An Essential Adaptor for Apicoplast Fission and Inheritance in Malaria Parasites
James Blauwkamp, Krithika Rajaram, Sophia R. Staggers, Oliver Harrigan, Emma H. Doud, Sean T. Prigge, Stella Y. Sun, Sabrina Absalon

TL;DR
Researchers discovered a protein called PfAnchor that is crucial for the division and inheritance of the apicoplast in malaria parasites, offering a new target for antimalarial drugs.
Contribution
Identification of PfAnchor as the first apicoplast-specific dynamin adaptor protein regulating apicoplast fission in Plasmodium falciparum.
Findings
PfAnchor localizes to the apicoplast and is essential for its fission during parasite cell division.
Conditional depletion of PfAnchor disrupts apicoplast fission and leads to parasite death.
PfAnchor interacts with PfDyn2, a key mediator of organelle fission, establishing its role as a dynamin adaptor.
Abstract
Blood-stage Plasmodium falciparum parasites rely on a non-photosynthetic plastid, the apicoplast, for survival, making it an attractive target for antimalarial intervention. Like the mitochondrion, the apicoplast cannot be generated de novo and must be inherited by daughter parasites during cell division. This inheritance relies on coordinated apicoplast positioning and fission, but the molecular mechanisms controlling these processes remain poorly understood. Here, we identify a previously uncharacterized P. falciparum protein (Pf3D7_0613600), which we name PfAnchor, as a key regulator of apicoplast fission. Using Ultrastructure Expansion Microscopy (U-ExM), we show that PfAnchor localizes to the apicoplast throughout the asexual blood-stage. Conditional depletion disrupts apicoplast fission, leading to incomplete cytokinesis and parasite death. Notably, loss of the apicoplast’s…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
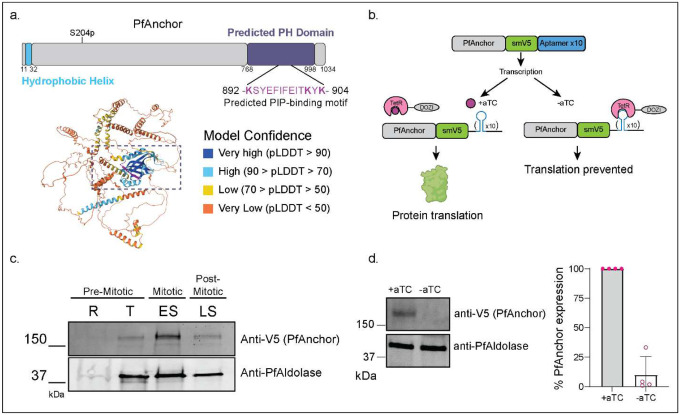 Figure 1
Figure 1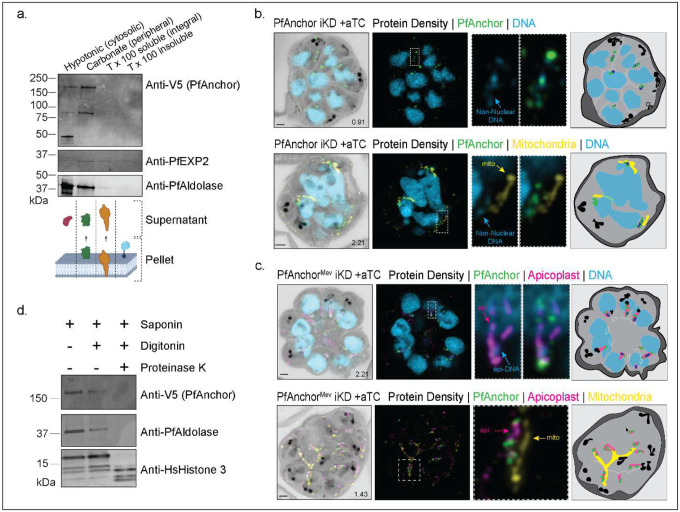 Figure 2
Figure 2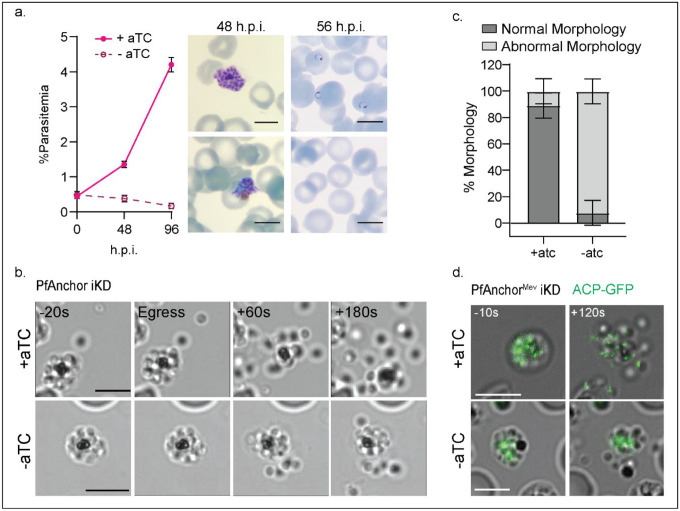 Figure 3
Figure 3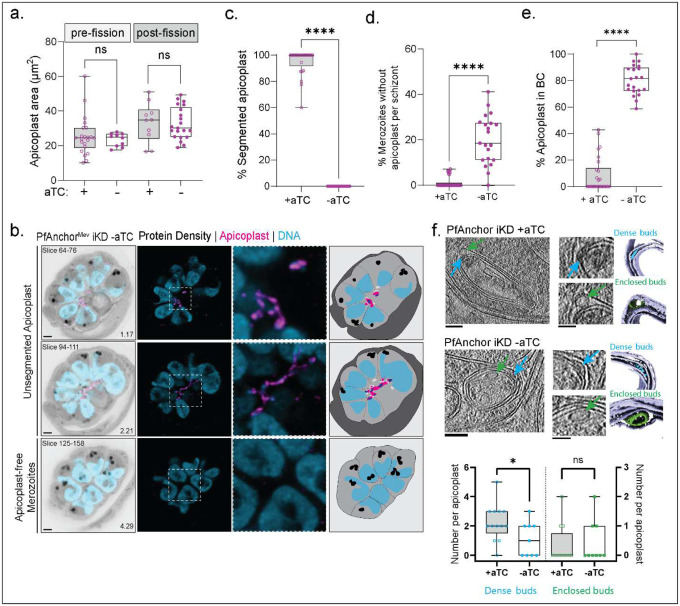 Figure 4
Figure 4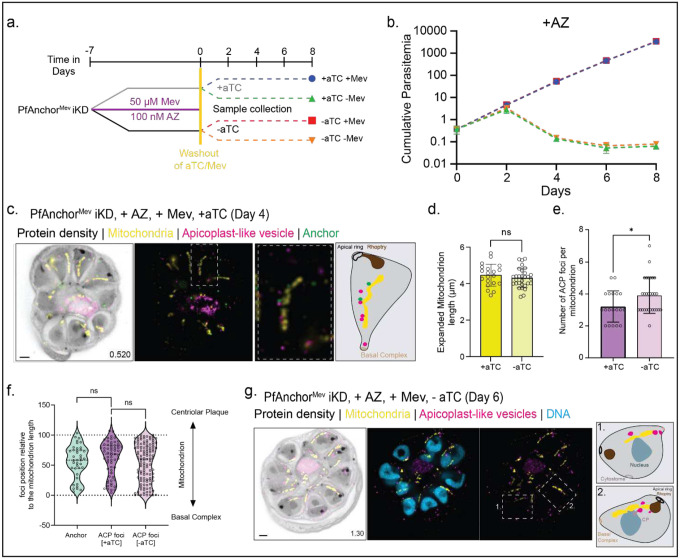 Figure 5
Figure 5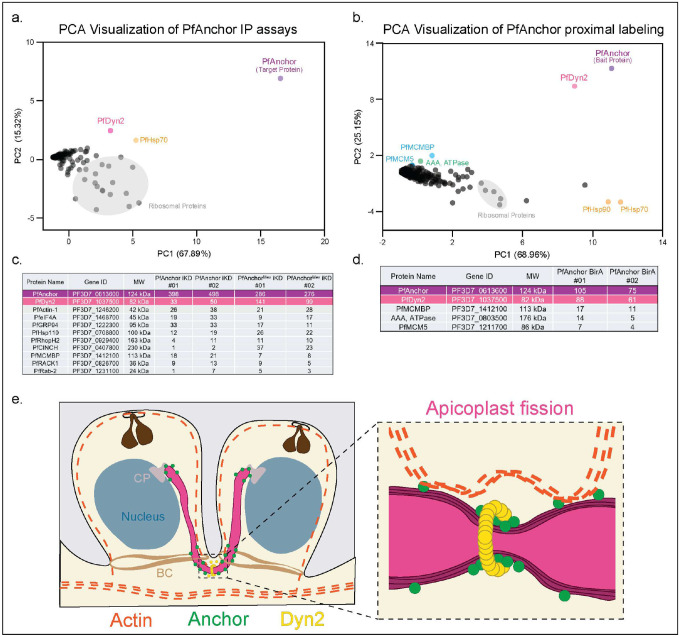 Figure 6
Figure 6Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsMalaria Research and Control · Mosquito-borne diseases and control · Vector-borne infectious diseases
Introduction
Protists, a diverse group of predominantly single-celled eukaryotes, represent the largest component of eukaryotic diversity and are found in nearly every environment [1, 2]. Like other eukaryotes, most protists contain mitochondria and/or plastid-derived organelles that support essential metabolic processes. Examples include photosynthetic plastids in dinoflagellates and the non-photosynthetic apicoplast found in most apicomplexan parasites [3]. Both mitochondria and plastids originate from endosymbiotic events where ancestral eukaryotes engulfed bacteria or algae that evolved into organelles [4]. Evidence for this evolutionary origin includes the presence of organelle genomes within mitochondria and plastids, as well as the multi-membrane structure of plastids derived from secondary endosymbiosis [5]. Given their fundamental roles in survival and metabolism and the absence of plastids in human hosts, these organelles represent attractive targets for novel antiparasitic drug development [6, 7].
