Paricalcitol and hydroxychloroquine modulates extracellular matrix and enhance chemotherapy efficacy in pancreatic cancer
Bassel El-Rayes, Dhana Reddy, Sujith Sarvesh, Jeremy Foote, Doug Welsch, Chengde Cheng, Mehmet Akce, Ganji Purnachandra Nagaraju

TL;DR
Paricalcitol and hydroxychloroquine improve chemotherapy effectiveness in pancreatic cancer by modulating the extracellular matrix.
Contribution
The study identifies a novel mechanism involving ITGB4 inhibition and ECM modulation to enhance chemotherapy in pancreatic cancer.
Findings
PH combination with 5FU/oxaliplatin increased cell death and apoptosis in PDAC cell lines.
In vivo, PH combination therapy inhibited PDAC growth and activated T and NK cells.
PH reduced ECM proteins, particularly ITGB4, which was confirmed through genetic knockdown.
Abstract
Pancreatic ductal adenocarcinoma (PDAC) is a highly aggressive cancer with poor prognosis and limited therapeutic options. In a previous publication, our group defined some of the mechanisms that vitamin D analogue paricalcitol (P) and hydroxychloroquine (H) potentiated the effects of gemcitabine-based chemotherapy in PDAC. Based on this, we hypothesized that PH may potentiate 5-fluorouracil (5FU) and oxaliplatin-based chemotherapy, and this may involve a novel mechanism of extracellular matrix (ECM) modulation. The combination of PH with 5FU/oxaliplatin significantly increased the cell death, apoptosis, and S-phase cell cycle arrest as compared to untreated or 