Dissecting Metabolic Control of Behaviors and Physiology During Aging in Drosophila
Elizabeth S. Pasam, Kishore Madamanchi, Girish C. Melkani

TL;DR
This study uses fruit flies to explore how metabolism affects aging-related changes in behavior and physiology, focusing on the role of specific genes and their effects on sleep, movement, and lipid balance.
Contribution
The study identifies cell-autonomous and non-cell-autonomous roles of metabolic genes in age-related decline and establishes Ampkα as a central regulator of behavioral and metabolic aging.
Findings
Knockdown of SdhD, Marf, and Gnmt reduces flight performance in aged flies, showing both cell-autonomous and non-cell-autonomous effects.
AMPK signaling modulates sleep and activity rhythms in an age- and tissue-specific manner, with overexpression altering lipid homeostasis in the brain.
Genes like mAcon1, LSD2, and Ald show cell-autonomous effects on flight performance and sleep patterns during aging.
Abstract
Aging disrupts physiological and behavioral homeostasis, largely driven by one-carbon metabolism, mitochondrial dysfunction, energy sensing, and metabolic imbalance. To elucidate the roles of conserved metabolic, energy sensing, and mitochondrial genes in age-related decline, we employed genetic manipulations in vivo using Drosophila melanogaster models, in a cell-autonomous and non-cell-autonomous manner. By using panneuronal and indirect flight muscle (IFM)- specific drivers, we assessed the impact of gene knockdown or overexpression on sleep-circadian rhythms, locomotion, and lipid metabolism in a cell-autonomous and non-cell-autonomous manner to address bidirectional neuro-muscle communications. Knockdown of genes such as SdhD, Marf, and Gnmt leads to decrease in flight performance especially in 6 weeks with both the drivers. Which demonstrates cell-autonomous and non-cell…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
