Distinct Neural Mechanisms in the Propagation of Itch in the Face and Body
Santosh Mishra, Joshua Wheeler, Nidha Williams, Thierry Olivry

TL;DR
This study explores how itch is processed differently in the face and body, revealing distinct neural mechanisms that could lead to better targeted treatments.
Contribution
The paper identifies region-specific roles of Substance P and histamine receptor 1 in facial itch processing.
Findings
Substance P modulates itch in the cheek but not in the neck.
Histamine receptor 1 (HRH1) is the primary mediator of itch in the cheek, not pain.
Trigeminal ganglion neurons show increased co-expression of HRH1 and neuropeptides compared to dorsal root ganglion neurons.
Abstract
Itch intensity, quality and characteristics differ between body regions. Despite their prevalence, the mechanisms underlying regional itch differences remain poorly understood. Here, we investigate the neural basis of regional differences, focusing on the functional role of neuropeptides and histamine receptors expression in trigeminal ganglia versus dorsal root ganglia innervating afferents to cheek and the neck. Using an interdisciplinary approach, we identified distinct regional differences between the cheeks and necks of mice and humans. Our findings reveal that Substance P modulates itch in the cheek, while histamine receptor 1 (HRH1) is the primary receptor mediating itch but not pain in the cheek. We also discovered regional differences in sensory neuron subpopulations, with increased co-expression of HRH1 and neuropeptides in trigeminal ganglion neurons. Our study provides new…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