5FU/oxaliplatin treated MIA PaCa-2, HPAC and KPC cell lines. In vivo, the combination therapy inhibited PDAC growth and altered the immune landscape by activating T and NK cells. Proteomic analysis revealed significant reduction…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
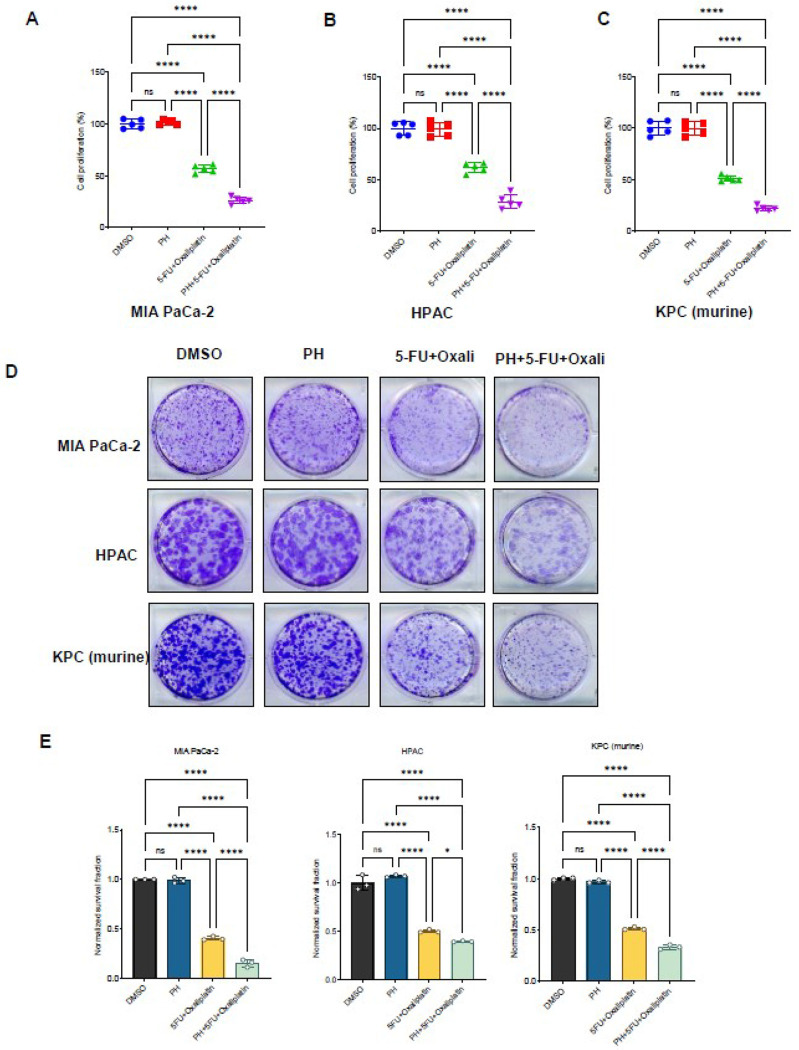 Figure 1
Figure 1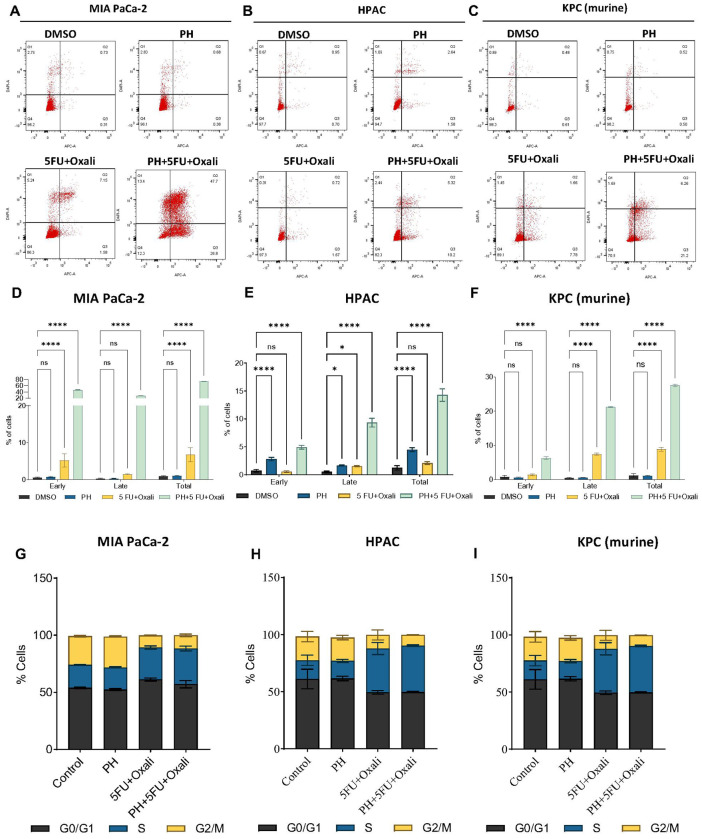 Figure 2
Figure 2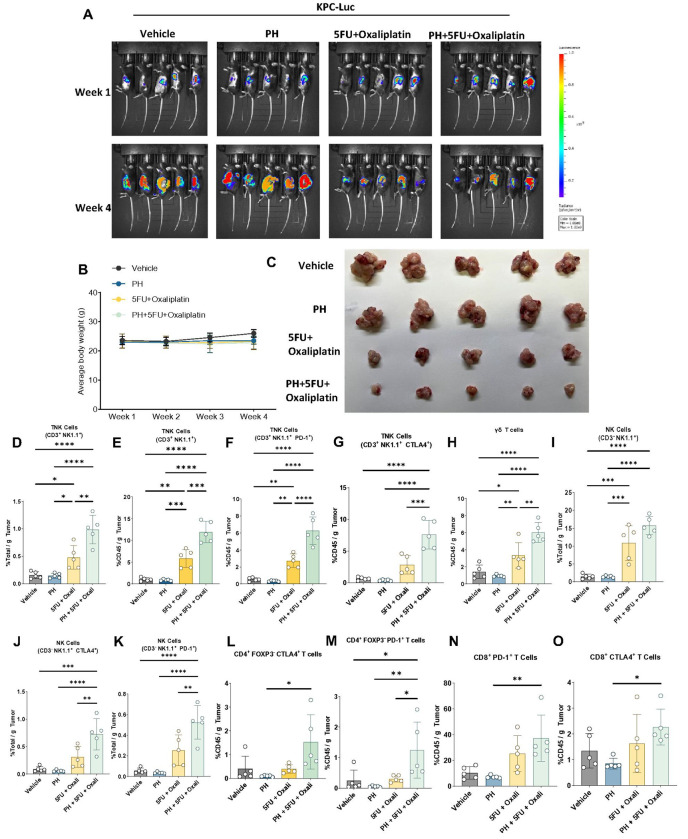 Figure 3
Figure 3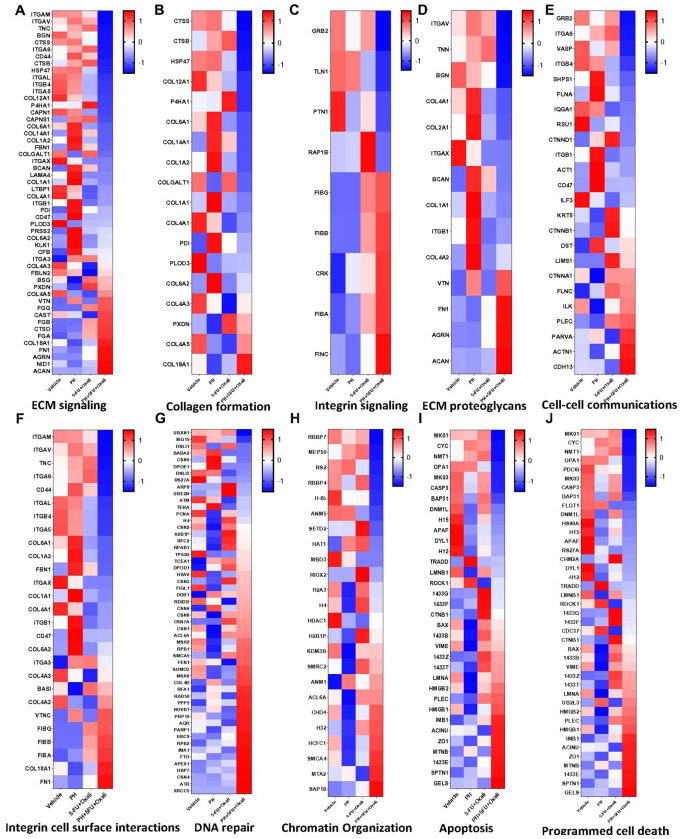 Figure 4
Figure 4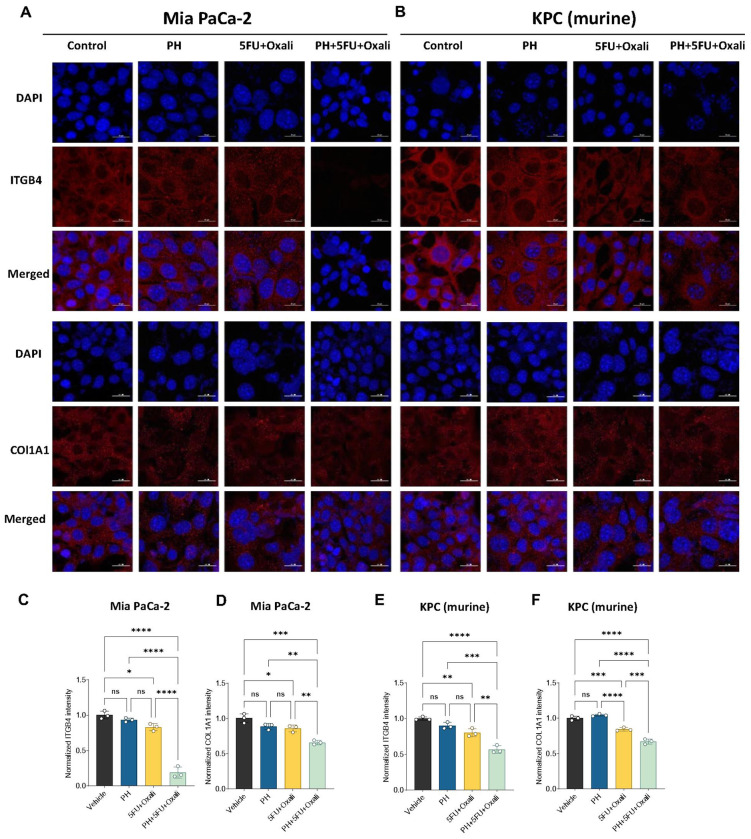 Figure 5
Figure 5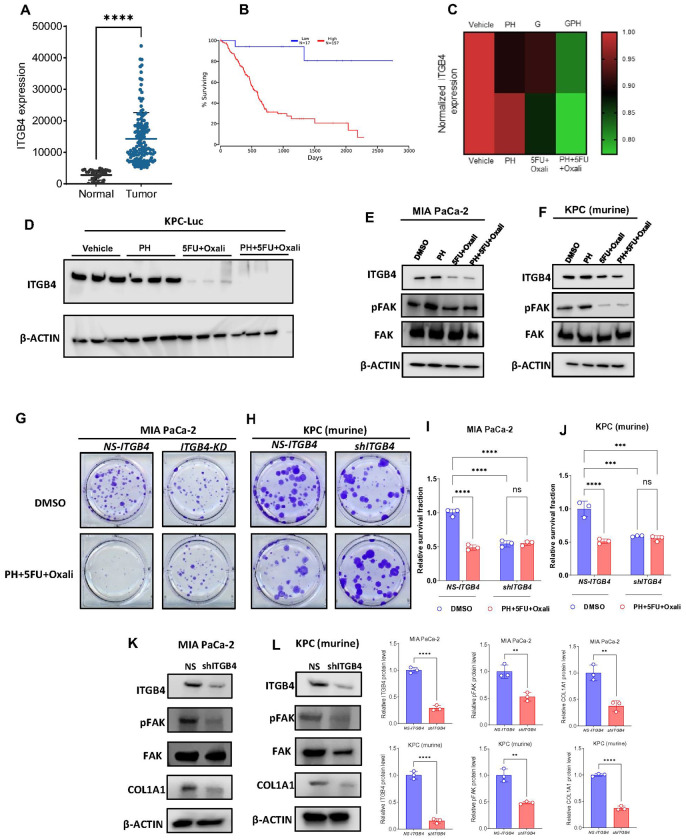 Figure 6
Figure 6- —University of Alabama at Birmingham, Birmingham, AL, USA
- —Comprehensive cancer center core support
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsPancreatic and Hepatic Oncology Research · Glioma Diagnosis and Treatment · Cancer, Lipids, and Metabolism
Introduction
Pancreatic ductal adenocarcinoma (PDAC) is one of the most deadly malignancies [1]. It is estimated that by 2030, PDAC will become second leading cause of cancer related deaths in the U.S. The 5-year survival rate for PDAC is poor (~ 11%) due to its tendency for early metastasis and resistance to systemic treatments including cytotoxic and immune based therapies [2, 3]. The resistance to systemic therapies is partly due to the presence of collagen-rich dense extracellular matrix (ECM) [4].