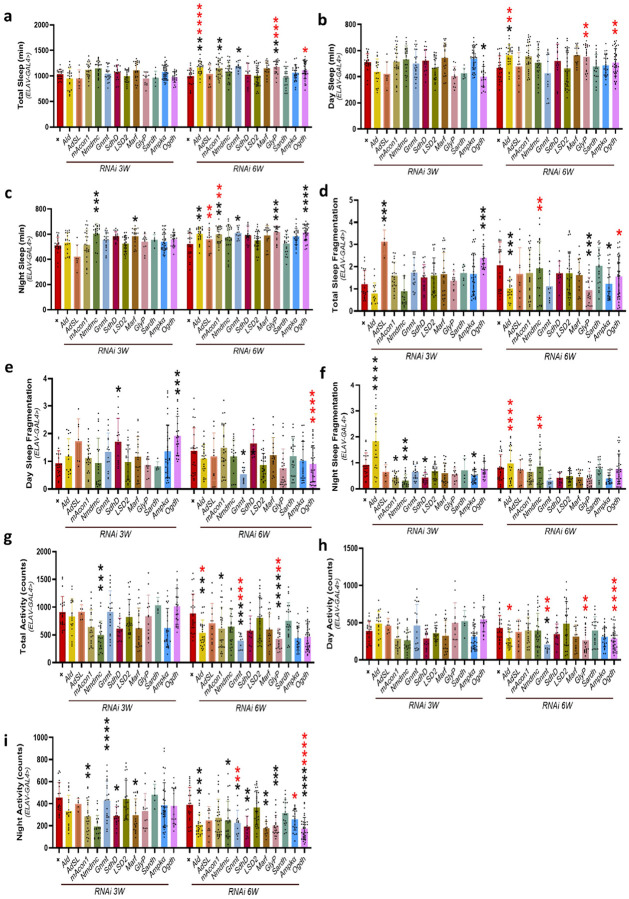 Figure 1
Figure 1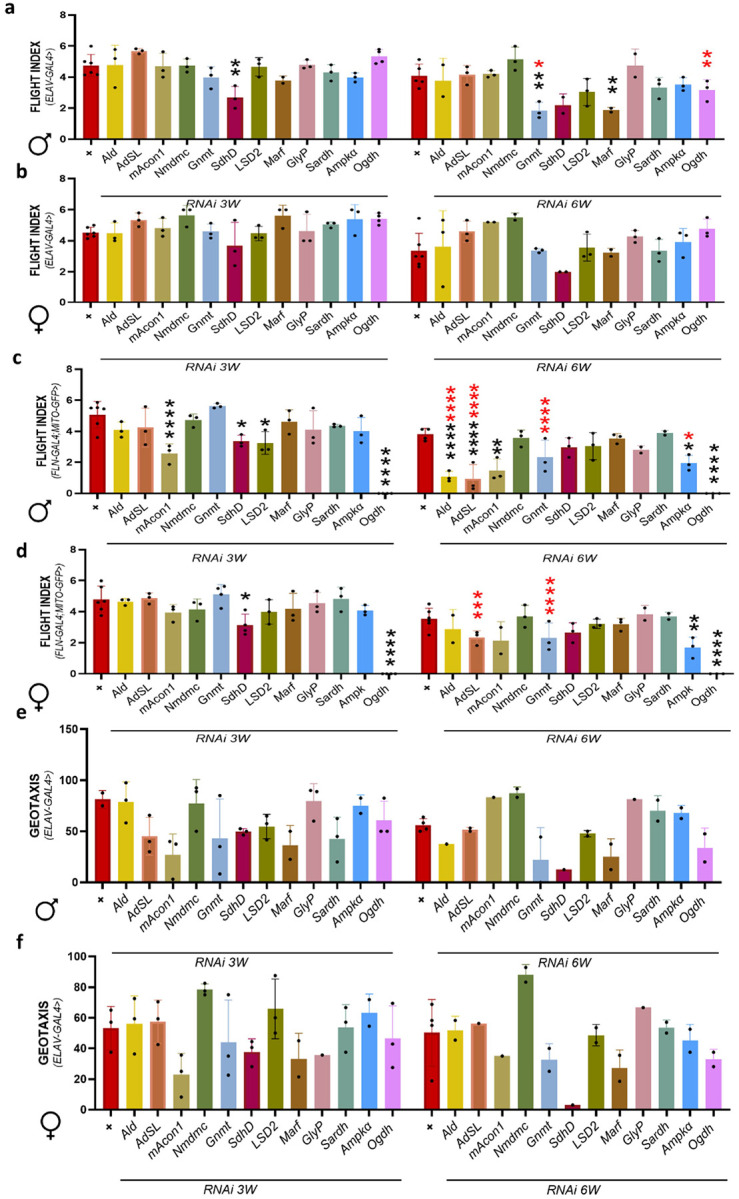 Figure 2
Figure 2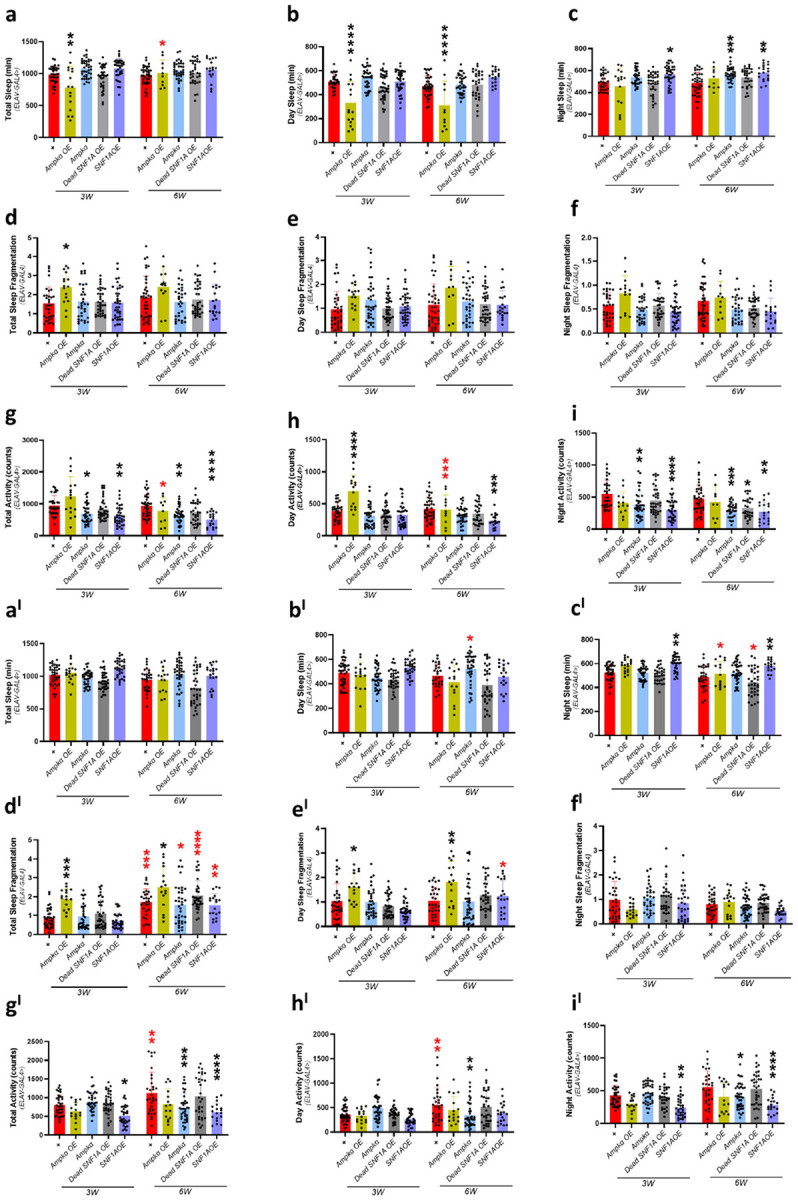 Figure 3
Figure 3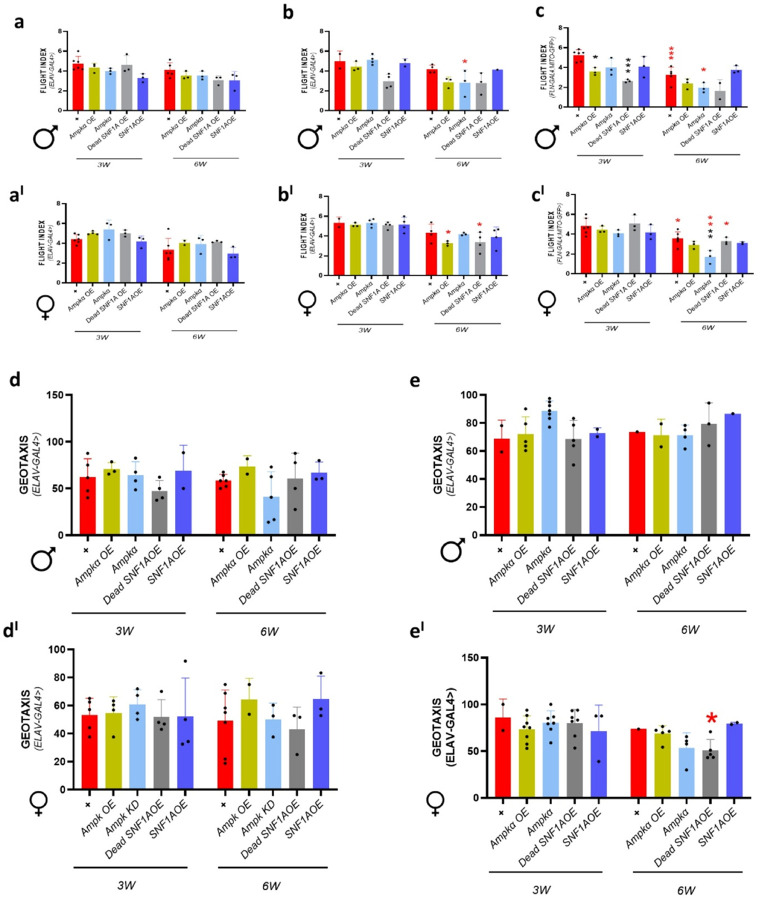 Figure 4
Figure 4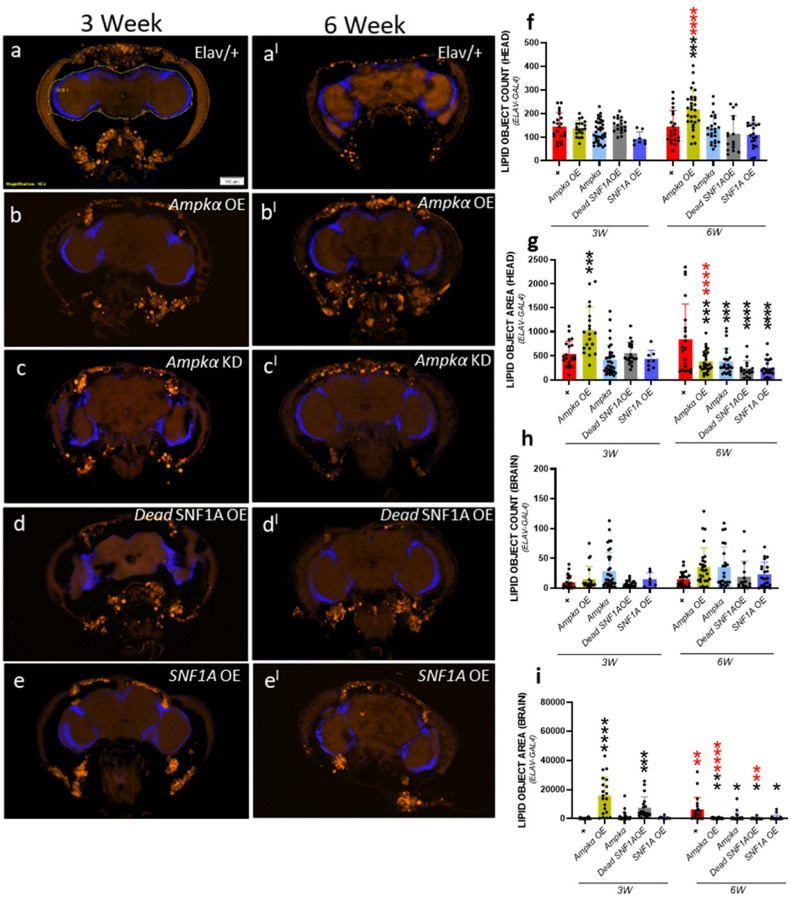 Figure 5
Figure 5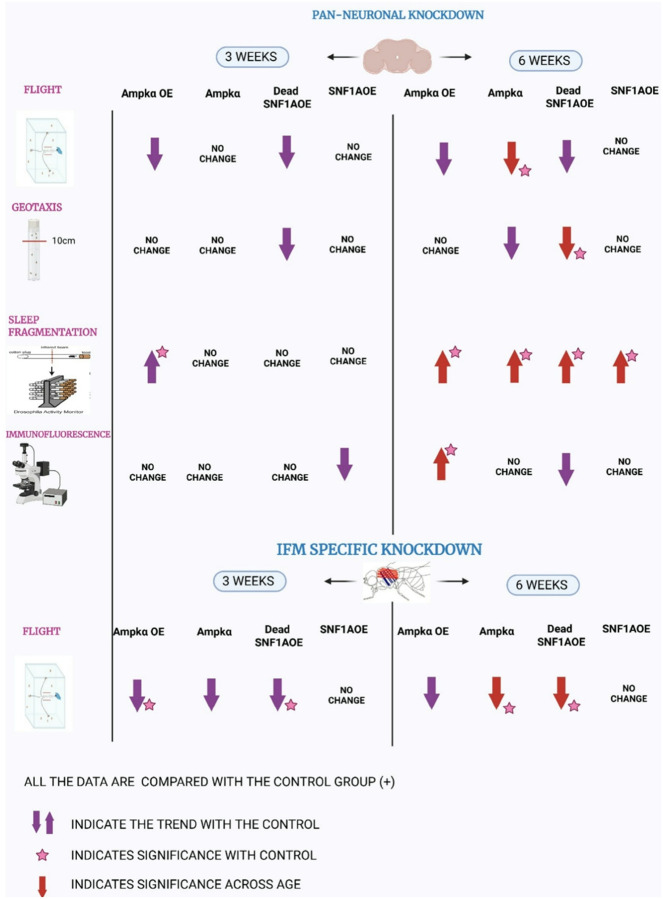 Figure 6
Figure 6Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsGenetics, Aging, and Longevity in Model Organisms · Neurobiology and Insect Physiology Research · Adipose Tissue and Metabolism
Introduction
Aging is a complex phenomenon that causes a gradual decline in the biological function of multiple organ systems in a time-dependent manner (Gellert and Alonso-Perez, 2024; Sergeev et al., 2025). It is typically related to lower stress resilience, altered metabolic balance, and impaired physiological, mental, and behavioral capacities (Majnaric et al., 2021). Age-related alterations in sleep architecture, motor coordination, cognitive performance, and muscle integrity are indicators of systemic deterioration across species, including humans (Lopez-Otin et al., 2013; Vaillancourt and Newell, 2002). These external symptoms are closely linked to underlying cellular changes, particularly mitochondrial malfunction and metabolic imbalance (Amorim et al., 2022; Bartman et al., 2024). At the cellular level, mitochondria control energy production, redox equilibrium, calcium buffering, and apoptotic signaling (Matuz-Mares et al., 2022). However, with age, mitochondria lose their efficiency, resulting in increased reactive oxygen species (ROS) production, decreased