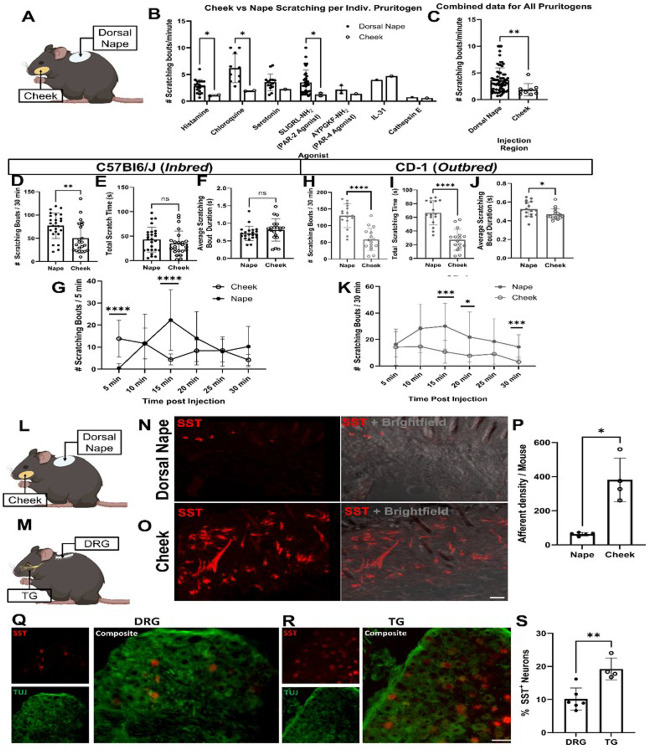 Figure 1
Figure 1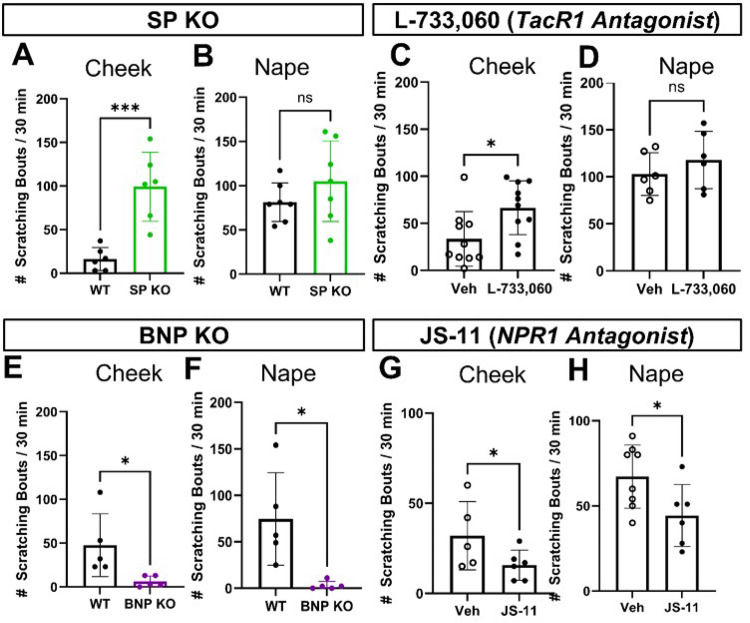 Figure 2
Figure 2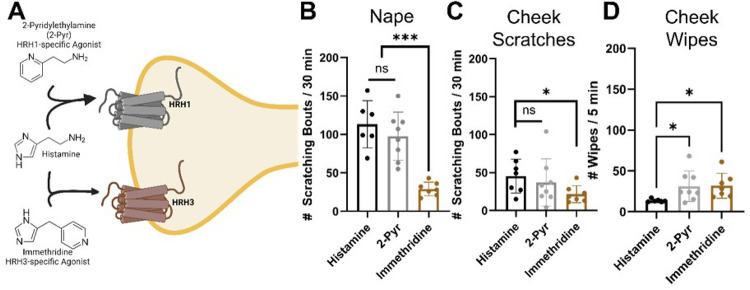 Figure 3
Figure 3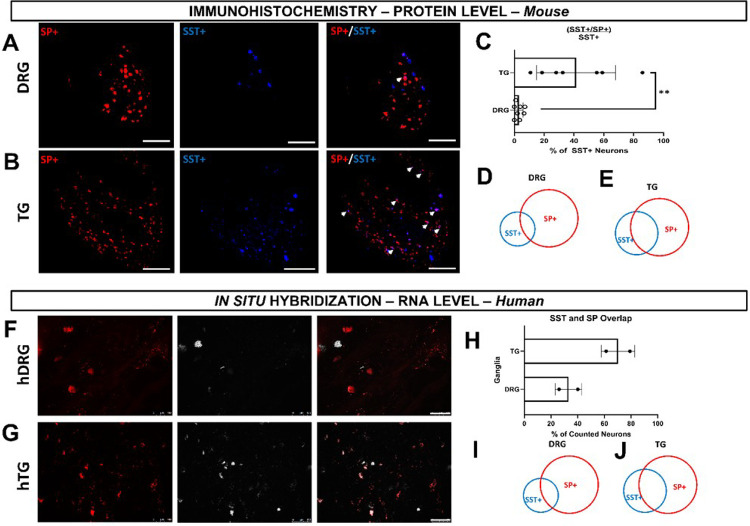 Figure 4
Figure 4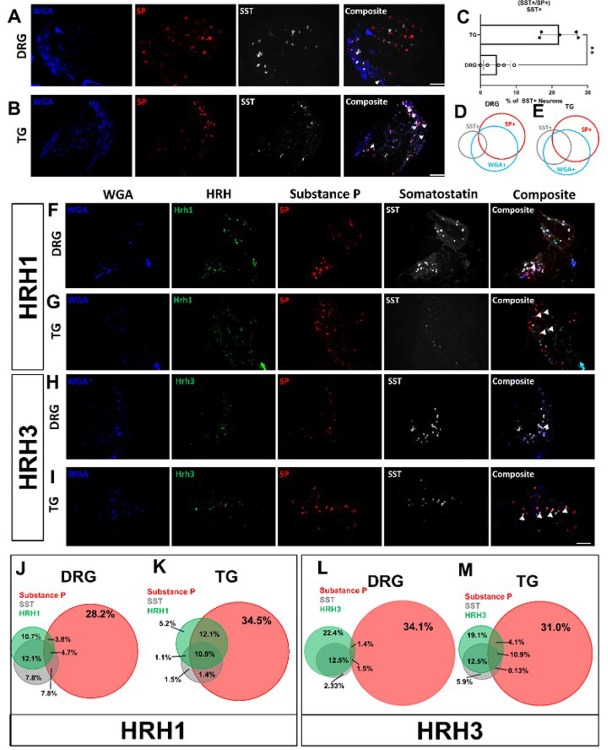 Figure 5
Figure 5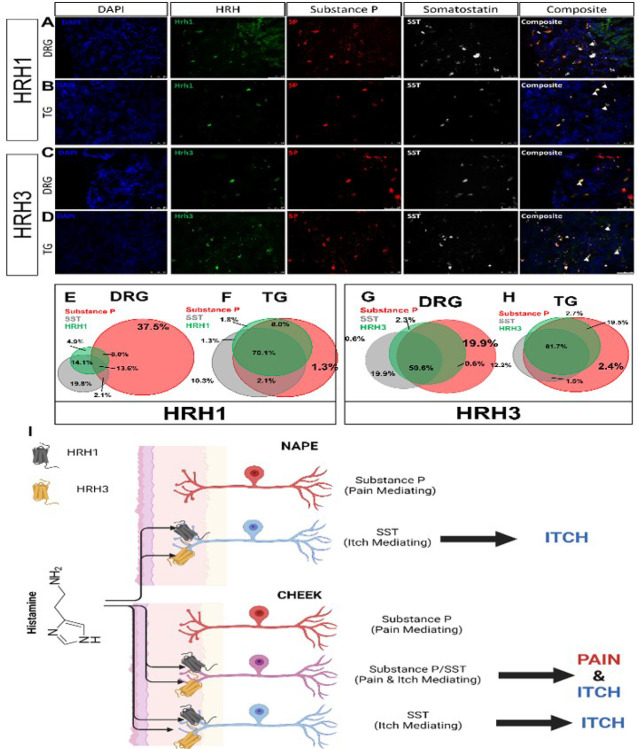 Figure 6
Figure 6Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsDermatology and Skin Diseases · Asthma and respiratory diseases · Stress Responses and Cortisol
INTRODUCTION
Somatosensory neurons in the dorsal root ganglia (DRG) and trigeminal ganglia (TG) detect various stimuli, generating distinct sensations and responses. DRG neurons innervate the skin of the limbs and torso, whereas TG neurons innervate unique sensory areas in the head and neck, including the eyes, nose, mouth, meninges, and vibrissae. Reports have identified genes linked to somatosensory neurons’ detection specificity in DRG and TG. DRG RNA-sequencing reveals distinct expression profiles for proprioceptive and itch-responsive neurons predominantly in mice^1,2^ and humans^3–5^. However, TG neuronal populations differ from DRG, as they lack proprioceptive neurons and innervate several unique sensory environments: the eyes, nasal passages and sinuses, meninges, and, in animals, vibrissae. Recent research has found TG-specific differences at the transcription level^4,6,7^, but whether that induces a differential behavioral response or differences in itch circuitry in mice is unknown.