Malaria is a parasitic disease caused by protozoan Plasmodium parasites, with an estimated 249 million cases and 608,000 deaths reported in 2022 [8]. Plasmodium parasites invade host red blood cells and undergo multiple rounds of asynchronous DNA replication within a shared cytoplasm, resulting in a multi-nucleated parasite [9]. These parasites contain a non-photosynthetic plastid known as the apicoplast. This metabolic organelle is thought to have originated from a secondary endosymbiosis event involving red algae and possesses its own reduced genome of approximately 35kb [10–12]. While the apicoplast plays a role in fatty acid and heme synthesis, studies have shown that its essential functions include the synthesis of isoprenoid precursors such as isopentenyl pyrophosphate (IPP) and coenzyme A (CoA) [13–19].
The apicoplast in Plasmodium is a single, branching organelle that expands in volume as the parasite progresses through the intraerythrocytic cycle. Volume electron microscopy (EM) studies, along with our recent work using ultrastructure expansion microscopy (U-ExM), have confirmed that the apicoplast undergoes multiple asynchronous fission events during schizogony, coinciding with parasite cytokinesis [20–22]. Recent studies have shown that apicoplast fission is mediated by a dynamin-like protein, PfDyn2 (Pf3D7_1037500), and is essential for parasite survival, as the apicoplast cannot be synthesized de novo [23, 24]. Unlike classical dynamin proteins, which contain a pleckstrin homology (PH) domain for direct membrane interaction during fission, dynamin-related proteins rely on adaptor proteins to recruit them to membranes and assemble functional fission complexes [25–27]. Mitochondrial fission adaptors vary widely in sequence and structure, with four identified in humans [28–31], three in yeast [32–34], two in the related parasite Toxoplasma gondii [35, 36], and one in Plasmodium [37]. However, no adaptor proteins specific to apicoplast fission have been identified in either T. gondii or P. falciparum, leaving the mechanism by which PfDyn2 facilitates apicoplast division unresolved.
In this study, we identify and characterize PfAnchor (Pf3D7_0613600), a nuclear-encoded protein that localizes to the apicoplast during the blood stage developmental cycle. Using the inducible TetR-DOZI conditional knockdown system [38], we demonstrate that PfAnchor is essential for parasite growth. Live microscopy of PfAnchor-deficient parasites revealed that while egress occurs, daughter cells form large clumps, preventing successful invasion. U-ExM analysis further showed that the apicoplast fails to undergo proper fission, resulting in most daughter parasites remaining tethered to each other via a collapsed apicoplast, while a subset of parasites emerges without an apicoplast. Blocking apicoplast branching via azithromycin treatment fully rescued the growth defect in PfAnchor-deficient parasites. Immunoprecipitation experiments identified an interaction between PfAnchor and PfDyn2, suggesting that PfAnchor acts as a dynamin adaptor protein required for apicoplast fission and inheritance. By uncovering a key regulator of apicoplast division, this study provides new insights into the molecular machinery governing organelle inheritance. Additionally, we highlight the potential of targeting apicoplast morphology as a novel strategy for inducing same-cycle parasite death, positioning PfAnchor as a promising therapeutic target for malaria intervention.
Results
PfAnchor: A conserved Plasmodium apicoplast-associated protein in the blood stage.
Proximal labeling experiments using P. falciparum Minichromosome Maintenance-binding protein (PfMCMBP) as bait [39] led to the identification of an uncharacterized protein that we named PfAnchor (Pf3D7_0613600, Figure S1-S2, Supp Data 1). PfAnchor is a Plasmodium specific 1034 amino acid protein. No homologs of PfAnchor could be identified in other apicomplexan organisms, suggesting it may represent a lineage-specific adaptation. The sequence structure contains a hydrophobic helix at the N-terminal end, predicted to be a transmembrane domain, as well as a potential phosphorylation site at S204 (Figure 1a). Alphafold3 structure prediction [40] revealed an ordered region at the C-terminal end which is present across Plasmodium species (Figure 1a, Figure S3). By combining Alphafold3 predictions with a DALI search [41], we, along with a recent publication [42], identified this region as a putative pleckstrin homology (PH) domain (Figure 1a). These domains are typically involved in membrane binding suggesting that PfAnchor may associate with a membrane. Sequence analysis of the predicted PH domain revealed strong conservation among Plasmodium species, with the highest similarity between P. falciparum and the Laverania subgenus (Figure S3a). Alphafold3 structure predictions across Plasmodium species with varying degrees of PfAnchor homology showed structural conservation of the predicted PH domain with high confidence model prediction in multiple species (Figure S3b). This structural conversation suggests that the predicted PH domain may have a conserved functional role in Plasmodium parasites.
Having identified PfAnchor as a potential membrane-associated protein through structural predictions, we next investigated its subcellular localization and membrane association. To enable detection and conditional regulation of PfAnchor, we engineered a C-terminal fusion of the protein with the spaghetti monster-V5 (smV5) epitope tag [43] and integrated the TetR-based conditional knockdown system [38] into the 3D7 parasite strain, named PfAnchor iKD (Figure 1b, Figure S4). This system allows for controlled expression of PfAnchor in the presence of anhydrotetracycline (aTC), a small molecule that stabilizes mRNA translation. In the absence of aTC, translation is suppressed, resulting in effective protein knockdown. Although PfAnchor’s transcriptional profile [44] suggests expression throughout the entire intraerythrocytic developmental cycle (IDC), we detected protein expression only from the trophozoite stage through segmented schizonts using western blot analysis (Figure 1c). To validate the functionality of the inducible knockdown system, we washed out aTC one cycle prior to analysis, approximately 10 hours before egress, and collected samples 40 hours post-invasion. Western blot analysis showed that PfAnchor expression was reduced by more than 90% across four independent biological replicates, confirming the efficacy of the knockdown system (Figure 1d).
To determine whether PfAnchor is membrane-associated, we conducted sodium carbonate solubility assays. Parasites were hypotonically lysed to remove soluble cytosolic proteins, followed by sequential extraction with sodium carbonate (to release peripherally associated membrane proteins) and Triton-X100 (to solubilize integral membrane proteins). The resulting fractions, including the supernatant and pellet, were analyzed by western blot to assess the distribution of PfAnchor. Although minor cytosolic contamination was detected in the sodium carbonate fraction, most of PfAnchor was enriched in this fraction, consistent with a peripheral association with membranes (Figure 2a).
Given its membrane association, we next sought to identify the specific membrane compartment with which PfAnchor associates. Using U-ExM, we observed that PfAnchor localizes to regions containing non-nuclear DNA, suggesting a potential association with a DNA-containing organelle. By staining PfAnchor parasites with MitoTracker, we showed that PfAnchor did not co-localize with the mitochondrion (Figure 2b). As the apicoplast possesses its own genome, we hypothesized that PfAnchor might localize to the apicoplast during this stage. To test this, we generated a new cell line by tagging PfAnchor with smV5 in the PfMev::Api-GFP background [45] that we named PfAnchor^Mev^ iKD (Figure S5). This strain enables conditional disruption of the apicoplast by mevalonate (MVA) supplementation, supporting viability of the parasite without the organelle. This system facilitates visualization of the apicoplast through GFP tagging and allows investigation of PfAnchor’s function in the absence of an intact apicoplast. Imaging analysis of this cell line revealed that PfAnchor localizes to the periphery of the apicoplast throughout schizogony (Figure 2b, c), supporting its involvement in apicoplast biology.
