report challenging 30-wk-old Panx3KO mice, we showed superficial cartilage erosion which was exacerbated by forced treadmill running and resulted in moderate evidence of synovial lining thickening, suggesting early OA development23A lifetime of cartilage erosion may be chronically stimulating synovial macrophages leading to these pathological changes. Previously, we found that aged Panx3KO mice had low lubricin expression in the superficial zone of the articular cartilage.14Lubricin is an essential lubricating protein for the joint surface,29and in vitro models have shown that lubricin has anti-inflammatory effects on synovial lining fibroblasts by binding to toll-like receptors two and four.30Taken together, the superficial erosion in adulthood of Panx3KO mice may lead to reduced lubricin levels in aging, which could be chronically activating synovial lining cells, and thus producing the severity of synovitis we observed in the present study.
Inflammation is associated with age-related pathologies31including primary OA, and NF-κβ is a proposed central pathway of inflammation in OA.26Interestingly, through mechanotransduction pathways, chondrocytes can release ATP, and this extracellular ATP has been shown to activate NF-κβ signaling and contribute to OA.32^,^33While some ATP release is required to maintain normal cartilage homeostasis,34abnormal mechanical loading of cartilage increases chondrocyte ATP release.35^,^36This suggests that there are physiologically healthy levels of ATP release required for cartilage maintenance, but dysregulation of this mechanism could contribute to inflammation and OA. Our previous reports showed that aging WT mouse cartilage maintains similar PANX3 protein expression at 6, 18, and 24 mo of age,14suggestings PANX3 is required to maintain cartilage health well into aging. Deletion of Panx3may dysregulate this ATP signaling given its canonical function as a mechanosensitive, plasma membrane ATP release channel in cells such as chondrocytes,4^,^37but this role in ATP release remains as a hypothesis to be tested in future studies.
In our previous report, Panx3deletion did not significantly impact the progression of age-associated histopathological IDD in male mice at 18 and 24 mo of age compared to WT mice.6The present study also determined that the aged female Panx3KO mice IVD histopathological analysis matched that of males. The contrasting difference between the knee joint and IVDs of Panx3KO mice is interesting considering the similar mechanism of disease progression between OA and IDD.38It appears that PANX3 is not essential to IVD health during normal aging, as our previous report showed low gene expression of Panx3in IVDs from 6 to 24 mo of age relative to levels at 2 mo of age.6It is possible that PANX3 is utilized in early life and development of the IVD but is dispensable in aging. Interestingly, forced treadmill running seemed to have no effect on histological features of the IVDs regardless of genotype or sex. This was surprising, considering Panx3KO mice developed histopathological features in the AF of IDD with forced treadmill running in younger adult mice.23It was reasonable to hypothesize that aging would have rendered these mice more susceptible to forced treadmill running-induced changes to the IVD. However, considering aging alone results in relatively severe spontaneous IDD in mice,23any potential impact of forced treadmill running, positive or negative, may have been undetectable with histopathological scoring.
With all this in mind, we propose employing the aged Panx3KO mouse model utilized in this study as a novel model of primary knee OA. In contrast to many existing mouse models of spontaneous age-induced OA, which often exhibit high variability in disease incidence [39], this mouse model demonstrated a notable incidence rate of over 40% for severe OA. Moreover, the bimodal distribution of OARSI scores indicates that our Panx3KO model also displays low variation in disease severity. Additionally, this model only requires a single gene deletion and does not require the intervention of exercise protocols such as forced treadmill running, thereby reducing the technical demands associated with both its generation and maintenance. This minimization of demands also facilitates its integration with other models and interventions.
Future studies should involve a time course of Panx3KO mouse OA development. Specifically, analyzing earlier time points (eg, 6, 9, and 12 mo) to determine when distinct OA phenotypes arise, and to characterize its progression. This will allow for the analysis of the early cellular changes that may be driving this severe OA. Additionally, considering that obesity is a strong risk factor for OA, and coincides with aging,2high-fat diet studies in Panx3KO mice are also warranted. Lastly, tissue-specific KO models using various Cre driver lines are also warranted to determine the cell type responsible for this severe OA development.
Conclusion
Approximately half of aged male and female Panx3KO mice develop severe OA under both SED and FEX conditions, which occurs spontaneously as in human primary OA. Hence, we suggest the potential of the Panx3KO mice as a novel primary OA model. Additionally, Panx3KO mice have enthesitis of the quadriceps and distal patella more often than WT mice, but at the lumbar IVDs, Panx3KO mice have similar histological features as WT mice in aging. Collectively, our data suggests chronic suppression of PANX3 throughout life may be contraindicated and associated with severe OA. Future therapeutic interventions could include potential agonists of PANX3 to overcome the lower expression or reduced functionality of PANX3 channels in aged joint tissues.
Supplementary Material
SupplementaryFigures1-4_WakefieldTang_ziaf057
SupplementaryTables_WakefieldTang2025_ziaf057
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Cui A , Li H, Wang D, Zhong J, Chen Y, Lu H. Global, regional prevalence, incidence and risk factors of knee osteoarthritis in population-based studies. E Clinical Medicine. 2020;29-30:100587. 10.1016/j.eclinm.2020.10058734505846 PMC 7704420 · doi ↗ · pubmed ↗
- 2Johnson VL , Hunter DJ. The epidemiology of osteoarthritis. Best Pract Res Clin Rheumatol. 2014;28(1):5–15. 10.1016/j.berh.2014.01.00424792942 · doi ↗ · pubmed ↗
- 3Bond SR , Lau A, Penuela S, et al. Pannexin 3 is a novel target for Runx 2, expressed by osteoblasts and mature growth plate chondrocytes. J Bone Miner Res. 2011;26(12):2911–2922. 10.1002/jbmr.50921915903 · doi ↗ · pubmed ↗
- 4Iwamoto T , Nakamura T, Doyle A, et al. Pannexin 3 regulates intracellular ATP/c AMP levels and promotes chondrocyte differentiation. J Biol Chem. 2010;285(24):18948–18958. 10.1074/jbc.M 110.12702720404334 PMC 2881817 · doi ↗ · pubmed ↗
- 5Veras MA , Mc Cann M, Tenn NA, Séguin CA. Transcriptional profiling of the murine intervertebral disc and age-associated changes in the nucleus pulposus. Connect Tissue Res. 2020;61(1):63–81. 10.1080/03008207.2019.166503431597481 · doi ↗ · pubmed ↗
- 6Serjeant M , Moon PM, Quinonez D, Penuela S, Beier F, Séguin CA. The role of Panx 3 in age-associated and injury-induced intervertebral disc degeneration. Int J Mol Sci. 2021;22(3):1–15. 10.3390/ijms 22031080 PMC 786592933499145 · doi ↗ · pubmed ↗
- 7O'Donnell BL , Penuela S. Pannexin 3 channels in health and disease. Purinergic Signal. 2021;17(4):577–589. 10.1007/s 11302-021-09805-734250568 PMC 8677855 · doi ↗ · pubmed ↗
- 8Appleton CT , Pitelka V, Henry J, Beier F. Global analyses of gene expression in early experimental osteoarthritis. Arthritis Rheum. 2007;56(6):1854–1868.17530714 10.1002/art.22711 · doi ↗ · pubmed ↗