In humans, the severity of itch is perceived differentially across body regions, with the face and torso^8–10^ exhibiting distinct intensity thresholds. Animal models have recapitulated these phenomena, showing fewer scratching bouts in response to intradermal pruritogen injections in the cheek than dorsal nape^11,12^. This difference is consistent across common pruritogens^11–19^, including histamine^11,12^. Notably, facial injection of pruritogens and algogens activates an overlapping population of second-order neurons found in the spinal trigeminal nucleus^15^. In contrast, itch and pain are processed in the DRG sensory neurons and activate separate populations of second-order neurons^19,20^.
The distinct neural processing of itch in the TG versus the DRG suggests the involvement of specific neuropeptides and signaling pathway involved in transmitting itch signals from the face versus the body. Itch and pain sensations are mediated through specific neuropeptides. Substance P (SP) mediates pain sensations^21^. Meanwhile, itch sensations are mediated by somatostatin (SST)-expressing neurons, which are > 99% co-localized with brain natriuretic peptide (BNP)-expressing neurons^20,22^. Activation of these first-order neurons triggers the release of these neuropeptides, which then activate second-order neurons in the dorsal horn of the spinal cord to further process these pain and itch sensations^19,20,22,23^.
The significant difference in itch processing between the face (cheek) and torso (neck) reveals a substantial knowledge gap. To address this, we employed a multifaceted approach combining mouse genetics and behavioral, pharmacological, cellular, and molecular techniques. Following intradermal histamine injections into the dorsal nape or cheek, we investigated itch behavior in C57Bl6/J (inbred) and CD-1 (outbred) mice. Additionally, we examined histamine-induced itch behavior in mice lacking BNP or SP as well as in mice treated with antagonists for their respective receptors (NPR1^24^ and TACR1). We also assessed nape and cheek skin innervation density by genetically marking somatostatin-expressing neurons with tdTomato. To identify differences in neuronal populations, we used tracing techniques followed by immunohistochemistry and in-situ hybridization to analyze histamine receptors H1 and H3, BNP/SST, and SP in human and mouse DRG and TG. Our findings show a crucial mechanism underlying the differences in itch processing between the face and body, which may have implications for future therapeutic interventions.
RESULTS
Regional scratching response between the face and neck
We initially performed a literature review of papers published between January 2009 and June 2023 (Fig. 1B, C) which revealed significant differences in scratching bouts, when normalized for time, between these two regions in response to various pruritogens, including for histamine (*p < 0.0001), chloroquine (*p = 0.0008), and SLIGRL-NH2, PAR-2 specific agonists (*p = 0.0006) (Fig. 1B). This data was further confirmed when we combined all scratching bouts/minute values for all pruritogens (*p = 0.0073) (Fig. 1C).
To investigate the regional scratching response between the face and neck, we comprehensively analyzed scratching behavior in C57Bl6/J mice following intradermal injections of pruritogens into the dorsal nape and cheek. Histamine elicited significantly fewer scratch bouts of ipsilateral cheek-directed scratching than dorsal nape-directed scratching bouts (*p = 0.0040) (Fig. 1D) in C57Bl6/J mice injected with histamine. In contrast, we did not find any significant differences when comparing the total amount of time spent scratching directed at these areas (*p = 0.2947) (Fig. 1E), and no significant differences were seen in the average bout duration (*p = 0.2540) (Fig. 1F). The time course of histamine-induced scratching bouts differed significantly between the dorsal nape and cheek (Fig. 1G). Ipsilateral cheek-directed scratching bouts peaked within the first 5 minutes (*p < 0.0001) (Fig. 1G) and the number of dorsal nape-directed scratching bouts peaked at 15 minutes (*p = 0.0001) (Fig. 1G).
To confirm that our findings in C57Bl6/J mice were not strain-specific, we repeated the experiments using CD-1 mice, a commonly used itch research outbred mouse line (Fig. 1H–K)^26^. Our results showed a similar pattern: fewer ipsilateral cheek-directed scratching bouts (*p < 0.0001); however, we found significantly shorter scratching times than dorsal nape-directed scratching bouts (*p < 0.0001) (Fig. 1H &I). Unlike with C57BL/6J mice, bout duration significantly differed between regions in the CD-1 mice (*p = 0.0380) (Fig. 1J). The time course of histamine-induced scratching bouts differed between regions, with cheek-directed scratching peaking within 5 minutes, and significant differences were observed at 15 (*p = 0.0002), 20 (*p = 0.0477) and 30 minutes (*p = 0.0009) (Fig. 1K).
Overall, our data suggests that regional differences in itch are conserved across outbred and inbred mouse lines, with the cheek being less responsive to histamine-induced itch compared to the dorsal nape.