To determine the orientation of PfAnchor within the apicoplast, we performed proteinase K protection assays. Infected red blood cells were lysed with saponin to release parasites, followed by treatment with digitonin to selectively permeabilize the parasite plasma membrane. The resulting pellet was exposed to proteinase K to degrade cytosolic-facing proteins while preserving proteins within organellar compartments. We found that PfAnchor was susceptible to proteinase K digestion, indicating that its C-terminal epitope tag faces the cytosol (Figure 2d). Together, these findings demonstrate that PfAnchor is a cytosol-facing protein associated with the apicoplast membrane.
PfAnchor is essential for apicoplast partitioning and merozoite formation.
To investigate the role of PfAnchor in parasite development, we removed anhydrotetracycline (aTC) from the culture and monitored parasite growth by microscopy. PfAnchor-deficient parasites exhibited a complete growth arrest during the first cycle of depletion, with no early-stage parasites detected in the culture, suggesting a critical role in either egress or invasion (Figure 3a). To test this, we performed live microscopy on PfAnchor-expressing and PfAnchor-deficient parasites to investigate whether egress was affected. Parasites were treated with Compound 1 to arrest and synchronize them just prior to egress from red blood cells [46]. After washing with fresh media, egress events were monitored. PfAnchor-expressing parasites displayed rapid and explosive egress, with individual daughter merozoites dispersing quickly. By contrast, PfAnchor-deficient parasites underwent egress, yet the majority of daughter merozoites remained clustered and failed to fully separate, often remaining attached near the food vacuole (Figure 3b, c). To quantify egress phenotypes, we categorized parasites based on morphology: “normal” morphology was defined as minimal merozoite clumping with daughter parasites dispersing effectively, while “abnormal” morphology was characterized by limited separation of daughter parasites, which formed slow-moving or immobile clumps. PfAnchor-expressing parasites showed an average of 89.3 +/− 9.5% normal morphology (52/61 egress events) over three biological replicates while PfAnchor-deficient parasites showed an average of 10.7 +/− 9.3% normal morphology (3/39 egress events) over three biological replicates. We note that PfAnchor-deficient parasite morphology closely resembles the phenotype associated with impaired apicoplast segregation [47]. Given this resemblance, we hypothesized that disrupted apicoplast branching may underline the observed cytokinesis defect. To test this, we visualized the apicoplast morphology in PfAnchor-deficient parasites expressing ACP-GFP (PfAnchor^Mev^ iKD parasites). Imaging of PfAnchor deficient parasites revealed two distinct groups of merozoites: clumped merozoites that share a single, unsegregated apicoplast, and free merozoites that display no apicoplast signal. We hypothesize that these free merozoites lack an apicoplast. In contrast, in PfAnchor-expressing parasites exhibit proper segregation, with each merozoite containing a distinct apicoplast (Figure 3d). These findings demonstrate that PfAnchor is essential for apicoplast inheritance during merozoite formation.
PfAnchor is required for apicoplast division during IDC.
Given its localization to the apicoplast periphery and the observed defects in organelle inheritance upon knockdown, we reasoned that PfAnchor plays a direct role in apicoplast division. We hypothesize that in the absence of PfAnchor, apicoplast fission fails, resulting in merozoites tethered by a single, undivided apicoplast. To test this, we used U-ExM to track apicoplast division during parasite replication and determine how PfAnchor depletion disrupts organelle division and inheritance. To assess whether PfAnchor loss affects apicoplast biogenesis, we measured apicoplast area in cell projections [21]. No significant differences were observed between PfAnchor-expressing and deficient parasites, indicating that apicoplast biogenesis remains unaffected (Figure 4a). Additionally, as we and others previously demonstrated, the apicoplast adopts a characteristic crown shape during early segmentation, with a single branched apicoplast connecting all outer centriolar plaques (CPs) [21, 48, 49]. This organization was also observed in PfAnchor-deficient parasites, indicating that the pre-fission positioning of the apicoplast and its association with the CPs remain unaffected in the absence of PfAnchor (Figure S6). However, PfAnchor-deficient parasites exhibited a striking defect in apicoplast inheritance during cytokinesis. Unlike control parasites, which display individual, segmented apicoplasts in daughter parasites, PfAnchor-deficient parasites retained a single, interconnected apicoplast linking multiple daughter parasites (Figure 4b, c). Live microscopy previously revealed a subset of daughter parasites lacking apicoplast signal, and U-ExM further confirmed this phenotype, showing that these apicoplast-negative merozoites had separated from the cluster and moved freely (Figure 4d). This resulted in two distinct populations of daughter parasites in the PfAnchor-deficient samples: (1) parasites connected by a shared apicoplast traversing the basal complex (Figure 4e), and (2) parasites that were physically separated from the residual body but devoid of apicoplast signal (Figure 4d). Notably, while PfAnchor-deficient parasites displayed severe apicoplast inheritance defects, mitochondria biogenesis and division remained unaffected, with each daughter cell inheriting a single mitochondrion (Figure S7). These findings establish PfAnchor as a critical factor of apicoplast division and inheritance, and its loss likely prevents parasite propagation and survival.
To further investigate the impact of Anchor depletion on apicoplast structure, we performed cryo-focused ion beam (cryo-FIB) milling and cryo-electron tomography (cryo-ET) on PfAnchor iKD parasites in the presence and absence of aTC. To ensure stage-matched analysis, we treated cultures with Compound 1 (4-[2-(4-fluorophenyl)-5(1-methylpiperidine-4-yl)-1H-pyrrol-3-yl]pyridine or C1), a reversible protein kinase G inhibitor that arrests schizonts immediately prior to egress by preventing parasitophorous vacuole membrane (PVM) rupture [46, 50]. C1-stalled schizonts were Percoll-purified and incubated in media containing 2.5 μM C1 for 2–3 hours before fixation. We analyzed a total of 12 tomograms from PfAnchor-expressing (+aTC) parasites, all containing identifiable apicoplasts, and 9 tomograms from PfAnchor-deficient (–aTC) parasites that featured apicoplasts. The apicoplast was distinguished in tomograms based on its characteristic four-membrane structure. 2D Measurement of the smallest internal diameter, taken from a 2D slice at the center of the apicoplast volume, revealed no significant difference between PfAnchor-expressing and PfAnchor-deficient parasites (Figure S8a). In both PfAnchor-expressing and PfAnchor-deficient parasites, we observed cryo-EM densities within the intermembrane space of the apicoplast (Figure 4f). These densities included dense intermembrane buds, defined as electron-dense tethering structures between neighboring apicoplast membranes (Figure 4f, blue arrows). Interestingly, the frequency of these dense intermembrane buds was significantly reduced in PfAnchor-deficient parasites, with PfAnchor-expressing parasites present ~2.2 dense buds per apicoplast on average (S.D. = 1.2), while PfAnchor-deficient parasites averaged ~1.1 dense buds per apicoplast (S.D. = 1.2).
In addition to the dense intermembrane buds, we identified enclosed membrane-like buds—small, circular densities forming between apicoplast membranes (Figure 4f, green arrows). Unlike the dense intermembrane buds, the number of enclosed buds remained unchanged between PfAnchor-expressing and PfAnchor-deficient parasites. This suggests that while PfAnchor influences the formation or maintenance of dense intermembrane buds, it does not significantly impact the presence of enclosed buds, indicating that some aspects of membrane organization remain intact. Beyond these structures, we observed additional internal features within the apicoplast lumen. These included single-layer and double-layer membrane structures, which may represent distinct internal compartments or intermediates in apicoplast remodeling (Figure S8b-e). Quantification of single-layer membrane structures revealed no significant differences in their abundance or internal diameter between PfAnchor-expressing and PfAnchor-deficient parasites (Figure S8c, d), suggesting that their formation is independent of PfAnchor. However, double-layer membrane structures were exclusively detected in PfAnchor-deficient parasites (Figure S8e), raising the possibility that PfAnchor depletion alters the internal architecture of the apicoplast. These findings suggest that while PfAnchor is not required for overall membrane biogenesis, it may play a role in maintaining the organization or remodeling of apicoplast membrane organization, potentially influencing its division and inheritance.
Loss of apicoplast-branched structure rescues growth in PfAnchor-deficient parasites.
We hypothesized that the egress defects observed in PfAnchor-deficient parasites result from physical tethering of daughter merozoites by the branched apicoplast structure and that disrupting this tethering could restore parasite viability. Bacterial translation inhibitors such as doxycycline, clindamycin, and azithromycin are known to disrupt apicoplast structure, leading to the formation of apicoplast-like vesicles during the second cycle of treatment and parasite death [13, 45]. The growth of parasites treated with bacterial translation inhibitors can be rescued with the addition of exogenous IPP [13], leading to parasites growing with apicoplast-like vesicles instead of a branching apicoplast and irreversible loss of the apicoplast genome [51]. The high cost and the difficulties associated with the IPP-rescue system [13] for long-term studies led to the development of an apicoplast bypass system by Swift et al. in 2020 [45]. This system introduces four enzymes into the cytoplasm of the parasite, allowing for IPP synthesis via the mevalonate (MVA) pathway in the cytoplasm, thereby bypassing the endogenous methylerythritol phosphate (MEP) pathway in the apicoplast. By adding mevalonate to the growth medium, the bypass system significantly reduces the cost and improves the feasibility of long-term experimental studies.