The PDAC ECM consists of several components such as collagens, glycoproteins, fibronectins, proteoglycans, hyaluronic acid, and laminins. Cancer associated fibroblasts (CAF) play a pivotal role in the development and maintenance of the ECM. PDAC cells interact with ECM components through integrins and receptor tyrosine kinases to form focal adhesion complexes, which help in growth, metastasis, and drug resistance [5, 6]. ECM affects the behavior of immune and angiogenic cells in the tumor microenvironment (TME) promoting a tumorigenic effects [7]. The compact and heterogeneous physical network of ECM components inhibit drug delivery further contributing to resistance and poor prognosis [8]. Therefore, the ECM has pleotropic effects in PDAC facilitating cancer cell survival, metastasis, and chemo-resistance. Modulating the ECM represents a rational approach to inhibit PDAC progression and promote the effects of systemic therapies.
Previous reports have indicated that vitamin D analogue such as paricalcitol (P) and hydroxychloroquine (H) have effects on CAF and immune TME. Vitamin D analogues has been shown to suppress activated CAF in PDAC. It has also been shown to suppress the expression of several ECM proteins such as smooth muscle alpha-actin (SMA) in breast cancer [9–12]. H selectively reduced the deposition of ECM in hepatic stellate cells by inhibiting α-SMA and collagen I [13]. H alleviates renal interstitial fibrosis by inhibiting PI3K/Akt signaling and attenuates ECM and EMT by suppressing NF-κB signaling [14]. H has also been shown in clinical trials to modulate the immune microenvironment in PDAC [15]. The effects of the combination of H and P with 5FU-based chemotherapy on modulation of ECM and immune microenvironment in PDAC has not been previously studied.
Clinical trial targeting ECM components including matrix metalloproteinase or hyaluronidase in PDAC have resulted in limited success [16]. The heterogeneity of CAF’s and redundant ECM composition and structure, limits the benefit of these therapies, which are very specifically designed against one component or pathway in the ECM. PDAC cells frequently develop resistance mechanisms, such as alterations in integrin expression and ECM remodeling, which reduce the effectiveness of such treatments. In our prior study, we reported the addition of HP to gemcitabine resulted in immune modulation and effects on autophagy [17]. These effects were observed in preclinical models and clinical samples from paired biopsies obtained from patients on a trial [17]. While gemcitabine based therapy is a standard chemotherapy for PDAC, accumulating evidences suggests that 5-FU based regimens may have shown superior efficacy [18]. Encouraged by the observed effects of PH with gemcitabine, we planned to assess this combination with 5FU-based therapy. In addition, our objective was to evaluate if the mechanisms involved in potentiation of 5FU-based therapy by H and P were mediated through mechanisms involving modulation of immune cells and ECM proteins in the TME.