Regional itch differences are not explained by innervation densities
Given the difference in histamine-induced itch between the dorsal nape and cheek, we hypothesized that there may be a difference in innervation density of itch sensing neurons between the facial dermatomes innervated by the TG^27^ and the dorsal nape dermatomes innervated by the DRG^28^. To assess this, we generated SST-cre::Ai9 mice in which tdTomato was conditionally expressed in SST-expressing neurons. We chose to generate SST-cre::Ai9 mice since previous work has demonstrated that SST-expressing DRG sensory neurons are responsible for mediating itch signals induced by histamine^20,29^. We found that the skin of the cheek has a significantly higher degree of innervation by SST-cre::Ai9 neurons as compared to the dorsal nape (*p = 0.0158) (Fig. 1N – O). Due to the increased innervation density of SST-cre::Ai9 afferents in the cheek, we hypothesized that there was an increase in the number of SST-expressing neurons in the TG. As expected, we found significantly more SST-expressing neurons in the TG as compared to the DRG (*p = 0.0029) (Fig. 2Q–S). This increase is likely to be responsible for increased SST-positive afferent density between the dorsal nape and the cheek. These findings also establish that the higher innervation density of itch-sensing neurons in the cheek does not correlate with the reduced scratching response observed in this region.
Distinct role of SP in modulating itch response in the cheek and dorsal nape.
Since innervation density did not explain the regional differences in itch response, we hypothesized that pain-associated neuropeptides, specifically Substance P (SP)^21^, might modulate ipsilateral cheek-directed scratching behavior. Based on previous observations where we found the dorsal nape, all SST-expressing DRG sensory neurons are positive for BNP^20^. Further, these BNP-expressing DRG sensory neurons are not positive for Substance P (SP)^22^, a canonical pain-associated neuropeptide.^21^ While there is literature about the relative co-expression of these neuropeptides in DRG sensory neurons, there is no information about their co-expression in TG sensory neurons.
To test if SP modulated itch responses in the cheek, we used knockout (KO) mice lacking SP or BNP and quantified scratching bouts after histamine injection into both the dorsal nape and the cheek (Fig. 2). We found that SP-KO mice exhibited significantly more ipsilateral cheek-directed scratching bouts than wild-type controls (*p = 0.0007) (Fig. 2A), with no differences in dorsal nape-directed scratching bouts (*p = 0.2390) (Fig. 2B). We used BNP-KO mice as a control, as these mice should lack an itch behavior phenotype, and demonstrated that the loss of BNP resulted in a significant decrease in the total number of dorsal nape (*p = 0.0126) and ipsilateral cheek-directed (*p = 0.0338) scratching bouts (Fig. 2E & F).
To confirm that our behavior results in SP-KO and BNP-KO mice, we pharmacologically inhibited the SP receptor (TacR1) with L-733,060^31,32^. We found that these mice exhibited significantly higher amounts of ipsilateral cheek-directed scratching bouts than vehicle-treated mice (*p = 0.0198) (Fig. 2C). Like the SP-KO mice, L-733,060 treated mice had no significant differences in dorsal nape-directed scratching bout compared to vehicle-treated mice (*p = 0.3568) (Fig. 2D). Mice treated with the NPR1 antagonist JS-11^33^ experienced significantly fewer scratching bouts in both the ipsilateral cheek (*p = 0.0338) and the dorsal nape (*p = 0.0396) (Fig. 2G & H). These findings suggest that SP plays distinct roles in modulating itch response in the cheek and dorsal nape, with SP inhibiting and BNP promoting itch in the cheek.
Histamine receptor-specific itch induction.
To investigate which histamine receptors, mediate histamine-induced itch in these regions, we built on our previous findings on regional differences in itch between the cheek and nape. We focused on the four histamine receptors, three of which are expressed in DRG sensory neurons. scRNA-sequencing data showed that HRH1 mRNA is expressed predominantly in SST- and MrgprA3-positive DRG sensory neurons, while HRH3 is expressed in SP-positive DRG sensory neurons involved in pain^2^.
To determine which histamine receptor(s) are responsible for transducing histamine-induced itch, we injected 2-pyridylethylamine (HRH1-specific agonist)^34^ and immethridine (HRH3-specific agonist)^35^ into the dorsal nape and cheek of mice. We found that 2-pyridylethylamine induced HRH1-mediated itch in both the dorsal nape and ipsilateral cheek, with no significant differences in scratching bouts compared to histamine-injected mice (*p = 0.5208, nape; *p = 0.7708, cheek) (Fig. 3B & C). We found that 2-Pyr induced significantly more cheek wipes as compared to histamine in addition to the cheek directed scratching (*p = 0.0230) (Fig. 3D). Immethridine also induced more ipsilateral cheek wipes (*p = 0.136) (Fig. 3D), but fewer scratching bouts than histamine (*p < 0.0001, nape; *p = 0.0264, cheek) in both regions. Immethridine induced fewer scratching bouts than 2-pyridylethylamine in the nape (*p = 0.0002, nape), but not the cheek (*p = 0.2373). Our results suggest that HRH1 is the primary histamine receptor mediating itch in dorsal nape-innervating DRG sensory neurons, while HRH3 may play a role in pain-behavior.