To test our hypothesis, we used the apicoplast bypass PfAnchor^Mev^ iKD cell line and treated parasites with the antibiotic azithromycin (AZ) to prevent apicoplast branching during PfAnchor knockdown [13, 52]. Briefly, PfAnchor^Mev^ iKD parasites were initially cultured in the presence of 50 μM mevalonate (Mev) and 100 nM AZ for 7 days. The parasites were then split into four experimental groups: (1) +aTC -Mev (green), (2) -aTC -Mev (orange), (3) +aTC +Mev (blue), and (4) -aTC +Mev (red) (Figure 5a). These groups were cultured for an additional 8 days, taking samples every 2 days for growth analysis. To prevent culture collapse, parasite cultures were diluted (1:8) every 2 days. U-ExM validated the loss of the apicoplast branching structure and PCR analysis confirmed the loss of the apicoplast genome regardless of the presence of PfAnchor (Figure 5c, g, Figure S9).
Growth analysis revealed distinct phenotypes depending on PfAnchor expression and the presence of Mev. In parasites with apicoplast-like vesicles (+AZ), parasites supplemented with Mev displayed equivalent growth rates regardless of PfAnchor expression (blue and red lines overlapping, Figure 5b), indicating that disruption of the branched apicoplast structure fully rescued the growth defects associated with PfAnchor depletion. Conversely, parasites with apicoplast-like vesicles but not supplemented with Mev failed to grow regardless of PfAnchor expression (green and orange lines overlapping, Figure 5b), validating the experimental setup. These results demonstrate that the loss of the branched apicoplast structure is sufficient to fully rescue the growth defects in PfAnchor-deficient parasites. This finding underscores the critical role of PfAnchor in apicoplast fission. Since parasite viability is restored in the absence of a branched apicoplast, it appears unlikely that PfAnchor has any other essential functions beyond facilitating apicoplast fission.
As controls we tested the growth of parasites with a branched apicoplast (-AZ) in the presence or absence of Mev or PfAnchor. Branched apicoplast PfAnchor-expressing parasites showed robust growth with or without Mev supplementation (green and blue lines overlapping, Figure S10). Branched apicoplast PfAnchor-deficient parasites failed to grow without Mev, as evidenced by a decline in parasitemia (orange line, Figure S10b). Interestingly, PfAnchor-deficient parasites supplemented with Mev initially exhibited reduced growth but subsequently recovered, achieving growth rates comparable to PfAnchor-expressing parasites (red line, Figure S10b). We hypothesize that this initial recovery of growth is caused by free merozoites which contain no apicoplast in the PfAnchor population (Figures 3d, 4b) that can survive and propagate when supplemented with Mev.
Apicoplast-like vesicles are retained in daughter parasites in association with mitochondria, independent of PfAnchor.
To investigate the fate of the apicoplast following its disruption by azithromycin, we examined the spatial organization and inheritance of the resulting apicoplast-like vesicles using U-ExM (Figure 5c, g). In PfAnchor-expressing parasites, the majority of apicoplast-like vesicles were excluded from the daughter parasites and instead localized within the residual body at the completion of cytokinesis (Figure S11). However, a subset of apicoplast-like vesicles remained within daughter cells, and these retained vesicles were consistently associated with the mitochondrion (Figure 5c, S11). This suggests that mitochondrial association plays a key role in apicoplast vesicle inheritance, while vesicles not tethered to the mitochondrion are largely excluded from daughter parasites during cytokinesis.
To determine whether PfAnchor is required for apicoplast-like vesicle inheritance, we examined PfAnchor-deficient parasites and found that their vesicle localization and distribution were largely similar to PfAnchor-expressing parasites (Figure 5g, S12). However, quantification revealed a modest but significant increase in the number of mitochondrion-associated apicoplast vesicles in PfAnchor-deficient parasites compared to PfAnchor-expressing parasites (Figure 5e). This suggests that while PfAnchor is not required for vesicle retention, its depletion may subtly influence the balance between vesicle inheritance and clearance. Despite this increase, the spatial distribution of apicoplast-like vesicles along the mitochondrion was unchanged between conditions (Figure 5f), reinforcing the conclusion that PfAnchor does not impact the positioning of retained vesicles. As the mitochondrion served as a spatial reference for apicoplast vesicle distribution, we quantified mitochondrial length to ensure that mitochondrial organization remained unchanged in the presence or absence of PfAnchor. No significant differences in mitochondrial length were observed between PfAnchor-expressing and PfAnchor-deficient parasites following azithromycin treatment (Figure 5d), confirming that any differences in apicoplast-like vesicle localization would not be due to changes in mitochondrial morphology. Together, these data indicate that mitochondrial association is a key determinant of apicoplast vesicle retention, while PfAnchor is not required for their inheritance.
PfAnchor interacts with PfDyn2 to facilitate apicoplast fission.
Knockdown of PfAnchor disrupts apicoplast segregation and positioning, yet its precise molecular function remains unclear. To identify potential interacting partners, we performed immunoprecipitation (IP) experiments using two PfAnchor-tagged cell lines (PfAnchor iKD and PfAnchor^Mev^ iKD), followed by unbiased mass spectrometry. Parental untagged (3D7-Cas9) parasites served as a control to distinguish specific interactors from background proteins. Principal component analysis (PCA) of the mass spectrometry data confirmed the specificity of PfAnchor-associated interactions, as PfAnchor clustered distinctly from background interactors (Figure 6a, Supp Data 2). Among the most highly enriched proteins, PfDyn2 emerged as a primary interactor, supporting its role as a key regulator of apicoplast and mitochondrial fission [23, 24]. In addition to PfDyn2, PCA identified PfHsp70 (PF3D7_0818900) as a secondary interactor, suggesting a possible role in stabilizing protein complexes during apicoplast division [53, 54]. Beyond the PCA analysis, PfAnchor pulldown experiments identified additional interactors uniquely present in the PfAnchor dataset (Figure 6c). Among these, PfActin1 was specifically enriched, consistent with its established role in cytoskeletal regulation of organelle segregation [47, 55]. PfCinch, a basal complex protein [56], was also identified, suggesting a possible link between PfAnchor and cytoskeletal structures involved in organelle positioning. These findings provide strong evidence that PfAnchor interacts with PfDyn2, reinforcing its involvement in apicoplast fission. The identification of PfHsp70 as a secondary interactor suggests a potential chaperone function that may contribute to the stability of the fission complex. Additionally, the enrichment of PfActin1 and PfCinch supports a possible cytoskeletal role in apicoplast segregation.
To complement our IP approach and further define the PfAnchor interactome, we performed proximity-dependent biotin labeling (BioID), followed by mass spectrometry. BioID enables the identification of proteins in close spatial proximity to PfAnchor, including transient or membrane-associated interactors. Two independent biological replicates of BioID with PfAnchor-BirA-HA(Figure S13) were performed, with parental untagged parasites serving as a control to account for background labeling. PCA analysis of BioID mass spectrometry data demonstrated that PfAnchor clustered distinctly from background interactors, with PfDyn2 emerging as the most enriched protein in proximity to PfAnchor (Figure 6b, Supp Data 3). This further supports a functional association between PfAnchor and PfDyn2, consistent with the results of our immunoprecipitation experiments. Additional proteins detected in the BioID dataset included components of the MCM complex, though their potential relevance to apicoplast function remains unclear. PfDyn2 was consistently enriched in both BioID and IP datasets, reinforcing its role as PfAnchor’s primary interactor (Figure 6d). This strong association suggests that PfAnchor may act as an adaptor or regulator in apicoplast fission, potentially recruiting PfDyn2 to the organelle membrane. Together, these findings support a model in which PfAnchor serves as a key adaptor protein, bridging cytoskeletal and membrane remodeling factors to facilitate apicoplast fission (Figure 6e).
Discussion
This study reveals a fundamentally new mechanism of parasite death and identifies PfAnchor as an essential adaptor that enables apicoplast fission in Plasmodium falciparum. We show that PfAnchor localizes to the apicoplast throughout the intraerythrocytic cycle and is required for its division and inheritance. Conditional depletion of PfAnchor disrupts apicoplast fission, resulting in physically tethered merozoites that fail to complete cytokinesis and cannot invade new red blood cells. This phenotype demonstrates, for the first time, that failure to divide the apicoplast can directly impair cell division and lead to same-cycle parasite death. Additionally, we highlight the potential of targeting apicoplast morphology as a novel strategy for inducing same-cycle parasite death, positioning PfAnchor as a promising therapeutic target for malaria intervention.