ATP synthesis, and dysregulated metabolic pathways (Chaudhari and Kipreos, 2018; Giorgi et al., 2018). These changes not only affect cellular function but also lead to overall tissue degeneration, especially in high-energy-demanding neurological and muscular systems (Clemente-Suarez et al., 2023; Hargreaves and Spriet, 2020). Alterations in metabolism, particularly those involving lipid, amino acid, and glucose pathways, have been linked to age-related diseases (Lien et al., 2023; Mirzaei et al., 2014; Semba et al., 2018), but the precise roles of various metabolic and mitochondrial genes in tissue-specific aging outcomes are unclear.
Despite increased recognition of mitochondria and metabolism’s critical role in aging (Amorim et al., 2022), considerable knowledge gaps exist in understanding how these pathways differentially govern age-related decline across tissues and behaviors. The functional role of evolutionarily conserved metabolic and mitochondrial genes in controlling behavioral aging characteristics such as sleep-circadian cycles, locomotion, and physical performance is particularly poorly understood. To reduce these gaps, we employed Drosophila as a model system to conduct a functional genetic search for conserved metabolic and mitochondrial genes (Brischigliaro et al., 2023). Drosophila provides distinct benefits for aging research, including a well-characterized genome, a short life cycle, and well-conserved energy regulatory mechanisms (Chatterjee and Perrimon, 2021). Furthermore, the availability of tissue-specific Gal4 drivers allows precise regulation of gene expression in neurons and muscles (Tain et al., 2021), both of which are crucial for maintaining behavioral and physiological homeostasis during aging.