Regional variations in sensory neuron populations underlying itch and pain
Our previous findings on regional differences in itch and SST/SP innervation led us to investigate the populations of SST-, SP-, HRH1-, and HRH3-expressing sensory neurons in the dorsal root ganglion (DRG) and trigeminal ganglion (TG) of mice, which we also further validated in humans. Our SST-cre::Ai9 innervation results indicated that there should be a larger proportion of SST-positive TG sensory neurons. Additionally, our behavior results indirectly indicate that SP-positive TG sensory neurons are important in modulating ipsilateral cheek itch. Since pain signaling is capable of modulating itch sensation and vice versa^19^, we hypothesized that there is a change in the number of TG sensory neurons that are positive for both SST and SP. We found that overall, there was a significant increase in the percentage of TG sensory neurons positive for both SST and SP as compared to the DRG (*p = 0.0012) (Fig. 4A – D). We further validated this in human using in situ hybridization (ISH; Fig. 4E–H).
Using wheat germ agglutinin (WGA-350) as a neuronal tracer, used here to identify skin-innervating sensory neurons, we also found a larger population of cheek skin-innervating TG sensory neurons positive for both SST and SP compared to dorsal nape-innervating DRG sensory neurons (*p = 0.0051) (Fig. 5). Next, we investigated the expression of HRH1 and HRH3 in skin-innervating sensory neurons in mice (Fig. 5). Our results showed an increase in the percentage of TG sensory neurons expressing HRH1, SST, and SP, as well as HRH3, SST, and SP, compared to the DRG. We further corroborated these findings in human TG versus DRG regional-specific differences in human sensory neurons (Fig. 6).
DISCUSSION
Our study reveals significant regional differences in itch response between the cheek and dorsal nape (Fig. 1). These differences are not attributed to variations in innervation density (Fig. 1), but to distinct populations of sensory neurons expressing specific neuropeptides and receptors (Fig. 2–3). Here, we demonstrate that the difference in facial histaminergic itch processing is due to differences in the proportion of TG neurons that co-express Substance P with SST, HRH1, and HRH3 (Figs. 4 &5). Behaviorally, this results in the likely activation of HRH1, inducing pain responses in the cheek. Further, we demonstrate that HRH3 might transduce histamine activation into pain (Fig. 3). Our histological results were also recapitulated in human DRG and TG samples (Figs. 4 & 6), extending their translational relevance. Finally, our results indicate that the TG processes facial itches fundamentally differently by a mechanism similar to what is seen in the gate control model of itch and pain processing (Fig. 6I).
Here, we confirmed that the number of histamine-induced ipsilateral cheek-directed scratching bouts is significantly reduced as compared to the number of histamine-induced dorsal nape scratching bouts by systematically comparing the scratching bouts by using the C57Bl/6J mice and the same amount of histamine in the same injection volume for both the cheek and nape injections. These results confirm that the reduction in the total number of ipsilateral cheek-directed scratching bouts is due to a difference in the neurophysiology of the cheek skin as opposed to differences in injection volumes or dose (at least for histamine). Interestingly, outbred CD-1 mice, which are thought to be a better model for human research^36^, had significantly fewer ipsilateral cheek-directed scratching bouts, corroborating the results from the inbred line. Furthermore, CD-1 mice have significantly shorter scratching bout durations (Fig. 1I), implying that they also perceive histaminergic itch in the cheek as less intense than histaminergic itch in the dorsal nape. Validation of regional differences in histamine-induced itch between inbred and outbred lines suggests that the study findings are not limited to a specific mouse strain and likely translatable to humans.
One of the limitations of our study is that we specifically investigated the role of histamine in regional itch differences, despite literature suggesting that other itch mediators, such as chloroquine, serotonin, SLIGRL-NH2, IL31, and proteases, may also exhibit similar regional differences (Fig. 1). There are mostly three reasons for this focus: Firstly, histamine is a well-known endogenous mediator for itch. Further, its involvement in itch processing is a helpful model for understanding the neural mechanisms underlying regional differences itch sensitivity. Secondly, in sensory neurons, histamine’s-induced itch primarily mediated by the histamine receptor 1 (HRH1). This receptor specificity allows for targeted investigation of histamine’s role in regional itch differences, without the confounding effects of other itch mediators. Lastly, histamine allows for a more in-depth investigation of the neural mechanisms underlying regional itch differences, which can serve as a foundation for future studies on other itch mediators, and the clinical relevance of histamine in itch disorders makes it a prime target for therapeutic intervention at sensory neurons carrying message to the CNS.
Histamine-induced behavior hints at TG-specific processing of itch behavior
Reports have indicated differences between the DRG and TG at the transcriptome level^3,4,6^. Yet, the overall assumption has been that itch and pain processing in the TG follows the same pathways as itch processing in the DRG. Electrophysiology work performed by Akiyama et al.,^15^ further hinted at a difference in pain and itch signaling when they reported that pruritogens and algogens, itch inducers and pain inducers, respectively, results in postsynaptic activation of an overlapping population of neurons in the spinal trigeminal (TG) nucleus.