PfAnchor’s Structural Features and Functional Implications
Alphafold3 predictions and domain analysis suggest that PfAnchor contains a pleckstrin homology (PH) domain, a common motif for membrane binding. Given that the apicoplast membrane is enriched in PI3P [57, 58], it is plausible that PfAnchor’s PH domain mediates its localization. Additionally, PfAnchor’s N-terminal region contains a hydrophobic helix, which carries a mutation linked to resistance against actin-polymerization inhibitors [59]. The co-occurrence of mutations in PfAnchor and profilin (a key actin regulator) in parasites resistant to MMV020291, an antimalarial that targets actin polymerization, suggests a functional link between PfAnchor and actin dynamics [59]. Actin dynamics are increasingly recognized as important for apicoplast segregation, and our identification of PfActin1 as a PfAnchor interactor supports this idea [47, 55]. In other eukaryotic systems, profilin-actin interactions regulate fission events by remodeling membrane-associated cytoskeletal networks [60–62]. By analogy, PfAnchor may facilitate actin polymerization at the apicoplast fission site, potentially through indirect interactions with profilin. Future studies should investigate whether targeted disruption of PfAnchor’s hydrophobic helix affects actin-mediated apicoplast segregation.
PfAnchor’s Role in Organelle Structure and Function
Despite its role in apicoplast fission, PfAnchor is not required for overall apicoplast biogenesis. Parasites lacking PfAnchor remain viable when the metabolic function of the apicoplast is bypassed using mevalonate supplementation. Furthermore, treatment with azithromycin, which eliminates the branched apicoplast structure, rescued the growth defects of PfAnchor-deficient parasites. These findings confirm that PfAnchor’s primary function is in organelle fission rather than metabolism. Interestingly, while PfAnchor is not required for other apicoplast functions such as fatty acid synthesis, iron-sulfur cluster biosynthesis, or heme production, its potential role in isoprenoid precursor synthesis remains unclear. Inhibition of the MEP pathway using fosmidomycin results in an unbranched, stagnant apicoplast [13, 14]; however, PfAnchor-deficient parasites still exhibit normal biogenesis and branching, making a direct role in isoprenoid metabolism unlikely.
Cryo-electron tomography of PfAnchor-deficient and control parasites further revealed the presence of distinct electron-dense structures within the apicoplast. We identified two main morphologies: “dense buds,” small, localized densities between membranes, and “enclosed buds,” larger invaginated membrane profiles. Similar membranous profiles—sometimes described as tubular whorls—have been reported in apicoplasts of Plasmodium and Toxoplasma, though their functional role remains unresolved [63–65]. Comparative plastid systems such as etioplasts, also contain analogous intra-organelle structures, including protein complexes, plastoglobuli, and tubular arrays involved in membrane remodeling and metabolic scaffolding [66]. The persistence of these features in the Plasmodium apicoplast, independent of PfAnchor expression, raises intriguing questions about their function in membrane remodeling, metabolite transport, or structural maintenance of the apicoplast’s four-membrane architecture. Whether these intramembrane structures persist within the apicoplast-like vesicles remains an open question. Further investigation will be required to determine whether these features are involved in apicoplast homeostasis, division, or are vestiges of its photosynthetic ancestry.
Mitochondrion-Apicoplast Tethers and Organelle Inheritance
Our findings provide the first high-resolution evidence that mitochondrion-associated apicoplast-like vesicles are selectively inherited [67], reinforcing the functional dependency between these organelles [49, 65]. While most vesicles were excluded from daughter cells, the retained subset consistently colocalized with mitochondria (Figures 5, S10, S11). This strong spatial association suggests a role for physical tethering between the apicoplast and mitochondrion in ensuring faithful organelle inheritance. Moreover, the molecular components mediating this tethering are likely preserved on apicoplast-like vesicles that are inherited. Consistent with this idea, electron-dense structures at the interface between the mitochondrion and apicoplast have recently been described by cryo-ET in P. falciparum, suggesting the presence of defined tethering complexes at points of close membrane contact [65]. While we did not specifically image vesicles in contact with mitochondria by cryo-ET in this study, it remains an open and important question whether these dense tethering structures persist in the apicoplast-like vesicles that are inherited following AZ treatment. Interestingly, PfAnchor depletion resulted in a slight but significant increase in mitochondrion-associated vesicles (Figure 5e), indicating that while PfAnchor is not required for inheritance, its loss may alter the balance between vesicle retention and clearance. The nature of the mitochondrion–apicoplast tethers remains to be elucidated, but it is likely mediated by tethers that help coordinate spatial organization during division. Are these tethers simple structural linkages, or true membrane contact sites that enable metabolite exchange or signaling? The role of the cytoskeleton, particularly actin, in stabilizing these tethers warrants further investigation, as cytoskeletal elements are known to contribute to apicoplast segregation in Plasmodium and other apicomplexans[47, 55].
PfAnchor Interacts with PfDyn2 and the Cytoskeleton
To elucidate the molecular function of PfAnchor, we performed immunoprecipitation and proximity labeling experiments, which identified PfDyn2 as its primary interacting partner. PfDyn2 is a dynamin-related GTPase known to mediate apicoplast and mitochondrial fission [23, 24]. The enrichment of PfDyn2 in both datasets supports a model in which PfAnchor functions as a dynamin adaptor, likely recruiting PfDyn2 to the apicoplast membrane to facilitate membrane fission. This is analogous to mitochondrial fission adaptors in other eukaryotes, which tether dynamin-related proteins to fission sites. Additionally, we identified interactions between PfAnchor and cytoskeletal components, including PfActin1 and PfCINCH, suggesting a potential role in coordinating apicoplast fission with parasite cytoskeletal dynamics [55, 56]. Notably, the basal complex, which facilitates cytokinesis, may also coordinate organelle fission. Positioned at the parasite plasma membrane, it could serve as a platform for synchronizing apicoplast and mitochondrial division with daughter cell separation. Future studies should examine whether basal complex components, such as PfCINCH, actively remodel membranes or anchor fission machinery at the apicoplast and mitochondria during schizogony. Additionally, PfHsp70 (PF3D7_0818900), identified in our interactome analysis, could stabilize the fission complex, as its homologs in other eukaryotes support mitochondrial and chloroplast division by assisting in protein complex assembly and function [54, 68].
Finally, we identified PfAnchor via proximal labeling using PfMCMBP, a DNA helicase that functions during DNA replication [39]. While reciprocal proximity labeling with PfAnchor as bait confirmed the presence of PfMCMBP and PfMCM5, our immunoprecipitation experiments did not recover these proteins, suggesting that they are not direct binding partners of PfAnchor. Instead, their detection in proximity labeling assays likely reflects their spatial localization within the parasite during schizogony rather than a functional interaction. The apicoplast and PfAnchor are positioned near the centriolar plaque (CP) immediately before segmentation (Figure S6), and the CP is surrounded by nuclear pore complexes, indicating active nucleocytoplasmic transport in this region [69]. Given this, it is plausible that the MCM complex, which shuttles between the cytoplasm and nucleus during DNA replication, transiently occupies a similar cellular space. While a direct functional relationship between PfAnchor and the MCM complex remains unlikely, these findings highlight the complex organization of cellular structures during P. falciparum blood-stage parasites replication and underscore the utility of proximity labeling in capturing transiently colocalized proteins within the parasite.
PfAnchor in Other Developmental Stages
Although our study establishes PfAnchor’s role in blood-stage parasites, its expression in other life cycle stages suggests broader functions. Transcriptomic data indicate that PfAnchor is expressed during gametocytogenesis, where male and female gametocytes harbor a single, non-branching apicoplast [70, 71]. Whether PfAnchor plays a role in gametocyte development or is dispensable at this stage remains unclear. By contrast, during mosquito and liver stages, parasites undergo extensive replication, and their apicoplasts adopt a branching tubular morphology similar to blood stages [72]. This implies that apicoplast fission must occur to ensure organelle inheritance during liver merozoite and sporozoite formation. Investigating PfAnchor’s function in these stages could reveal conserved mechanisms of apicoplast division throughout the parasite life cycle and identify new transmission-blocking targets.