We selected genes based on their established roles in key metabolic processes, like glucose metabolism, glycogen metabolism, tricarboxylic acid (TCA) cycle, fat, lipid metabolism, and mitochondrial functions. Such as glycogen phosphorylase (GlyP), adenylosuccinate lyase (ADSL), glycine N-Methyltransferase (GNMT), sarcosine dehydrogenase (SARDH), succinate dehydrogenase subunit complex D (SDHD), oxoglutarate dehydrogenase (OGDH), AMP-activated protein kinase α (Ampkα), NAD-dependent methylenetetrahydrofolate dehydrogenase (NMDMC), mitochondrial aconitase 1 (mAcon1), Aldolase (ALD), lipid storage droplet 2 (LSD-2), and mitochondrial assembly regulatory factor (MARF). These genes were selected with the knowledge of prior evidence in mitochondrial stress response, aging-related metabolic regulation, and evolutionary conservation (Livelo et al., 2022). Our hypothesis is embedded in the observation that age-dependent decline in behavior and physiology may be driven by tissue-specific mitochondrial and metabolic dysfunction. To test this, we used panneuronal driver (Elav-Gal4) and indirect flight muscle (IFM) specific driver, recombinant stock with Mito-GFP (Fln-Gal4;Mito-GFP) to knock down and overexpress these genes and evaluated their effects on sleep architecture, activity patterns, geotaxis, flight performance, and lipid accumulation at mid (3-week-old) and late (6-week-old) adult ages. It is known that Ampkα plays a key role in behavioral aging (Burkewitz et al., 2014; Salminen and Kaarniranta, 2012), as our investigation could reveal tissue-specific changes in energy homeostasis, behavioral functions. This study contributes to our understanding of how mitochondrial and metabolic genes influence the aging trajectory in a tissue-specific way by combining targeted gene alteration with behavioral phenotyping. Additionally, it offers a useful framework for locating potential targets that could be the basis of conserved mechanisms underlying age-related declines in cognitive and physical performance.
We hypothesize that aging-associated physiological and behavioral decline is driven in part by tissue-specific dysregulation of conserved metabolic and mitochondrial genes. Specifically, neuronal and muscular impairment in sleep-circadian rhythms, locomotion, and energy balance may result from disruptions in genes involved in energy sensing, TCA cycle flux, amino acid metabolism, lipid regulation, and mitochondrial dynamics. By functionally modulating these genes in a tissue- and age-dependent manner, we aim to uncover how their differential roles contribute to systemic aging phenotypes. We further hypothesize that Ampkα represents a key node that integrates metabolic stress and behavioral aging, and its dysregulation may drive both autonomous and non-cell-autonomous effects on neural and muscular function.
Materials and Methods
Fly stocks and expression system:
All Drosophila stocks were maintained on a standard cornmeal-yeast-agar diet composed of 11 g/L agar, 30 g/L active dry yeast, 55 g/L yellow cornmeal, and 72 mL/L molasses, supplemented with 8 mL/L of 10% nipagin and 6 mL/L propionic acid to prevent microbial growth. Flies were reared under controlled environmental conditions at 22°C with 50% relative humidity and a 12:12 hour light-dark cycle. Fresh food vials were provided every three days (Moraes et al., 2024). To evaluate the functional impact of designated metabolic and mitochondrial genes on muscle physiology and sleep behavior, we used tissue-specific overexpression and RNA interference (RNAi) lines. UAS-RNAi lines were identified from the Vienna Drosophila Resource Center (VDRC) and the Bloomington Drosophila Stock Center (BDSC). The RNAi lines included: Gnmt (BDSC #42637), Sardh (BDSC #51883), Marf (BDSC #31157), Nmdmc (BDSC #62268), Adsl (BDSC #34347), Ampkα (BDSC #57785), GlyP (VDRC #27928), Ald1 (BDSC #26301), Ogdh (BDSC #33686; VDRC #50393), mAcon1 (BDSC #34028), SdhD (BDSC #65040), Sicily (BDSC #55442), LSD-2 (VDRC #40734), and corresponding control RNAi lines (BDSC #36303, #36304). Most of the lines are previously used in our previous studies; hence, we are using a single line for each gene (Livelo et al., 2023). We employed several overexpression lines obtained from BDSC: UAS-GFP (BDSC #5431), UAS-Ampkα (BDSC #32108), UAS-Dead SNF1A (BDSC #32112), UAS-SNF1A (BDSC #32110). Tissue-specific gene manipulation was performed using the UAS-Gal4 system (Barwell et al., 2017). Panneuronal expression of RNAi or overexpression constructs was achieved using Elav(X)-Gal4 from Bloomington Drosophila Stock Center (BDSC) (BL#458) and Elav-Gal4 (BL#8765). To explore the cell-autonomous contribution in the muscle tissue, we employed the Flitin-Gal4 driver, recombined with UAS-Mito-GFP (referred to as Fln-Gal4:Mito-GFP, hereafter) driver, which targets gene expression specifically to IFM (BDSC #84977). Each knockdown and overexpressed line was crossed with panneuronal or IFM-specific drivers, F-1 progeny were collected. Males and female progeny were separated and transferred to fresh food vials every 3–4 days throughout our study. All these experiments were performed with the indicated number of flies and replicates as shown in the source data file using 3-week (mid age) and 6-week (old age) male and female flies.
Locomotor Performance
Flight assay:
To evaluate the functional role of target genes on neuromuscular performance, flight ability was evaluated in adult Drosophila using knockdown or overexpression lines under the control of the panneuronal Elav-Gal44 or the indirect flight muscle-specific Fln-Gal4; Mito-GFP drivers. The flight assay was adapted from standard methodologies to quantify age- and genotype-dependent motor performance (Drummond et al., 1991; Livelo et al., 2025). Briefly, groups of 10–20 adult Drosophila were gently released into the center of a vertically oriented Plexiglass flight chamber illuminated from above. Based on their directional flight responses upward (score = 6.0), horizontal (4.0), downward (2.0), or flightless (0.0), individual Drosophila were scored, and a Flight Index (FI) was calculated for each cohort. This index reflects the average flight capacity of a group and was used to compare across genotypes and age groups. All experiments were conducted in parallel with age-matched control lines. Detailed information regarding fly age, genotype, experimental conditions, number of cohorts, total Drosophila tested, and cohort-wise flight index values is provided in the Source Data file.
Geotaxis assay:
Flies were moved to a fresh vial (with 10–20 Drosophila per trial using at least 3 biological replicates per condition) and given a 2-minute period to acclimate. Subsequently, the vial underwent three taps to induce a negative geotaxis reaction. The Drosophila climbing behavior was recorded on video for later analysis. At 10-s intervals, the proportion of Drosophila that successfully reached the 10cm mark was recorded (Livelo et al., 2025; Villanueva et al., 2019).