There are currently two main theories about how the nervous system processes itch and pain. The first is a modified label-line theory, which posits that itch and pain are processed by two parallel pathways by the peripheral nervous system and the spinal cord dependent on the input simulus^37,38^. Overall, the label line theory is correct for the DRG; SST-positive/BNP-positive neurons primarily process itch, and pain is mediated mainly by SP-expressing DRG neurons^21,22^. Further supporting this theory is the finding that activation of the MrgprA3-expressing DRG neurons by capsaicin in TRPV1-KO::MrgprA3^ROSA − TRPV1^ mice, wherein TRPV1 is only in MrgprA3-expressing neurons, results in itch – not pain^18^. The second theory of itch and pain processing is the Gate Control theory, which states that separate pain and itch-responding DRG neurons activate the same set of secondary neurons located in the dorsal horn of the spinal cord. This theory was founded by discovery of dynorphin-expressing spinal interneurons activated following TRPV1 or TRPM8 neurons activation^19^. Activation of these dynorphin-expressing interneurons triggers the release of dynorphin, which then suppresses activation of gastrin-related peptide receptor (GRPR)-expressing spinal interneurons, thus preventing the transmission of itch signals^19^. Current research has determined that these GRPR-expressing spinal interneurons are where itch signals converge^19,20,22,23,39,40^ before these interneurons transmit itch signals to the parabrachial nucleus^41,42^. This theory is further supported by the finding that SST-dependent activation of SSTR2-subpopulation inhibits pain responses^20^. Both of these theories on itch processing have been assumed to be the de facto pathways by which the TG processes itch behavior despite the lack of evidence, either for or against, the presence of these pathways in the TG nucleus.
Our results suggest that the TG itself uses gate control to process facial itch signals despite the unclear presence of these pathways in the spinal trigeminal nucleus^43^. Our mouse data demonstrate that TG have a significantly increased percentage of SST-expressing neurons expressing Substance P compared to the DRG (Fig. 4). HRH1, which transduces histamine receptor activation into itch, has significant changes in downstream neuropeptide populations (Figs. 5 & 6). In the DRG, HRH1 is predominantly co-expressed with SST, with few neurons being positive for HRH1 and Substance P; however, in the TG, this proportion of HRH1-positive and Substance P-positive neurons roughly triples (Fig. 5). This change in HRH1 and Substance P expression also results in a change in behavioral outcomes (Fig. 3). HRH3 underwent similar changes in the proportion of neurons positive for Substance P in the TG compared to the DRG (Figs. 5 & 6). Interestingly, immethridine, an HRH3-specific agonist^35^, did not induce itch but significantly more ipsilateral cheek-directed wipes than histamine, indicating that histaminergic itch is both receptor-dependent and neuropeptide-dependent. These findings provide a basis for examining how the gate control theory of pain and itch processing may be relevant in this context. The gate control theory of pain and itch processing has evidence that there might be two subtypes: the leaky gate subtype suggesting itch and weak pain signals activate the same pathway^44^, and an intensity coding subtype suggests itch signal transmission is dependent on a frequency or burst of stimulation^45^. While we could not test this hypothesis here, but our results, and those reported in Akiyama et al^15^, alluding to an intensity coding in the TG. Another avenue to explore would be to determine if the histamine-triggered SP-release can activate a previously identified monosynaptic facial pain circuit^46^ as these neurons express the receptor for SP^47^. Activation of this pathway by histamine would also likely account for the decrease in itch behavior and increase in pain following intradermal injection of histamine into the cheek.
In summary, we identified distinct sensory neuron subpopulations, highlighting the importance of considering regional differences in itch response when designing treatment strategies. Overall, our study has significant implications for the understanding and treatment of itch disorders by developing therapies targeting HRH1 and HRH3 receptors that may provide new treatment options for itch disorders.
METHODS
Chemicals
Histamine was ordered from Sigma Aldrich (Catalog # H7125). Wheat Germ Agglutinin conjugated to Alexa Fluor 350 was ordered from Invitrogen (Catalog # W11263). L-733,060 (Catalog # 1145), 2-Pyrdylethylamine (Catalog # 2478), and immethridine (Catalog # 2315) were ordered from Tocris. JS-11 (1-cyclohexyl-3-(cyclopropylmethyl)-N-((3-methylisoxazol-5-yl)methyl)-2-oxo-2,3-dihydro-1H-benzo[d]imidazole-5-carboxamide) was custom synthesized by ChemBridge.
Animals
All mice were housed at NC State University and all experiments were IACUC approved (protocols 22–167-B, 19 −047-B, and 16–038-B). Mice were given food and water ad libitum and kept on a 12-hour light dark cycle (6am −6pm EST).
All inbred mouse lines were bred in-house from mice obtained from The Jackson Laboratory or the NIH. The following lines were purchased from the Jackson Laboratory: C57Bl6/J (Strain # 000664), B6.Cg-Tac1^tm1Bbm^/J (Substance P (SP) KO) (Strain # 004103)^21^, Sst^tm2.1(cre)Zjh^/J (SST-cre) (Strain # 013044)^49^, and B6.Cg-Gt(ROSA)26Sor^tm9(CAG0tdTomato)Hze^/J (Ai9) (Strain # 007909)^50^. BNP KO mice were a generous gift from Dr. Mark Hoon (NIH)^22^. Crl:CD1(ICR) (CD-1) mice were ordered from Charles River laboratories (Strain # 022). SST-cre::Ai9 mice were generated by crossing the SST-cre and Ai9 lines.