Our findings identify PfAnchor as an essential adaptor linking dynamin-mediated membrane scission to actin-dependent apicoplast partitioning. In many eukaryotes, the same dynamin-related protein (such as Yeast Dnm1 or Human Drp1) is recruited to different organelles via organelle-specific adaptors, enabling functional specificity despite shared GTPase machinery [25, 26]. Similarly, in Plasmodium, the dynamin-related protein Dyn2 mediates fission of both the mitochondrion and the apicoplast (PMID: 39611847). Here, we demonstrate that PfAnchor is specifically required for apicoplast division but dispensable for mitochondrial fission, revealing the first known organelle-specific adaptor for dynamin function in this system. The adaptor responsible for mitochondrial fission in Plasmodium remains to be identified. The selective requirement for PfAnchor in apicoplast division may reflect the evolutionary origin and complex membrane architecture of this organelle, which is derived from a red algal endosymbiont and surrounded by four membranes. Importantly, studying Plasmodium and other myxozoans—including dinoflagellates—offers a unique opportunity to explore how a single dynamin pathway is adapted to remodel and divide organelles of diverse evolutionary origins within the same cellular environment. While the apicoplast in Apicomplexa has evolved into a non-photosynthetic, metabolism-focused organelle essential for parasitism, dinoflagellates retain photosynthetic plastids derived from the same secondary endosymbiotic event. This divergence provides a powerful comparative framework to dissect how conserved membrane fission machinery is repurposed across lineages to accommodate organelle-specific architectures and functions. Given that apicoplast division is essential for parasite survival and lacks a counterpart in human cells, targeting PfAnchor or its associated machinery could offer a promising new avenue for antimalarial strategies. Future studies will be critical to uncover the molecular composition and regulation of this fission pathway and to understand how cytoskeletal remodeling and membrane scission are coordinated to ensure faithful inheritance of the apicoplast.
Materials and Methods
Parasite culture:
The 3D7 Plasmodium falciparum strain, obtained from the Walter and Eliza Hall Institute (Melbourne, Australia) was cultured in human O^+^ RBCs at 4% hematocrit in RPMI-1640 containing 25 mM HEPES, 50 mg/L hypoxanthine, 0.21% sodium bicarbonate, and 0.5% w/v Albumax II [73]. Cultures were incubated at 37 °C while shaking in a gas mixture of 1% O_2_, 5% CO_2_, and 94% N_2_ as previously described The PfAnchor iKD (smV5-Tet Pf3D7_0613600) and PfAnchor^Mev^ iKD (smV5-Tet Pf3D7_0613600 in PfMev background) cell lines were maintained under selection of 2.5 nM WR99210 and supplemented with 500 nM anhydrotetracycline (aTC).
Plasmid generation and transfection:
To generate the PfMCMBP-BirA-HA cell line for proximal labeling, the homology region of pSAB60 (PfMCMBP-3HADD [39]) was cut with Not1/Xho1 and cloned into our BirA plasmid (pRR28) [74] to create pSAB99. To generate the PfMCMBP-BirA-HA strain, 100 μg of pSAB99 plasmid was transfected into synchronized ring staged parasites using electroporation. Upon transfection, stable single crossover parasites were selected by cycling 2.5 nM WR99210 on and off as previously described [75]. Integration of pSAB99 plasmid into the genome of 3D7 was confirmed via PCR using primer pairs oSAB504/oJDD44, oSAB505/oJDD44, and oSAB504/oSAB506 (Figure S1).
To create the Pf3D7_0613600-smV5-Tet (PfAnchor iKD) plasmid, the Pf3D7_0613600 5’ and 3’ homology regions were PCR amplified from 3D7 genomic DNA with oligonucleotides oJDD4308/4310 and oJDD4307/4309 respectively. The two pieces were fused together using Sequence Overlap Extension PCR (SOE-PCR) using oJDD4803/4309 and the piece was digested using Not1/Nco1 and ligated onto pRR92 [56] (which contains the smV5 epitope tag, ten copies of the Tet-aptamer for the TetR-DOZI aptamer knockdown system, and the expression cassette for human dihydrofolate reductase (hDHFR)) to form pSAB177. To create Pf3D7_0613600 targeting guide RNA plasmid, oJDD4347/4348 were annealed, phosphorylated, and ligated into BpiI-digested pRR216 [23] to generate pSAB175. All oligonucleotide sequences are shown in Table S1. To generate the PfAnchor iKD strain, 100 μg of pSAB177 plasmid was linearized with Stu1 and transfected into 3D7-Cas9 parasites, along with 100 μg of pSAB175 and supplemented with 500 nM aTC. 24 hours following transfection parasites were treated with 5 nM WR99210 for 7 days before maintaining on 2.5 nM WR99210 and 500 nM aTC until resistant parasites were detected. Integration of pSAB177 plasmid into the genome of 3D7 was confirmed via PCR using primer pairs oSAB243/oJDD2933, oSAB367/oSAB2933, oSAB243/oSAB366, and oSAB367/oSAB366. Primer binding locations and predicted PCR product sizes are listed in Figure S4a-b.
To generate the PfAnchor^Mev^ iKD strain, 350 μL of RBCs were electroporated using the GenePulser XCell system (Bio-Rad) with 75 μg each of pSAB175 and linearized pSAB177 obtained by digestion with EcoRV. The electroporated RBCs were combined with 1 mL of PfMev^attB^ parasite culture at 3% parasitemia and cultured in 10 mL of CMA (Complete Medium with Albumax) for two days. Subsequently, the cultures were transferred to selective medium containing 2.5 nM WR99210 for seven days. Afterward, the cultures were returned to CMA until parasites were detected on blood smears, at which point WR99210 was reintroduced to the medium. The transgenic parasite culture was cloned by limiting dilution. Clones were screened for an intact aptamer array via PCR amplification using primers oSAB249 and oJDD44. Clone F3 was selected for further experiments (Figure S14). Integration of pSAB177 plasmid into the genome of NF54-PfMev parasites [45] was confirmed via PCR using primer pairs oSAB243/oJDD2933, oSAB367/oSAB2933, oSAB243/oSAB366, and oSAB367/oSAB366. Primer binding locations and predicted PCR product size found in Figure S4a, c.
To generate the PfAnchor-BirA-HA proximal labeling plasmid, homology region from pSAB177 was amplified using oSAB356/358 and the piece was digested using Not1/Xho1 and ligated into pSAB99 to create pSAB233. To generate the PfAnchor-BirA-HA strain, 100 μg of pSAB233 plasmid was linearized with Stu1 and transfected into 3D7-Cas9, along with 100 μg of pSAB175. 24 hours following transfection parasites were treated with 5 nM WR99210 for 7 days before maintaining on 2.5 nM WR99210 until resistant parasites were detected. Integration of pSAB233 plasmid into the genome of 3D7 was confirmed via PCR using primer pairs oSAB243/oJDD44, oSAB243/oSAB366, oSAB367/oJDD44, and oSAB367/oSAB366. Primer binding locations and predicted PCR product size found in Figure S13. All parasite lines used in this study are listed in Table S2.
Growth analysis:
For parasite growth analysis, parasite cultures were synchronized at the schizont stage by either density centrifugation using 60% Percoll PLUS or by magnetic separation using MACs columns, incubated at 37 °C for 2–3 hours with fresh erythrocytes, and then newly invaded ring stage parasites were purified by treatment with 5% w/v sorbitol. In three biological replicates, cultures were plated and incubated at 37 °C at 4% hematocrit in the presence or absence of aTC. Over the course of 96 hours (2 intraerythrocytic cycles) timepoints were taken at 0, 48, and 96 hours and the parasitemia was determined by counting a minimum of 400 RBCs.
Sodium carbonate extraction:
For the sodium carbonate extraction assay, the protocol from Liffner et al was used [76]. Saponin-lysed parasite pellets from 10 mL of high parasitemia late schizonts (40–48 hpi) were resuspended in 100 μL of Milli-Q water and snap-frozen using liquid nitrogen four times before passing through a 28 gauge needle. Samples were centrifuged for 10 minutes at 18,000 × g at 4 °C, with the supernatant reserved as the hypotonic (water soluble) sample. The pellet was washed twice with 500 μL Milli-Q water and once with 500 μL 1x PBS before the pellet was resuspended in 100 μL 0.1 M sodium carbonate (Na_2_CO_3_) and incubated in ice for 30 minutes. The sample was centrifuged at 18,000 × g at 4 °C for 5 minutes and the supernatant was reserved as the sodium carbonate (peripheral membrane) sample. The remaining pellet was washed 3 times in 100 μL 1x PBS before resuspending the pellet in 100 μL ice cold 0.1% Triton X-100/PBS and incubated for 30 minutes on ice. Samples were centrifuged at 18,000 × g at 4 °C and the supernatant was reserved before the pellet was washed and resuspended in 100 μL 1x PBS. Samples were analyzed by western blotting.