Sleep activity analysis:
Sleep-wake behavior and circadian activity were assessed using the Drosophila Activity Monitoring (DAM) system (TriKinetics Inc., MA, USA) under controlled 12-hour light:12-hour dark (12L:12D) conditions at 25°C. Experiments were conducted on male progeny of Elav(x)-Gal4 and ElavII-Gal4 drivers for neuronal knockdown alongside respective genetic control lines. Activity was recorded as infrared beam crossings in individual glass tubes, representing locomotor bouts. A sleep bout was defined as a minimum of 5 consecutive minutes of inactivity (i.e., zero beam crossings). Sleep behavior was quantified using ClockLab (Actimetrics) and RStudio (Yadav et al., 2025), with custom R scripts available at https://github.com/jameswalkerlab/Gill_et.al.
Immunofluorescence analysis:
As recently reported in detained in the method paper (Watson et al., 2025), under a microscope, the experimental fly heads were dissected and fixed for 15 minutes in 4% paraformAldehyde (PFA) in phosphate-buffered saline (PBS) to quantify lipid accumulation. The heads were then rinsed three times (10 minutes each) in 1× PBS with agitation. To ensure cryoprotection, these samples were treated overnight in 10% sucrose in PBS. Heads were implanted in OCT compound (Fisher Scientific #4585) and cryosectioned at 20 μm thickness on a Leica CM3050 S cryostat. Sections were mounted on pre-warmed microscope slides (Fisher #15-188-48), air-dried for 30 minutes, and then protected with a hydrophobic barrier. After washing, the slides were incubated for an hour with Lipid Spot 488 (Thermo Fisher Scientific #70065). Slides were mounted using VECTASHIELD Vibrance Antifade Mounting Medium with DAPI (0.9 μg/mL, H-1800). Images were acquired at 10× magnification using an Olympus BX63 fluorescence microscope with CellSens software.
Statistical analysis:
Statistical analyses were carried out using GraphPad Prism version 10. For behavioral tests such as sleep parameters and climbing abilities (geotaxis), two-way ANOVA was used to establish significance, followed by Sidak’s multiple comparisons test to assess the effects of genotype and age. We have also compared each of the knock-down genes with the control RNAi and each of the overexpressed genes with the GFP overexpression line. These data will be available. Data are reported as mean ± SD. Statistical significance was defined as the following: p < 0.05 (), p < 0.01 (), p < 0.001 (), p < 0.0001 (****). Detailed statistical analyses among different genotypes during aging have been shown in the source data.
Results
Panneuronal knockdown of metabolic and mitochondrial genes altered sleep duration and enhanced sleep fragmentation:
To understand the impact of these genes on physiological and behavioral functions, we have tested knockdown of various genes that are involved in glycogen metabolism, TCA cycle, fatty acid metabolism, mitochondrial function, and energy production. We have used Adsl, Gnmt, GlyP, Sardh, Ampkα, ALD, LSD-2, mAcon1, Nmdmc, SdhD, Marf gene RNAi fly lines to understand their impact on physiological and behavioral functions individually, during aging under panneuronal (Elav-Gal4) driver (Figure 1). At mid (3-week-old) age, we observed increased total sleep in mAcon1, Gnmt, compared to controls, and Ald, GlyP, Ogdh showed increased total sleep compared to 3-week Drosophila and with control (Figure 1a). Decreased day sleep at 3-week-old in Ogdh, increased day sleep in Ald at 6-week-old compared to their control. Whereas Ald, GlyP, and Ogdh increased day sleep at 6-week-old compared to 3-week-old Drosophila (Figure 1b). Night sleep increased in Nmdmc, Marf at 3-week-old, Ald, mAcon1, Gnmt, GlyP, Ogdh at 6-week-old compared to the control. Whereas Adsl, mAcon1 showed increased night sleep compared to 3-week-old Drosophila (Figure 1c). Total sleep fragmentation (Figure 1d) increased in Adsl, and Ogdh at 3-week-old and decreased in Ald, GlyP, and Ampkα at 6-week-old compared to controls. Whereas Nmdmc showed increased and Ogdh showed decreased total sleep fragmentation compared to 3-week-old Drosophila. Day sleep fragmentation increased in SdhD and Ogdh (Figure 1e) at 3-week-old compared to control and decreased in Ogdh at 6-week-old compared to 3-week-old Drosophila. Night sleep fragmentation increased in Ald and decreased in mAcon1, Nmdmc, SdhD, Ampkα compared to control at 3-week-old. At 6 weeks old, Ald showed a decrease and an increase in Nmdmc statistically significantly compared to 3-week-old Drosophila (Figure f). Total activity statistically significantly decreased in Nmdmc compared to control in 3-week-old and decreased in Ald, mAcon1, Gnmt, GlyP compared to 6-week-old control, also Ald, Gnmt, GlyP compared to 3-week-old Drosophila (Figure 1g). At old (6-week-old) age, Day activity (Figure 1h) decreased in Gnmt compared to control and Ald, Gnmt, GlyP, and Ogdh compared to 3-week-old Drosophila. Panneuronal knockdown of metabolic and mitochondrial genes caused apparent and vigorous changes in sleep architecture and activity during aging. Genes such as Ald, mAcon1, Gnmt, and GlyP showed increased sleep and reduced activity, indicating a compensatory energy-conservation response. In contrast, Ogdh, Adsl, and Nmdmc were associated with increased sleep fragmentation and reduced daytime activity, suggesting interrupted neuronal energy regulation. These findings highlight the critical role of neuronal metabolism in maintaining sleep-wake stability with age.
Locomotor ability was compromised upon IFM-specific knockdown of metabolic and mitochondrial genes compared to panneuronal knockdown:
Panneuronal expression of SdhD at 3-week-old and 6-week-old Gnmt, Marf knockdown Drosophila showed decreased flight performance in males compared to control Drosophila. Also, at 6 weeks old, Gnmt and Ogdh knockdown Drosophila have reduced flight index compared to 3-week-old male Drosophila (Figure 2a). Whereas female Drosophila have no statistically significant difference at 3-week-old as well as 6-week-old age, unlike males (Figure 2b). IFM specific expression of mAcon1, SdhD, LSD-2, and Ogdh has shown statistically significant reduction at 3 weeks and Ald, Adsl, mAcon1, Ampkα, Ogdh at 6 weeks of age compared to controls. In addition, at 6-week-old Ald, Adsl, Gnmt, and Ampkα knockdown Drosophila showed statistically significantly low flight performance compared to 3-week-old knockdown male Drosophila (Figure 2c). In female Drosophila SdhD, and Ogdh Drosophila at 3-week-old and Ampkα, Ogdh knockdown at 6-week-old showed statistically significant reduction compared to controls. Whereas Adsl, Gnmt knockdown Drosophila showed reduced flight performance compared to 3-week-old female knockdown Drosophila (Figure 2d). We then analyzed the geotaxis performance using panneuronal driver to understand the non-cell-autonomous Elav males (Figure 2e) and Elav females (Figure 2f) relationship. In our study, we have noticed a close trend in some of the knockdown genes, but the significance of the difference is hindered due to the limited number of replicates. Our study showed that IFM and panneuronal-specific knockdown of metabolic and mitochondrial genes such as SdhD, Marf, and Gnmt (Fernandez-Tussy et al., 2019) lead to the statistically significant decline in locomotor ability, showed neuromuscular interaction specifically in aging male Drosophila. Key genes like Ampkα, Ogdh, Adsl, and mAcon1 were extensively impaired in-flight performance in a sex- and age-dependent manner. Overall, female Drosophila were least affected but still showed genotype-specific vulnerabilities.