Itch Behavior
Histamine injection: All mice received intradermal injections of 100 μg/20 μL histamine in sterile 1X phosphate buffered saline (1X PBS). This concentration and volume were used for all intradermal injections. Immethridine and 2-Pyridylethylamine were intradermally injected with the same 100 μg/20 μL concentration. Mice were injected in either Dorsal Nape^11^ or Cheek^12^. L-733,060 was dissolved in sterile 1X PBS and 50 μL of a 1mg/kg solution was administered intravenously 30 minutes prior to histamine injection^31,32^. JS-11 was dissolved in DMSO, and the injection solution consisted of 10% Tween-80, 20% DMSO, and 70% Normal saline. A total of 16.3 μg/50 μL of JS-11 was administered intraperitoneally 30 minutes prior to histamine injection^33^.
The same mice were used for both the Cheek and Nape tests. For drug administration injections, mice were randomly assigned to receive Vehicle, L733–060, orJS-11. Mice were then randomized to the injection site. After a 10-day washout period, mice were treated with vehicle were then treated with JS-11 or L-733,060 and mice treated with JS-11 or L-733,060 were treated with vehicle.
Immunohistochemistry
For Tuj antibody staining, DRG and TG were isolated from SST-cre::Ai9 mice, fixed in 4% PFA for 24 hours at 4°C, cryoprotected using a 30% sucrose solution incubation (~ 72 hrs at 4°C), and then frozen in OTC media. Sections were cut at 14 μm and placed in 4% PFA for 15 minutes at room temperature. Slides were then blocked with solution of 5% BSA with 0.1% Triton-X100. Rabbit Anti-mouse Tuj (Abcam, Catalog # ab18207) (1:500 in 5% blocking solution) was applied to slides, which were then incubated overnight at 4°C. After 16–24 hrs, the slides were washed in ice cold 1X PBS trice and then incubated for 45 minutes at room temperature with Alexa Fluor 488 conjugated goat anti-rabbit IgG (Thermofisher, catalog # A-11008). Samples were images on a Leica DM500B epifluorescent microscope.
Skin Innervation Tracing
Mice were briefly anesthetized with isoflurane prior to injection of wheat germ agglutinin (WGA) that was conjugated to Alexa Fluor 350 (Invitrogen, Catalog # W11263) (WGA-350) to determine changes in the population sizes of skin-innervating DRG and TG neurons^52^. WGA-350 was injected intradermally into both cheeks and the dorsal nape. Both TG and C3-T2 DRG were isolated 24 hours after injection^52^. TG and DRG were then placed in OCT media and quickly frozen on dry ice and were stored at −80°C for up to 3 months. Cheek injections were bilateral to obtain the greatest potential number of TG sensory neurons that innervate the skin.
Fluorescent In Situ Hybridization
In situ hybridization was carried out using the RNAscope Multiplex Fluorescent Reagent Kit v2 (ACD, Catalog # 323100). DRG and TG were isolated from 8-week-old mice 24 hours after receiving 50 μL of a 1mg/mL solution of Alexa Flour-conjugated Wheat Germ Agglutinin (WGA-350) and frozen in OTC media at −80°C within 30 minutes. These sections were cut at thickness of 10 μm and placed onto Superfrost Gold Plus Slides (Fisher, cat. # 15-188-48). After sectioning, sections were air dried at −20°C for 2 hours. After this, slides were placed in 4% paraformaldehyde (4% PFA) made with DEPC water (Thermofisher, Catalog # 750023) at 4°C for 15 minutes. Slides were then washed twice in 1X PBS made with DEPC water. Following this wash step, slides were dehydrated at room temperature in 50% ethanol for 5 minutes, 70% ethanol for 5 minutes, and two washes in 100% ethanol for 5 minutes. The 50% ethanol and 70% ethanol solutions were made with DEPC water. The slides were then kept at −20°C in 100% ethanol overnight. Ethanol was purchased from Millipore Sigma (Catalog # 459836–2L).
The following day, the slides were removed from 100% ethanol and allowed to air-dry at room temperature for 5 minutes. A hydrophobic barrier was drawn around the samples using a hydrophobic barrier pen (ACD, Catalog # 310018). The hydrophobic barrier was allowed to dry for 5 minutes. After drying, roughly 5 drops of RNAscope hydrogen peroxide was added to fill the area within the hydrophobic barrier on the slide and incubated for 10 minutes at room temperature. The slides were then washed in DEPC treated water. After this wash, roughly 5 drops of RNAscope Protease III was added to fill the area within the hydrophobic barrier and incubated for 30 minutes at room temperature. The slides were then washed twice in 1X PBS made with DEPC water. Sections were hybridized and on the same day; therefore, sodium citrate was not used.
For hybridization, probes the following probes were ordered from ACD: Human Tac1 (Substance P) (Catalog# 310711-C3), human SST (Catalog # 310591-C2), human HRH1 (Catalog # 416501), human HRH3 (Catalog # 402191), mouse Tac1 (Catalog # 410351-C2), mouse SST (Catalog # 404631-C3), mouse Hrh1 (Catalog #491141), and mouse Hrh3 (Catalog # 428481). All DRG and TG slides were stained with the corresponding species-specific probes (human or mouse). All slides were stained with SST and Tac1, and either Hrh1 or Hrh3. To hybridize the probes, the hydrophobic barrier area was filled with ~ 6 drops of the appropriate probe mixture and then incubated for 2 hours at 40°C in a HybEZ oven. After this incubation, slides were washed in 1X wash buffer twice for two minutes (each wash). For the first two amplification steps, ~ 6 drops of RNAscope Multiplex FL v2 Amp1 or Amp2 was added to the area within the hydrophobic barrier and incubated for 30 minutes at 40°C. For the third hybridization step, RNAscope Multiplex v2 Amp3 was added to the area within the hydrophobic barrier and incubated for 15 minutes at 40°C. After each hybridization step slides were washed twice in 1X was buffer for 2 minutes at room temperature.