Proteinase K protection:
For Proteinase K protection assay, the protocol from Liffner et al was used [76]. 3× 10 mL cultures of schizonts (36–48 hpi) were lysed with 0.15% saponin/PBS-protease inhibitors (Sigma S8820–20 TAB, 1 tab for 100 mL PBS) for 10 minutes before centrifuging at 18,000 × g for 10 minutes at 4 °C. Pellets were washed 3x in 1x PBS-protease inhibitors before further treatment. Three treatments were performed: one tube treated with 250 μL SOTE buffer (0.6 M Sorbitol, 20 mM Tris HCl, 2 mM EDTA) alone. A second tube was treated with 250 μL 0.02% w/v digitonin in SOTE for 10 minutes on ice before centrifuging at 800 × g for 10 minutes at 4 °C. The resulting pellet was washed once with SOTE buffer. A third tube of sample was treated with digitonin followed by treatment with 0.1 mg/mL Proteinase K in SOTE for 30 minutes on ice. The sample was then centrifuged at 18,000 × g for 10 minutes at 4 °C before inactivating the Proteinase K by treatment of the pellet with 100 μL of 5 mM PMSF (ThermoFisher #36978) in SOTE buffer for 10 minutes on ice. Samples were analyzed by western blotting.
Western Blot:
Protein samples were collected by saponin lysis with 0.15% w/v saponin/PBS-protease inhibitors for 10 minutes on ice, parasite material was pelleted by centrifugation before washing 3x with PBS-protease inhibitors. Parasite pellets were resuspended in Laemmli sample buffer + 5% v/v β-mercaptoethanol (Aldrich), heated at 37 °C for 1 hour with vortexing every 30 minutes, and separated by size using BioRad 4–20% Mini-PROTEAN TGX stain free gels (BioRad Cat. #4568095) at 200 V for 40 minutes. Proteins were transferred to a nitrocellulose membrane using a BioRad Transblot Turbo Transfer System with the MixedMW setting (25 V, 1.3 A, 7 minutes) before blocking using 1:5 BioRad EveryBlot Blocking Buffer (BioRad Cat. #12010020): PBS for 30 minutes at room temperature. Membranes were incubated in primary antibodies (listed in Table S3) overnight at 4 °C, washed 3x in 1x TBS+ 0.01% Tween20 before incubating with secondary antibodies (listed in Table S3) at room temperature for 45 minutes. Western Blots were visualized using a BioRad ChemiDocMP Imaging system. Western blot quantification was done using BioRad Image Lab V6.1 software.
Live microscopy:
Glass bottomed microscopy dishes (Cellvis D35–20-1.5-N) were coated with 0.5 mg/mL concanavalin A (Sigma, C0412–5MG) solution for 1 hour at 37 °C before 3x washes with Milli-Q water. Synchronous late stage +aTC or -aTC parasites were purified by MACs columns and treated with compound 1 [46] before being applied to the glass bottomed dishes and incubated for 3 hours at 37 °C to allow parasites to adhere. After 3 hours samples were washed with 1 mL complete phenol-red-free RPMI. Brightfield and fluorescent images were taken every 0.5–4.5 seconds for 30 minutes per sample using a Leica DMI6000 B microscope and a 63x objective with a numerical aperture of 1.4.
Ultrastructure Expansion Microscopy (U-ExM):
12 mm round coverslips (Fisher, Cat# NC1129240) were treated with poly-D-lysine for 1 hour at 37 °C, washed twice with MilliQ water, and placed in the wells of a 24 well plate. Five hundred uL of parasite cultures adjusted to roughly 1% hematocrit was added to the wells containing a coverslip. Samples were allowed to settle for 30 minutes at 37 °C before culture supernatant was removed and 500 μL of 4% v/v PFA in 1x PBS was gently added along the side of the well and incubated at 37 °C for 20 minutes. Coverslips were washed once with 1x PBS before being treated with 500 μL of 1.4% v/v formaldehyde/2% v/v acrylamide (FA/AA) in PBS overnight at 37 °C. Gelation, denaturation, staining, and expansion of gels were performed as previously described [21]. Stained gels were imaged using a Zeiss LSM900 AxioObserver with an Airyscan 2 detector. Images were taken using a 63x Plan-Apochromat objective lens with a numerical aperture of 1.4. All antibodies and their dilutions used in this study are listed in Table S3.
Cryo-Electron Tomography
Sample preparation
PfAnchor iKD parasites were grown and synchronized as described above. Late schizont pellets were obtained via percoll purification and allowed to incubate in 5 mL of media with 2.5 uM Compound 1 [46] for 3 hours. Parasites were chemically fixed with 4% PFA/0.01% glutaraldehyde for 20 minutes before resuspending in PBS prior to cryo-electron tomography.
Cell vitrification:
Quantifoil Cu R2/1 grids were glow-discharged for 1 minute at 25 mA using an EmiTech K100X glow discharger (EMS). 4 μL of PfAnchor iKD Compound 1 arrested parasites grew in presence or absence of aTC (previously fixed with 4% PFA/glutamate PBS solution) cell culture were applied in 4 μL to each grid, manually back-blotted on one side (22°C and 100% humidity) for 3–4 seconds, and plunge-frozen in a 1:1 mixture of liquid ethane and propane using a Vitrobot Mk IV (ThermoFisher). Plunge-frozen grids were stored in liquid nitrogen.
Cryo-FIB Milling:
Cryo-FIB milling was performed using an Aquilos 2, a cryo-dedicated DualBeam microscope (ThermoFisher). Cryo-grids were sputter coated for 10 seconds, followed by a layer of organic platinum deposited by the gas injection system for 15 seconds, followed by another 10 seconds of sputter coating. Cell clusters were milled to generate lamella at ~150–200 nm thickness, using 0.1–0.3 nA ion beam current for rough milling and 30–50 pA for final polishing.
Cryo-ET:
Cryo-FIB milled lamella grids were transferred to a Titan Krios 3Gi microscope (ThermoFisher, FEI), equipped with a 300 kV field emission gun, a Selectris energy filter and Falcon 4i direct electron detector. Tilt series were collected using Tomo5 software (ThermoFisher) at 42,000x magnification (2.9 Å pixel size) and the energy filter at 20.0 eV slit width. Tilt ranges were adjusted for each acquisition site to the minimum and maximum tilt angles between −60° and +60° that encompassed the feature of interest. Tilt series were acquired in 3° increments with each tilt image recorded over 5 movie frames at defocus values between −4 to −8 μm. The total dose applied for each tilt series was ~100 e-/Å^2^.
Tomogram Reconstruction and Analysis:
Movie frames for each tilt series were motion corrected using MotionCor2 through Relion 3.1 using 5×5 patches. Motion-corrected tilt series were generated using EMAN2. Tilt series alignment, tomogram reconstruction, and CTF estimation were performed using the automated EMAN2 pipeline. Feature segmentation was performed using neural network training in EMAN2 or manually using IMOD 3dmod. Tomogram features were visualized and analyzed using ChimeraX.
Mevalonate bypass experiments
Azithromycin treatment:
To generate PfAnchor^Mev^ iKD parasites with disrupted apicoplasts, Clone F3 parasites were treated with 100 nM azithromycin (1× IC_50_) (Sigma PZ0007) for one week, with continuous supplementation of 50 μM mevalonate (Sigma M4667). The untreated parental Anchor knockdown line served as a positive control for apicoplast genome detection (Table S1).
Measurement of parasite growth:
To assess the dependence of apicoplast-intact and apicoplast-disrupted PfAnchor^Mev^ iKD parasites on mevalonate for survival, asynchronous parasites were washed three times with 10 mL of CMA to remove residual mevalonate and aTC. They were seeded in a 96-well plate at an initial parasitemia of 0.5% and a hematocrit of 2%, with a total volume of 200 μL per well. The parasites were then cultured under four conditions, with each condition tested in quadruplicate: (1) CMA with 50 μM mevalonate and aTC, (2) CMA with 50 μM mevalonate, (3) CMA with aTC, and (4) CMA alone. Cultures were incubated at 37°C in chambers gassed with 94% N_2_, 3% O_2_, and 3% CO_2_. Every other day, 10 μL of each sample was collected, diluted 1:10 in phosphate-buffered saline (PBS), and stored in a 96-well plate at 4°C. To prevent overgrowth, cultures were simultaneously diluted 1:8.
For growth curve determination, parasite samples were analyzed by flow cytometry over two intervals: days 0 to 4 and days 5 to 8. On day 4, 10 μL of each diluted sample was transferred to a new 96-well plate containing 100 μL of 1× SYBR Green (Invitrogen) in PBS per well. The plates were incubated at room temperature in the dark with gentle shaking for 30 minutes. After staining, 150 μL of PBS was added to each well to dilute excess dye. The samples were analyzed using an Attune Nxt flow cytometer (Thermo Fisher Scientific) with a 50-μL acquisition volume, a flow rate of 25 μL/min, and a total collection of 10,000 events per sample. The same protocol was repeated on day 8 for the second set of samples collected on days 6 and 8.