Results
Paricalcitol and hydroxychloroquine enhances the growth inhibitory effects of 5FU and oxaliplatin in both murine and human PDAC cells
The effects of PH in combination with 5FU and oxaliplatin (oxali) on murine and human PDAC cells were evaluated by dosing the cells with varying concentrations of the drugs and then quantifying the survival rates by cell viability using MTT assay. Human PDAC cell lines MIA PaCa-2 and HPAC, and murine KPC PDAC cells were cultured in the presence of PH, 5FU/oxali either alone or in combination for 72 h (Fig. 1A–C). The combination treatment was extremely efficient in urging cytotoxicity in all the cell lines. The combination index (CI) values, calculated with PRISM software for dose-effect analysis, designated the presence of a robust synergism between 5FU/oxali with PH, exclusively at lower and intermediary drug doses compared to chemotherapy alone. We then asked whether this potential combination has effect on long-term survival of PDAC cells using clonogenic assays. Treatment of PH with 5FU (10 μM) and oxali (20 μM) inhibited colony-forming ability of PDAC cells (Fig. 1D and 1E).
PH with 5FU and oxaliplatin combination promotes apoptosis in PDAC cells
We analyzed the effect of this combination on inducing apoptosis in PDAC cells. Apoptosis measurement by annexin V and PI staining method. In MIA PaCa-2 cells, apoptosis was significantly higher in the combination arm (PH + 5FU + Oxali) compared to DMSO, PH, and 5FU/oxali alone. (Fig. 2A and 2D). Similar results were seen in HPAC (Fig. 2B and 2E) and KPC cells (Fig. 2C and 2F). Taken together these results prove that the combination treatment is significantly enhancing apoptotic effect in both human and murine PDAC cells highlighting potentiation of 5FU/oxali by PH in PDAC treatment comparing to existing treatment.
PH with 5FU and oxaliplatin combination inhibits cell cycle arrest in PDAC cells
To study the anti-proliferative mechanism of 5FU/oxali combined with PH, the effect of this combination treatment on the PDAC cell cycle was studied. Briefly, KPC, MIA PaCa-2 and HPAC cells were treated with 5FU (10 μM) and Oxaliplatin (20 μM), 0.35 μM of P and 25 μM of H alone or in combination for 72 h (Fig. 2G–I). 5FU/oxali alone or in combination with PH resulted in S phase arrest. This is an effect that did not occur in DMSO or PH treated cells. (Fig. 2G–I). The S phase arrest is due to effects of 5FU/oxali and is similar to effects previously reported [19, 20].
PH with 5FU and oxaliplatin combination more potently inhibits PDAC tumor growth
Since PH has effects on TME as well as cancer cells, orthotopic mouse model of PDAC were used to further evaluate this regimen. Mice were treated with vehicle, PH, 5FU/ oxali or their combination. The combination therapy significantly inhibited PDAC growth (Fig. 3A) compared to vehicle. Additionally, the combination therapy demonstrated significantly greater efficacy compared to either 5FU/oxali or PH alone. The body weight of all mouse groups was stable; suggesting all treatment were well tolerated (Fig. 3B). End-of-study KPC-Luc tumor pictures and weight confirmed the potentiation of 5FU/oxali by PH (Fig. 3C).
The combination of PH with 5FU and oxaliplatin alters immune landscape in the KPC-luc-TME
To assess the immune effects of this regimen, we assessed the quantities of T and NK cell subsets by multiparameter flow cytometry. The combination of PH with 5FU/oxali significantly activated T and NK cells population in total tumor well as in only CD45^+^ cells compared to vehicle, 5FU/oxali and PH. In addition, we noticed a significant increase in the proportions of CD3^+^ NK1.1^−^ PD-1^+^, and CD3^+^ NK1.1^−^ CTLA-4^+^ TNK cells suggesting that the combination therapy not only enhances the TNK cells activation but also promotes an immune environment favorable to antitumor responses in the KPC-luc TME (Fig. 3D–G). A substantial increase in the proportions of γδ T cells in the combination treatment compared to any other treatments was observed (Fig. 3H). Significant activation of NK cells in the combination treatment as compared to vehicle, PH and 5FU/oxali was observed (Fig. 3I). Increased expression of CD3^−^ NK1.1^+^ CTLA-4^+^ and CD3^−^ NK1.1^+^ PD-1^+^ NK cells in the combination treatment suggests a potential shift in the immune profile (Fig. 3J–K). Combination of PH with 5FU/oxali did not have any significant impact on the CD4^+^ and CD8^+^ T cells (Supplementary Fig. 2A-F). An increase in the expression of CD4^+^ FOXP3^−^ CTLA-4^+^ (Fig. 3L) and CD4^+^ FOXP3^−^ PD-1^+^ (Fig. 3M) (non-significant) in combination treatment was observed, suggesting the combination resulted in exhaustion and increase in regulatory capacity of CD4^+^ T cells. No change in the expression of CD8^+^ FOXP3^−^ PD-1^+^ (Fig. 3N) and CD8^+^ FOXP3^−^ CTLA-4^+^ (Fig. 3O) was observed in combination treatment.