Panneuronal modulation of Ampkα and SNF1A reveals kinase-dependent roles in regulating sleep and activity rhythms.
In this study, we employed panneuronal modulation of Ampkα signaling using Elav-Gal4 to investigate its role in neuronal metabolism and function. We used two independent Elav-Gal4 drivers inserted on different chromosomes, Elav-Gal4 (X) and Elav-Gal4 (II). Both drive expressions in post-mitotic neurons, but differ in chromosomal location, which helps control for position effect variegation and background genotype effects. We overexpressed Ampkα to enhance energy-sensing activity and promote neuroprotection, while Ampkα knockdown allowed us to assess its necessity in maintaining neuronal homeostasis. To explore the evolutionary conservation of function, we overexpressed the yeast homolog SNF1A, and to further dissect kinase-dependent versus independent roles, we expressed a kinase-dead SNF1A mutant (DeadSNF1A). This approach enables us to evaluate both the functional significance and mechanistic specificity of Ampkα and SNF1A signaling in the nervous system. With the Elav(II) driver, we have observed a statistically significant reduction in total sleep (Figure 3a) at 3-week-olds in Ampkα overexpression compared to control, and at 6-week-olds, total sleep was increased compared to 3-week-old Ampkα overexpression Drosophila. Day sleep in statistically significantly reduced in Ampkα overexpression at 3– 6-week-old age (Figure 3b) compared to the respective control Drosophila. Night sleep was statistically significantly increased in SNF1A overexpression at 3-week-olds and 6-week-olds in Ampkα knockdown, SNF1A overexpression Drosophila compared to wild-type control (w^1118^) Drosophila (Figure 3c). Total sleep fragmentation statistically significantly increased in 3-week Ampkα overexpression Drosophila compared to wild-type control (w^1118^) Drosophila (Figure 3d). Day sleep fragmentation (Figure 3e) and night sleep fragmentation (Figure 3f) did not show any statistically significant difference compared to controls. Total activity was reduced in Ampkα knockdown, and SNF1A overexpression at 3-week-old and at 6-week-old age compared to controls, and Ampkα overexpression showed statistically significant reduction compared to 3-week-old Drosophila (Figure 3g). Day activity was statistically significantly increased in Ampkα overexpression compared to wild-type control (w^1118^) at 3-week-olds and at 6-week-olds, Ampkα overexpression Drosophila showed reduced day activity compared to 3-week-old Ampkα overexpression Drosophila (Figure 3h). The night activity was statistically significantly reduced in Ampkα knockdown and SNF1A overexpression Drosophila at 3-week-olds, compared to control and at 6-week-old Ampkα knockdown, Dead SNF1A overexpression and SNF1A overexpression Drosophila showed reduced night activity compared to control Drosophila (Figure 3i). With the Elav(X) driver at 3-week-olds, Ampkα overexpression showed reduced total sleep and at 3-week-olds compared to controls, and at 6-week-olds increased total sleep compared to 3-week-old Ampkα overexpression Drosophila (Figure 3aI). Day sleep (Figure 3bI) decreased only in Ampkα overexpression Drosophila at 3,6-week-old compared to age-matched wild-type control (w^1118^). Night sleep increased in Ampkα overexpression at 3-week-old compared to controls, with no observed change in other genotypes (Figure 3cI). Total sleep fragmentation (Figure 3dI) increased only in Ampkα overexpression Drosophila at 3,6-week-old compared to age-matched wild-type control (w^1118^). We found a statistically significant rise in day sleep fragmentation at 6-week-olds and night sleep fragmentation at 3-week-olds in Ampkα overexpression Drosophila (Figure 3eI, fI) compared to age-matched wild-type control (w^1118^) Drosophila. Total activity was statistically significantly decreased in Ampkα knockdown and SNF1A overexpression at 3-week-old age compared to age-matched Drosophila wild-type control (w^1118^), at 6-week-olds, Ampkα overexpression showed reduced total activity than 3-week-old Drosophila and SNF1A overexpression Drosophila compared to control (Figure 3gI). Day activity showed a statistically significant increase in Ampkα overexpression, compared to control Drosophila at 3-week-olds, but 6-week-old Drosophila showed a statistically significant reduction than 3-week-old Drosophila (Figure 3hI). Whereas night activity statistically significantly decreased in SNF1A overexpression at 3-week-old and Ampkα knockdown at 6-week-old compared to age-matched wild-type control (w^1118^) Drosophila (Figure 3iI). Our results indicate that panneuronal modulation of Ampkα and SNF1A distinctly affects sleep architecture and activity rhythms in an age-dependent manner in two different drivers. Ampkα overexpression initially reduces total sleep and increases fragmentation, but reverses at older ages, proposing adaptive or compensatory mechanisms. SNF1A and Dead SNF1A variants demonstrate selective effects on night activity and sleep, highlighting both kinase-dependent and independent roles in behavioral aging. The differential patterns observed between Elav(X) and Elav(II) further highlight the significance of a genomic perspective in functional studies.