Analysis of apicoplast genome
To assess the integrity of the apicoplast genome after treatment with azithromycin, PfAnchor^Mev^ iKD parasites were grown for 5 days in the presence of 100 nM azithromycin and 50 μM mevalonate before aTC washout. Genomic DNA was extracted using QIAmp DNA blood mini kit (Qiagen, #51106) every two days up to day eight. Control samples from PfAnchor^Mev^ iKD not treated with azithromycin were also collected. PCR analysis was performed using primers oSAB484/oSAB485 (GAPDH, nuclear DNA), oSAB486/oSAB487 (tufa, apicoplast DNA), and oSAB488/oSAB489 (cytb3, mitochondrion DNA) as previously described [77].
Immunoprecipitation and proximal labeling
Sample preparation
Protein extraction for both immunoprecipitation (IP) and BioID experiments was performed using the same lysis procedure. Parasites were cultured at 2% hematocrit and 4–5% parasitemia before harvesting. Red blood cells were lysed using 0.05% saponin in PBS supplemented with protease inhibitors (SigmaFast, Sigma S8820–20TAB) and incubated on ice for 10 minutes. Parasites were washed three times with ice-cold PBS + protease inhibitors by centrifugation at 10,000 g for 5 minutes at 4°C. The resulting pellets were resuspended in 1 mL PBS + protease inhibitors, centrifuged at 10,000 rpm for 2 minutes at 4 °C, and stored at −80 °C or processed immediately. Parasite pellets were lysed in 1 mL RIPA buffer (50 mM Tris-HCl pH 7.5, 150 mM NaCl, 1% NP-40, 0.5% sodium deoxycholate, 0.1% SDS) supplemented with protease inhibitors. Samples were incubated on a rotator at 4 °C for 30 minutes. To ensure efficient lysis, three rounds of sonication (30 seconds at 20 amplitude) were performed on ice, with 3-minute rest intervals between pulses. Lysates were cleared by centrifugation at 15,000 g for 30 minutes at 4 °C, and the supernatant was transferred to a fresh tube. For the MCMBP-BioID experiments, soluble and insoluble fractions were processed separately using the same protocol described in our previous study [39]. For immunoprecipitation, 50 μL of anti-V5 magnetic beads (Thermo, #88816) per sample were washed twice with 1 mL RIPA buffer and incubated with the cleared lysates overnight at 4 °C with constant rotation. Beads were collected using a magnetic stand, washed three times with 50 mM ammonium bicarbonate buffer, and resuspended for on-bead digestion and mass spectrometry analysis. For BioID, 50 μL of streptavidin magnetic beads (Thermo, #88816) were used instead of V5 magnetic beads. After overnight incubation, beads were washed following the same procedure as in IP before processing for mass spectrometry analysis.
On bead digests:
After washing, beads were covered with 8 M Urea, 100 mM Tris hydrochloride, pH 8.5, reduced with 5 mM tris (2-carboxyethyl) phosphine hydrochloride (TCEP, Sigma-Aldrich Cat No: C4706) for 30 minutes at room temperature to reduce the disulfide bonds. The resulting free cysteine thiols were alkylated using 10 mM choloracetamide (CAA, Sigma Aldrich Cat No: C0267) for 30 minutes at RT, protected from light. Samples were diluted to 2 M Urea with 50 mM Tris pH 8.5 and proteolytic digestion was carried out with Trypsin/LysC Gold (0.4 μg, Mass Spectrometry grade, Promega Corporation Cat No: V5072) overnight at 35 °C. After digestion, samples were quenched with 0.4% trifluoroacetic acid (v/v, Fluka Cat No: 91699). Samples were either injected onto a trap column followed by analytical column or first subjected to a solid phase extraction clean up on Pierce C18 spin columns (Cat No 87870).
Liquid Chromatography Mass Spectrometry:
Approximately 1/15^th^ of each IP sample was loaded onto a 5 cm C18 trap column Acclaim^™^ PepMap^™^ 100 (3 μm particle size, 75 μm diameter; Thermo Scientific, Cat No: 164946) followed by a 25 cm EASY-Spray column (Thermo Scientific, Cat No: ES902) and analyzed using a Q-Exactive Plus mass spectrometer (Thermo Fisher Scientific) operated in positive ion mode. Solvent B was increased from 5%−35% over 100 min, to 90% over 2 min, back to 3% over 2 minutes (Solvent A: 95% water, 5% acetonitrile, 0.1% formic acid; Solvent B: 100% acetonitrile, 0.1% formic acid). A data dependent top 15 method was used with MS scan range of 350–2000 m/z, resolution of 70,000, automatic gain control (AGC) target 3e6, maximum injection time (IT) of 200 ms. MS2 resolution of 17,500, scan range of 200–2000 m/z, normalized collision energy of 30, isolation window of 4 m/z, target AGC of 1e5, and maximum IT of 150 ms. Dynamic exclusion of 10 sec, charge exclusion of 1, 7, 8, >8 and isotopic exclusion parameters were used. For samples post C18 cleanup, after drying in a speed vacuum, samples were resuspended in 25 μL 0.1 % FA. Approximately 1/5^th^ of each sample was then injected using an EasyNano1200 LC coupled to an Exploris 480 orbitrap mass spectrometer (Thermo Fisher Scientific)). Solvent B (80% Acetonitrile, 0.1 % FA) was increased from 8–35 % over 90 minutes, increased from 35–65 % over 15 min, increased to 85% over 5 min, held at 85% for 5 min, and decreased to 4% over 5 min. The mass spectrometer was operated in positive ion mode, advanced peak determination on, default charge state of 2 and user defined lock mass of 445.12003. 4 second cycle time was used with MS1 parameters of scan range 375–1500 m/z: orbitrap resolution of 60,000, standard AGC, automatic max IT, and RF lens of 40%. Monoisotopic peak determination was set to peptide with a minimum intensity filter of 5e3, charge state filter of 2–7, and dynamic exclusion of 30 s with 5 ppm mass tolerance. MS2 parameters included an isolation window of 4 m/z, normalized high energy dissociation energy of 30 %, orbitrap resolution of 15,000, user defined first mass of 110 m/z, standard AGC target and auto max IT. Data were recorded using Tune application 4.2.362.42.
Data analysis:
Data were analyzed using Proteome Discoverer 2.5.0.400 (Thermo Fisher Scientific). Plasmodium falciparum 3D7 and Homo sapiens reference proteome databases (downloaded from Uniprot on 04/06/2021 with 5381 sequences and on 05/13/2022 with 78806 sequences respectively), plus common laboratory contaminants (73 sequences) were searched using SEQUEST HT. Precursor mass tolerance was set to 10 ppm and fragment mass tolerance set at 0.02 Da with a maximum of 3 missed cleavages. A maximum of 3 modifications were allowed per peptide. Dynamic modifications included methionine oxidation; phosphorylation on serine, threonine, and tyrosine; dynamic protein terminus modifications were acetylation, met-loss, and met-loss plus acetylation. Static modifications were carbamidomethylation on cysteines. Percolator false discovery rate (FDR) filtration of 1% was applied to both the peptide-spectrum match and protein levels. Search results were loaded into Scaffold Q + S Software (version 5.2.2, Proteome Software, Inc) for visualization.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Burki F., , The New Tree of Eukaryotes. Trends in Ecology & Evolution, 2020. 35(1): p. 43–55.31606140 10.1016/j.tree.2019.08.008 · doi ↗ · pubmed ↗
- 2Collier S.L., , Modes and mechanisms for the inheritance of mitochondria and plastids in pathogenic protists. PLOS Pathogens, 2025. 21(1): p. e 1012835.39847585 10.1371/journal.ppat.1012835 PMC 11756805 · doi ↗ · pubmed ↗
- 3Lewis W.H., , Plastid translocon recycling in dinoflagellates demonstrates the portability of complex plastids between hosts. Current Biology, 2024. 34(23): p. 5494–5506.e 3.39571577 10.1016/j.cub.2024.10.034PMC 7617431 · doi ↗ · pubmed ↗
- 4Gray M.W., The endosymbiont hypothesis revisited. Int Rev Cytol, 1992. 141: p. 340, 221–242.10.1016/s 0074-7696(08)62068-91452433 · doi ↗ · pubmed ↗
- 5Gould S.B., Waller R.F., and Mc Fadden G.I., Plastid evolution. Annu Rev Plant Biol, 2008. 59: p. 491–517.18315522 10.1146/annurev.arplant.59.032607.092915 · doi ↗ · pubmed ↗
- 6Goodman C.D., Buchanan H.D., and Mc Fadden G.I., Is the Mitochondrion a Good Malaria Drug Target? Trends in Parasitology, 2017. 33(3): p. 185–193.27789127 10.1016/j.pt.2016.10.002 · doi ↗ · pubmed ↗
- 7Low L.M., Stanisic D.I., and Good M.F., Exploiting the apicoplast: apicoplast-targeting drugs and malaria vaccine development. Microbes and Infection, 2018. 20(9): p. 477–483.29287981 10.1016/j.micinf.2017.12.005 · doi ↗ · pubmed ↗
- 8Organization, W.H., World Malaria Report 2023. 2023. 1–356.