The combination of PH with 5FU and oxaliplatin inhibits ECM formation through ITGB4
As the tumor ECM plays major role in PDAC progression, we then asked if the combination treatment has impact on ECM inhibition. To this end, we utilized mass spectrometry (MS)-based proteomics on mouse models of PDAC treated with DMSO, PH, 5FU/oxali, and combination. Since the matriosome includes several of ECM protein such as collagens, matrix metalloproteinase (MMP), glycoproteins, and proteoglycans, we were interested to uncover the changes on these proteins in combination treatments [5]. In our study, PH with 5FU/oxali treatment caused a decreased abundance of several ECM-related proteins. Inhibition in collagen formation proteins such as COL1A1, COL2A1, COL4A1, COL161, COL9A1, COL12A1, COL14A1, CTSB, HSP47, TNC, and BCAN was observed in the animals treated with the combination (Fig. 4A). This suggests changes in the tumor progression and alterations in the ECM remodeling and tumor stroma interactions. We further extended our analysis to check if this inhibition was mediated through integrin signaling and identified that several integrin signaling proteins along with collagen proteins were significantly inhibited in the combination treatment, including ITGB4, ITGAL, ITGAM, ITGA5, and ITGAV (Fig. 4B–C). To conform this, we performed a western blot from the tumor tissues and found that combination treatment significantly inhibits the expression of ITGB4 (Fig. 6D). This result points to a potential mechanism by which the combination treatment disrupts integrin-mediated pathways that are necessary for the ECM stability and tumor progression in PDAC. Examining the ECM proteoglycan signaling identified several collagen-binding proteins such as COL4A1, COL2A1, COL1A1 along with integrin’s (ITGB and ITGAV) were reduced in combination treatment (Fig. 4D). Results of cell-cell communication revealed reduction of GRB2, ITGB4, SHPS1, FLNA, RSU1, and CD47, which impairs cell proliferation, adhesion, migration, and immune evasion. Activation of CDH13, PARVA, ILK, and FLNC enhances cell adhesion, migration, and survival, potentially driving tissue remodeling (Fig. 4E).
Proteins such as ITGAM, ITGAV, ITGA6, ITGB4, ITGB1, ITGA5, and ITGA3, along with ECM-related genes like TNC, COL1A1, COL1A2, COL6A1, COL6A2, FBN1, COL4A1, ITGAX, CD47, and CD44, play critical roles in regulating cell-matrix interactions and ECM remodeling. The downregulation of the expression of these genes in the combination treatment arm likely impairs key cellular processes, including adhesion, migration, and signaling, potentially affecting tissue structure and cellular responses to the treatment (Fig. 4F). In combination treatment, the downregulation of the expression of various genes involved in DNA repair and signaling, including UBXN1, ISG15, DNLI1, BABA2, CSN5, DPOE1, DNLI3, RS27A, ARP8, UBE2N, ATM, TERA, PCNA, H4, and CSN2, points to a potential impairment in the DNA damage response and repair mechanisms (Fig. 4G). These proteins are critical for various aspects of DNA repair, including ubiquitination, DNA damage sensing, repair pathway activation, and cell cycle regulation. In addition, DNA repair pathways are key mechanisms of resistance to DAN damaging agents such as oxaliplatin. Combination treatment potently inhibited the expression of chromatin organization proteins such as RBBP7, MEP60, RBBP4, SETD2, and HAT1 (Fig. 4H). Reduced expression of apoptosis proteins like CASP3, Cytochrome C, TRADD, MAPK1, MAPK3, and APAF1 and activation of pro-apoptotic protein such as BAX were observed in the combination treatment confirming the apoptosis induction observed in the PDAC cell line experiments (Fig. 4I). Programmed cell death proteins such as CYC, NMT1, OPA1, CASP3, and APAF1 found to be downregulated in combination treatment (Fig. 4J).