Behavioral impacts of Ampk/SNF1A manipulation using panneuronal and muscle-specific drivers in Drosophila:
Using neuronal and mitochondrial drivers, we explore their role in energy regulation and neural function using Ampkα overexpression, Ampkα knockdown, Dead SNF1A overexpression, and SNF1A overexpression genes. This helps us understand how energy imbalance impacts movement and coordination. In our study, we did not observe any statistically significant difference in male (Figure 4a), female (Figure 4aI) Drosophila flight index at 3 and 6 weeks of age, individually with Elav (II) driver. With the Elav(X) driver, we have observed a statistically significant decrease in flight index of Ampkα knockdown Drosophila at 6-week-old compared to 3-week-old Drosophila in males (Figure 4b), and 6-week-old Ampkα overexpression, Dead SNF1A overexpression female Drosophila compared to 3-week-old Drosophila (Figure 4bI). While using Fln-Gal4;Mito-GFP, we observed wild-type control (w^1118^) and Ampkα knockdown Drosophila at 6-week-old-old showed reduced flight index compared to 3-week-old male Drosophila (Figure 4c). Whereas female Drosophila showed statistically significant reduction in wild-type control (w^1118^), Ampkα knockdown, and Dead SNF1A overexpression Drosophila of 6-week-old age compared to 3-week-old Drosophila, and Ampkα knockdown showed statistically significant reduction compared to 6-week-old control Drosophila (Figure 4cI). We then studied the geotaxis (climbing) behavior of these Drosophila with Elav(II) and Elav(X) drivers. In our study with Elav(II), we found no statistically significant difference in male (Figure 4d) and female (Figure 4dI) Drosophila at both 3- and 6-week-old age. Whereas with the Elav(X) driver, we observed that Dead SNF1A overexpression in 6-week-old females statistically significantly reduced compared to 3-week-old Drosophila (Figure 4eI), but no significance was observed in males (Figure 4e). Our results underline that Ampkα and SNF1A signaling influence age-related motor behaviors in a driver-, sex-, and tissue-specific manner. While the Elav(II) driver showed the least impact, Elav(X) and Fln-Gal4, Mito-GFP revealed statistically significant age-associated declines in flight and geotaxis in Drosophila. This implies that mitochondrial and neuronal energy imbalance compromises neuromuscular coordination during aging.
Panneuronal Ampkα regulation differentially impacts the lipid metabolism:
Since the behavioral responses are statistically significantly controlled by metabolic status and Ampkα expression along with age in Drosophila, we further tested the impact of panneuronal expression of Ampkα and its variants on lipid accumulation in the brain and head regions of the fly. Object count represents the number of lipid spots detected and the area represents the mean area of lipid spots in each brain section. Lipid objects count in head region data showed an increased lipid accumulation in 6-week-old Ampkα overexpression Drosophila, compared to age-matched wild-type control (w^1118^) and 3-week-old Ampkα overexpression Drosophila (Figure 5f). Whereas lipid object area increased in Ampkα overexpression at 3 weeks compared to wild-type control (w^1118^) Drosophila. At 6 weeks old, lipid object area was significantly reduced in Ampkα overexpression, Ampkα knockdown, Dead SNF1A overexpression, and SNF1A overexpression Drosophila compared to 6-week-old control Drosophila. We found a statistically significant increase in lipid object area in Ampkα overexpression compared to 3-week-old (Figure 5g). In the brain region at 3 weeks of age, we did not observe a statistically significant difference in lipid object count (accumulation) (Figure 5h), but lipid object area was higher than control Drosophila. At 6 weeks of age, statistically significantly less in Ampkα overexpression, Ampkα knockdown, Dead SNF1A overexpression, and SNF1A overexpression Drosophila compared to 6-week-old control and compared to 3-week-old Ampkα overexpression and Dead SNF1A overexpression Drosophila. We also found an increased lipid object area in wild-type control (w^1118^) at 6-week-old age compared to 3-week-old wild-type control (w^1118^) Drosophila. Our findings show that lipid metabolism is considerably changed by panneuronal regulation of Ampkα in an age-dependent way. Ampkα overexpression causes dynamic changes in lipid droplet size across the head and brain regions, as well as increased lipid accumulation in the head at older ages. The decreased lipid object area seen in several genotypes at 6-week-old implies that lipid homeostasis is disrupted by long-term disruption of Ampkα signaling, whether by overexpression or knockdown, which may reflect changed metabolic needs or poor energy mobilization in the aged brain. At the same time, our results support the idea that age-related changes in brain lipid storage can be caused by metabolic imbalance in neurons and identify Ampkα as a major regulator of lipid remodeling throughout aging.
Discussion
Aging is a complex biological process characterized by an increasing decline in physiological, behavioral, and cognitive capabilities. At the cellular level, age-related degeneration is closely linked to metabolic imbalance and mitochondrial dysfunction, both of which disrupt energy homeostasis and increase susceptibility to neurodegenerative diseases. Our findings show that targeted modulation of conserved metabolic and mitochondrial genes, predominantly in a tissue and age-dependent manner, has a statistically significant impact on behavioral phenotypes and lipid metabolism in Drosophila, a widely used model for studying aging and energy regulation. Metabolic regulation of sleep and activity: Panneuronal knockdown of genes involved in glycogen metabolism, mitochondrial function, and fatty acid oxidation, such as Ald, Gnmt, GlyP, and mAcon1 led to increased total and night sleep with reduced overall activity in mid- and late-age Drosophila, indicating an energy-conserving behavioral adaptation. These changes correspond to studies in mammalian models where mitochondrial stress and reduced ATP availability increase sleep drive and impair arousal (Anderson et al., 2022; O’Hearn, 2024). On the contrary, genes like Adsl, Ogdh, and Nmdmc showed increased sleep fragmentation and reduced activity, consistent with age-related sleep variability driven by compromised neuronal energy metabolism (Mander et al., 2017). These findings support the hypothesis that behavioral aging is severely regulated by neuronal metabolic state and suggest that sleep disturbances in aging could be caused by compromised bioenergetics.
Tissue-specific metabolic control of locomotor performance: Our work found that indirect flight muscle-specific knockdown of metabolic genes such as Ampkα, Ald, Adsl, mAcon1, and Ogdh resulted in more locomotor impairments. Knockdown of genes such as SdhD, Marf, and Gnmt leads to a decrease in flight performance, especially in 6 weeks with Elav(II)-Gal4, which demonstrates non-cell autonomous effects of these genes. Negative geotaxis with panneuronal knockdown of Adsl, Gnmt, SdhD, Marf genes showed reduced locomotor performance in an age-dependent manner, consolidating their non-cell autonomous role and neuro-muscular interaction. Impaired flight performance was seen in aged male Drosophila, whereas female Drosophila displayed gene- and age-dependent variability. This sex-specific sensitivity is consistent with results from fly and animal models that show sexually dimorphic mitochondrial responses to aging and metabolic stress (Fang et al., 2023; Videlier et al., 2019). Given the high energy demand of continuous flying, poor performance in these Drosophila could be attributed to inefficient mitochondrial ATP synthesis and altered lipid metabolism within muscle tissues, which supports comparable findings in aging rodent models (Joseph et al., 2012; Wedan et al., 2024).