To develop the HRP signal the following steps were performed: roughly 6 drops of RNAscope Multiplex FL v2 HRP-C1, HRP-C2, or HRP-C3 were added to the area within the hydrophobic barrier on the slide and incubated for 15 minutes at 40°C. Slides were then washed twice for 2 minutes in 1X wash buffer at room temperature. To the area in the hydrophobic barrier 200 μL of Opal 520 (channel 1) (Akoya Biosciences, Catalog # FP1487001KT), Opal 570 (channel 2) (Akoya Biosciences, Catalog # FP1488001KT), or Opal 690 (channel 3) (Akoya Biosciences, Catalog # FP1497001KT) was added and the slides were incubated at 40°C for 30 minutes. All dyes were diluted 1:1000. After incubation, slides were washed twice in 1X wash buffer for 2 minutes at room temperature. Finally, roughly 6 drops of RNAscope Multiplex FL v2 HRP blocker was added to the area within the hydrophobic barrier and incubated at 15 minutes at 40°C. After this incubation, the slides were washed twice in 1X wash buffer for 2 minutes at room temperature.
Cell Counting and Quantification:
Images were collected on a Leica DMB500 epifluorescence microscope. Cells were counted based on appearance since DRG and TG neurons have distinctly round to avoid shapes with a large round central gap where the nucleus is located. Skin innervating neurons were normalized to the total number of WGA-350 neurons. In Figs. 5, 8, and 9 cell populations were normalized to the total number of cells counted. In Fig. 2, the percentage of SST-expressing neurons was normalized to the total number of TUJ positive neurons counted.
Statistical Analysis:
Data was analyzed in Graphpad Prism. Paired 2-tailed t-tests were performed to compare histamine-induced scratching in the cheek versus dorsal nape. Region-based data was analyzed with a 2-way ANOVA with a post hoc Holm – Šídák test for multiple comparisons. Other data was compared with a 1-way ANOVA with a post hoc Holm – Šídák test for multiple comparisons. Literature data was compared using an unpaired 2-tailed Mann-Whitney U-test. For all tests the initial significance value was set to α = 0.05.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Goswami S. C. Molecular signatures of mouse TRPV 1-lineage neurons revealed by RNA-Seq transcriptome analysis. J. Pain 15, 1338–1359 (2014).25281809 10.1016/j.jpain.2014.09.010PMC 5469214 · doi ↗ · pubmed ↗
- 2Unbiased classification of sensory neuron types by large-scale single-cell RNA sequencing | Nature Neuroscience. https://www.nature.com/articles/nn.3881.10.1038/nn.388125420068 · doi ↗ · pubmed ↗
- 3Goswami S. C. Itch-associated peptides: RNA-Seq and bioinformatic analysis of natriuretic precursor peptide B and gastrin releasing peptide in dorsal root and trigeminal ganglia, and the spinal cord. Mol. Pain 10, 44 (2014).25123163 10.1186/1744-8069-10-44PMC 4132360 · doi ↗ · pubmed ↗
- 4Flegel C. RNA-Seq Analysis of Human Trigeminal and Dorsal Root Ganglia with a Focus on Chemoreceptors. Plo S One 10, e 0128951 (2015).26070209 10.1371/journal.pone.0128951 PMC 4466559 · doi ↗ · pubmed ↗
- 5Ray P. Comparative transcriptome profiling of the human and mouse dorsal root ganglia: an RNA-seq-based resource for pain and sensory neuroscience research. Pain 159, 1325–1345 (2018).29561359 10.1097/j.pain.0000000000001217 PMC 6008200 · doi ↗ · pubmed ↗
- 6Manteniotis S. Comprehensive RNA-Seq Expression Analysis of Sensory Ganglia with a Focus on Ion Channels and GPC Rs in Trigeminal Ganglia. PLOS ONE 8, e 79523 (2013).24260241 10.1371/journal.pone.0079523 PMC 3832644 · doi ↗ · pubmed ↗
- 7Nguyen M. Q., Wu Y., Bonilla L. S., von Buchholtz L. J. & Ryba N. J. P. Diversity amongst trigeminal neurons revealed by high throughput single cell sequencing. Plo S One 12, e 0185543 (2017).28957441 10.1371/journal.pone.0185543 PMC 5619795 · doi ↗ · pubmed ↗
- 8Rukwied R., Zeck S., Schmelz M. & Mc Glone F. Sensitivity of human scalp skin to pruritic stimuli investigated by intradermal microdialysis in vivo. J. Am. Acad. Dermatol. 47, 245–250 (2002).12140471 10.1067/mjd.2002.120461 · doi ↗ · pubmed ↗