Pharmacological and genetic suppression of ITGB4 inhibits PDAC growth and ECM formation by COL1A1
Next, we pursued to ascertain the mechanisms by which this combination treatment affects PDAC. We hypothesized that inhibition of ITGB4 expression contributed to the decrease in ECM markers overexpressed in PDAC. To further investigate this, we first performed immunofluorescence study and identified that, combination treatment significantly lowers the protein levels of ITGB4 and COL1A1 in both human (Fig. 5A, 5C–D) and murine PDAC cells (Fig. 5B, 5E–F) compared to DMSO, PH or 5FU/oxali. We then analyzed the expression of ITGB4 in publicly available data sets and identified that, ITGB4 is highly expressed in PDAC (Fig. 6A) and its overexpression is associated with poor patient survival [21] (Fig. 6B). We then looked at our previous studies where we have performed the proteomics in the same murine model using gemcitabine instead of 5FU and oxaliplatin [17]. Interestingly, we identified that, ITGB4 is significantly reduced in the combination of PH with gemcitabine compared to gemcitabine, PH or vehicle- treated alone (Fig. 6C). This suggests that PH is enhancing the efficacy of chemotherapy drugs by inhibiting the ECM formation through ITGB4. This hypothesis was supported by our findings that ITGB4 expression was significantly reduced in both gemcitabine and 5FU/oxali treatments in combination with PH. We then validated the inhibitory effect of these combinations by performing western blot on tumor tissues (Fig. 6D) and cultured cells treated with the same conditions as in vivo experiment (Fig. 6E–F) and identified the similar reduction of ITGB4 and COL1A1.
To directly assess, if the ECM disruption in combination treatment was mediated by ITGB4, we genetically knockdown ITGB4 and observed the expression of COL1A1. Interestingly, the knockdown of ITGB4 showed the reduced growth in colony formation assay suggesting that ITGB4 is crucial for PDAC growth in both human (Fig. 6G and 6I) and murine cells (Fig. 6H and 6J). Furthermore, immunoblotting results in murine and human PDAC cells with ITGB4 knockdown showed reduced expression of phospho FAK (pFAK) and COL1A1, which is consistent with the combination therapy suggesting that genetic and pharmacological inhibition of ITGB4 inhibits the PDAC growth and ECM disruption through COL1A1 (Fig. 6K–L). These results collectively prove that the combination therapy disrupts expression of key elements of ECM such as COL1A1 by downregulating ITGB4, which in contributes to PDAC growth inhibition.
Discussion
The addition of PH to 5FU/oxali in PDAC significantly increased growth inhibition similar to the results previously reported with gemcitabine [17]. Increase in apoptosis as measured by annexin V and PI staining supports this is an underlying mechanism of inhibition [22, 23]. In orthotopic model, the combination exhibited significantly enhanced tumor growth inhibition compared to single treatments. The proteomic data supports increase in apoptosis pathways in the animal model like the observation in the cell lines. The unchanged bodyweight across all treatment groups indicates that this treatment does not have an increased toxicity profile, which is similar to the observed effects in gemcitabine-based clinical trials. The mechanisms underlying the enhanced anti-tumor effects in the animal model include modulation of ECM, changes in tumor immune microenvironment as well as alterations in DNA repair mechanisms.
PDAC microenvironment is immunosuppressive contributing to the aggressive behavior and resistance to therapy [24–26]. One of the most important aspects of our study is that the combination treatment alters the immune landscape in PDAC microenvironment. The activation of T NK cells and increased proportions of γδ T cells suggests that this combination not only targets tumor cells but also modifies the immune responses as well. The observed changes in the increased expression of CD3^+^ NK1.1^−^ PD-1^+^ and CD3^+^ NK1.1^−^ CTLA-4^+^ TNK cells indicate a favorable shift in the immune environment and favors the potential for combining this regimen with immune checkpoint inhibitors. The absence of significant changes in the CD4^+^ and CD3^+^ T cells could be because these populations are coupled with the activation specific regulatory cells (FOXP3^+^ PD-1^+^), suggesting a complex interplay where the combination treatment may promote regulatory pathways that could be combined for anti-tumor response.
ITGB4 is encoded with β4 subunit, which is a laminin receptor that exclusively interacts with α6 subunit and plays a vital role in the biology of infiltrating cancer [27]. ITGB4 overexpression plays a key role in cell adhesion by binding with ECM proteins to regulate the cellular functions by transmitting adhesion signals [28]. ECM is vital for developing resistant niches by permitting cancer cells to tolerate therapeutic drugs before mutagenic resistance mechanisms are acquired [29]. Overexpression of ITGB4 shows significant correlations with tumor development and metastasis in several cancers including PDAC [30]. The molecular mechanism of this overexpressed ITGB4 in the ECM formation of PDAC is poorly understood.