Functional role of Ampkα and SNF1A signaling in behavioral aging: Our study examines how Ampkα and its yeast counterpart, SNF1A, function in neurons. Ampkα is an important energy sensor that turns on during metabolic stress, and its dysregulation has been linked to aging and neurodegeneration (Guo et al., 2023; Liu et al., 2012; Wang et al., 2019). Overexpression of Ampkα leads to decreased total sleep and increased fragmentation at 3-week-old, but increased night sleep and decreased fragmentation at 6-week-old, showing adaptive sleep behavior in response to improved neuronal energy sensing. Ampkα knockdown and SNF1A overexpression led to decreased activity and adjusted sleep patterns, implying their role in behavioral control. The distinct phenotypes observed between Elav(X) and Elav(II) drivers further highlight the significance of genomic context and insertion site effects in functional studies using the GAL4/UAS system. These findings are consistent with previous reports that neuronal AMPK activation improves sleep quality and protects against circadian disruption (Healy et al., 2021; Jordan and Lamia, 2013).
Motor behavior and mitochondrial stress response upon Ampkα modulation: Behavioral responses like flight and geotaxis were differentially influenced depending on the driver and gene manipulated. Elav(II) had the least impact, whereas Elav(X) and Fln-Gal4;Mito-GFP lines showed statistically significant flight deficits in older Drosophila, with Ampkα knockdown and Dead SNF1A overexpression. These results are associated with studies exhibiting that mitochondrial dysfunction and disturbed lipid metabolism decrease motor performance and resilience to stress in aging Drosophila and animal models (Haynes et al., 2024; Lima et al., 2022; Zhao et al., 2022). The use of IFM-specific driver (Fln-Gal4;Mito-GFP) further confirmed that peripheral IFM-specific energy dysregulation contributes to systemic aging phenotypes, possibly via cell-autonomous signaling.
Lipid remodeling and brain metabolism
Our result also establishes that panneuronal modulation of Ampkα substantially changes lipid accumulation in the fly brain and head regions. Overexpression and knockdown of Ampkα increased lipid droplet number and area at mid-age, but exhibited reduced size at older ages brain and head regions. These results suggest that both hyperactivation and inhibition of AMPK signaling interrupt lipid homeostasis, consistent with earlier studies reporting AMPK as a key regulator of lipid organization, synthesis, and mitochondrial biogenesis (Herzig and Shaw, 2018; Jeon, 2016; Reznick and Shulman, 2006). The detected age-related decline in lipid droplet size, particularly in the brain, may suggest a reduced lipid recycling or increased demand for fatty acid oxidation due to neuronal stress, a mechanism that promotes neurodegeneration (Mallick et al., 2024; Szrok-Jurga et al., 2023; Vesga-Jimenez et al., 2022). Together, these results provide a comprehensive functional map of how conserved metabolic and mitochondrial genes contribute to aging-related behavioral and physiological decline. Our data emphasize the idea that tissue-specific metabolic regulation plays a critical role in behavioral aging and that Ampkα functions as a molecular core integrating energy stress, sleep regulation, locomotion, and lipid metabolism. The functional specificity of individual genes across neuronal and muscular systems highlights the need for precision-targeted approaches in therapeutic development for age-associated disorders. We believe transcriptomic and metabolomic profiling of dissected tissues will allow a deeper understanding of downstream pathways altered by gene modulation. Mitochondrial functional assays (e.g., ATP levels, ROS production, membrane potential) should be incorporated to validate bioenergetic stress. Finally, incorporating rescue experiments or pharmacological modulation of AMPK could establish causal links and translational relevance, particularly for interventions aimed at mitigating age-related functional decline. Our findings emphasize the value of using Drosophila as a model for dissecting conserved metabolic mechanisms of aging and provide the basis for detecting therapeutic targets for age-associated behavioral impairments and neurodegeneration.
Conclusions
This study provides compelling evidence that tissue-specific changes in conserved metabolic and mitochondrial genes control age-related behavioral deterioration in Drosophila. Manipulation of these genes at the neuronal and muscle levels revealed different but overlapping effects on lipid balance, motor coordination, and sleep. Ampkα has been identified as a key regulator that integrates physiological outcomes and energy sensing across tissues and aging phases. These findings suggest potential biological targets for age-related functional loss and neurodegeneration and support a foundation for investigating metabolic treatments in aging.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Amorim J.A., Coppotelli G., Rolo A.P., Palmeira C.M., Ross J.M., Sinclair D.A., 2022. Mitochondrial and metabolic dysfunction in ageing and age-related diseases. Nat Rev Endocrinol 18, 243–258.35145250 10.1038/s 41574-021-00626-7PMC 9059418 · doi ↗ · pubmed ↗
- 2Anderson L., Camus M.F., Monteith K.M., Salminen T.S., Vale P.F., 2022. Variation in mitochondrial DNA affects locomotor activity and sleep in Drosophila melanogaster. Heredity (Edinb) 129, 225–232.35764697 10.1038/s 41437-022-00554-w PMC 9519576 · doi ↗ · pubmed ↗
- 3Bartman S., Coppotelli G., Ross J.M., 2024. Mitochondrial Dysfunction: A Key Player in Brain Aging and Diseases. Curr Issues Mol Biol 46, 1987–2026.38534746 10.3390/cimb 46030130 PMC 10969191 · doi ↗ · pubmed ↗
- 4Barwell T., De Veale B., Poirier L., Zheng J., Seroude F., Seroude L., 2017. Regulating the UAS/GAL 4 system in adult Drosophila with Tet-off GAL 80 transgenes. Peer J 5, e 4167.29259847 10.7717/peerj.4167 PMC 5733373 · doi ↗ · pubmed ↗
- 5Brischigliaro M., Fernandez-Vizarra E., Viscomi C., 2023. Mitochondrial Neurodegeneration: Lessons from Drosophila melanogaster Models. Biomolecules 13.10.3390/biom 13020378 PMC 995345136830747 · doi ↗ · pubmed ↗
- 6Burkewitz K., Zhang Y., Mair W.B., 2014. AMPK at the nexus of energetics and aging. Cell Metab 20, 10–25.24726383 10.1016/j.cmet.2014.03.002PMC 4287273 · doi ↗ · pubmed ↗
- 7Chatterjee N., Perrimon N., 2021. What fuels the fly: Energy metabolism in Drosophila and its application to the study of obesity and diabetes. Sci Adv 7.10.1126/sciadv.abg 4336 PMC 818958234108216 · doi ↗ · pubmed ↗
- 8Chaudhari S.N., Kipreos E.T., 2018. The Energy Maintenance Theory of Aging: Maintaining Energy Metabolism to Allow Longevity. Bioessays 40, e 1800005.29901833 10.1002/bies.201800005 PMC 6314662 · doi ↗ · pubmed ↗