Paricalcitol inhibits the expression of ITGA6, ITGB4, and α-SMA in human breast cancer cells [11]. H binds to collagens required for ECM formation [31]. H is a known autophagy inhibitor, and the autophagy is considered as important regulator of ECM deposition, fibrosis and tissue remodeling. Secretory autophagosmes also promotes ECM proteolysis [32]. The significant reduction in the ECM proteins such as collagens observed in our model supports our hypothesis that the combination treatment with PH disrupts the structural and biochemical ECM that supports that PDAC growth and metastasis. A candidate pathway that could underlie the observed ECM modulation is ITGB4. Increased expression of ITGB4 is correlated with EMT hallmarks, reduces the expression of E-cadherin, and increases the expression of vimentin [30, 33]. ITGB4 is also known to be a key molecule in the interaction between PDAC and ECM and contributes to ECM formation. Supporting this role of ITGB4 in PDAC biology is the observed poor survival in patients with PDAC with increased expression of ITGB4. In this current study, the significant downregulation of ITGB4 by PH treatment suggested that the downregulation of ECM components such as collagens or fibrinogen binding proteins maybe driven in part by ITGB4. Genetic inhibition of ITGB4 demonstrated similar ECM disruption and inhibitory effects on PDAC growth. Given this novel finding, we evaluated effects of PH and gemcitabine from our prior study on ITGB4 expression. Similarly, a significantly reduction in ITGB4 was observed with the gemcitabine-based regimen further confirming the role of this pathway in modulation of ECM observed after treatment with PH.
In conclusion, PH sensitizes PDAC to the effects of 5FU and oxali. PH has diverse effects on PDAC biology including immune modulation, autophagy, and inhibition of CAF. In addition, PH can modulate the ECM partly through ITGB4 which is a new mechanism of action of this regimen. The results observed in this study further support testing this regimen in future clinical trials.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Du J, Wang X, Li Y, Ren X, Zhou Y, Hu W DHA exhibits synergistic therapeutic efficacy with cisplatin to induce ferroptosis in pancreatic ductal adenocarcinoma via modulation of iron metabolism. Cell death & disease 2021; 12: 705.34262021 10.1038/s 41419-021-03996-y PMC 8280115 · doi ↗ · pubmed ↗
- 2Siegel RL, Giaquinto AN, Jemal A. Cancer statistics, 2024. CA: a cancer journal for clinicians 2024; 74.10.3322/caac.2182038230766 · doi ↗ · pubmed ↗
- 3Kung HC, Yu J. Targeted therapy for pancreatic ductal adenocarcinoma: Mechanisms and clinical study. Med Comm 2023; 4: e 216.36814688 10.1002/mco 2.216PMC 9939368 · doi ↗ · pubmed ↗
- 4Cid-Arregui A, Juarez V. Perspectives in the treatment of pancreatic adenocarcinoma. World journal of gastroenterology: WJG 2015; 21: 9297.26309356 10.3748/wjg.v 21.i 31.9297 PMC 4541382 · doi ↗ · pubmed ↗
- 5Tian C, Huang Y, Clauser KR, Rickelt S, Lau AN, Carr SA Suppression of pancreatic ductal adenocarcinoma growth and metastasis by fibrillar collagens produced selectively by tumor cells. Nature communications 2021; 12: 2328.10.1038/s 41467-021-22490-9PMC 805808833879793 · doi ↗ · pubmed ↗
- 6Jin S, Lee W-C, Aust D, Pilarsky C, Cordes N. β8 integrin mediates pancreatic cancer cell radiochemoresistance. Molecular Cancer Research 2019; 17: 2126–2138.31337672 10.1158/1541-7786.MCR-18-1352 · doi ↗ · pubmed ↗
- 7Tian C, Clauser KR, Öhlund D, Rickelt S, Huang Y, Gupta M Proteomic analyses of ECM during pancreatic ductal adenocarcinoma progression reveal different contributions by tumor and stromal cells. Proceedings of the National Academy of Sciences 2019; 116: 19609–19618.10.1073/pnas.1908626116 PMC 676524331484774 · doi ↗ · pubmed ↗
- 8Zinger A, Koren L, Adir O, Poley M, Alyan M, Yaari Z Collagenase nanoparticles enhance the penetration of drugs into pancreatic tumors. ACS nano 2019; 13: 11008–11021.31503443 10.1021/acsnano.9b 02395 PMC 6837877 · doi ↗ · pubmed ↗