Closed State Structure of the Pore Revealed by Uncoupled Shaker K+ Channel
Francisco Bezanilla, Yichen Liu, Carlos Bassetto, Gustavo Contreras, Eduardo Perozo

TL;DR
Scientists discovered a new closed state structure of the Shaker potassium channel using a modified version that reveals how the channel's pore closes.
Contribution
The study reveals a novel closed pore structure of the Shaker K+ channel through an uncoupling mutation and cryoEM.
Findings
The uncoupled I384R mutant Shaker channel shows a fully closed pore with activated voltage sensors.
Pore closure involves a 'roll and turn' movement of S6 helices, differing from traditional models.
A noncanonical selectivity filter conformation was observed at G446, distinct from known states.
Abstract
Voltage gated potassium (Kv) channels play key roles in physiological processes, from cellular excitability to immune response and are among the most important pharmaceutical targets1. Despite recent advances in the structural determination of Kv channels, the closed state structure of strictly coupled Kv1 family remains elusive. Here, we captured the structure of Shaker potassium channel with a closed pore by uncoupling its voltage sensor domains from the pore domains. Structural determination of the uncoupled I384R mutant by single particle cryoEM revealed a fully closed pore in the presence of activated, non-relaxed voltage sensors. Putative conformational transitions estimated from a fully open pore domain indicates a “roll and turn” movement along the length of the pore-forming S6 helices, in sharp contrast to canonical gating models based on limited movements of S6 2–4. These…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
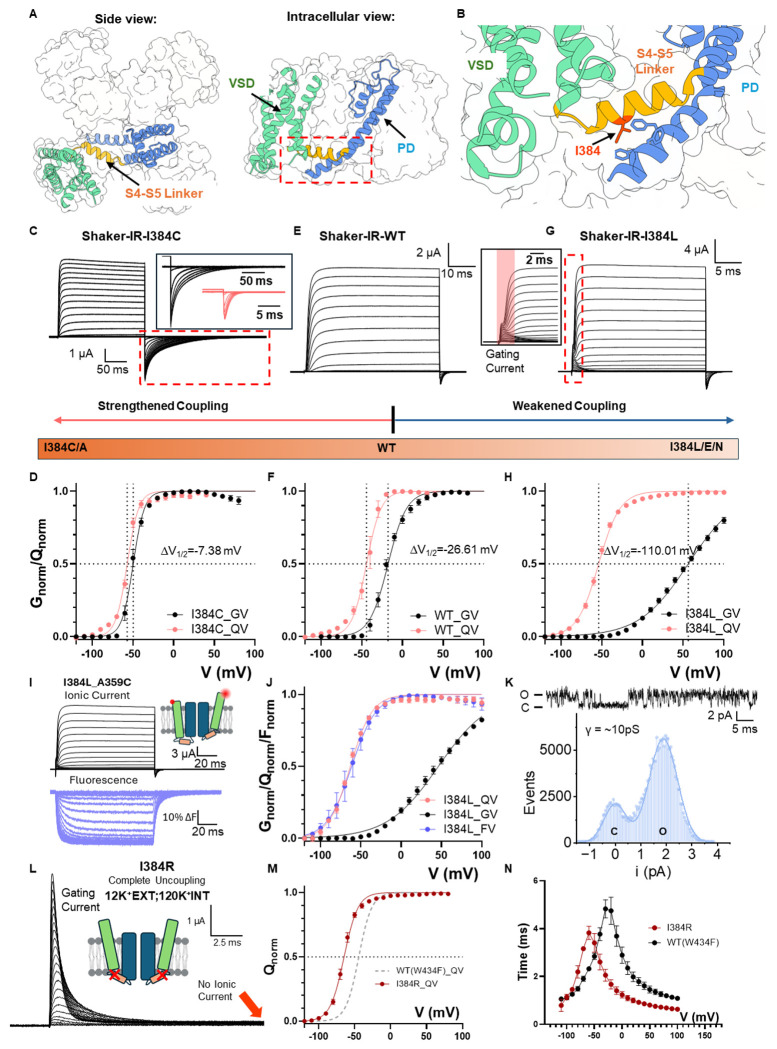 Figure 1
Figure 1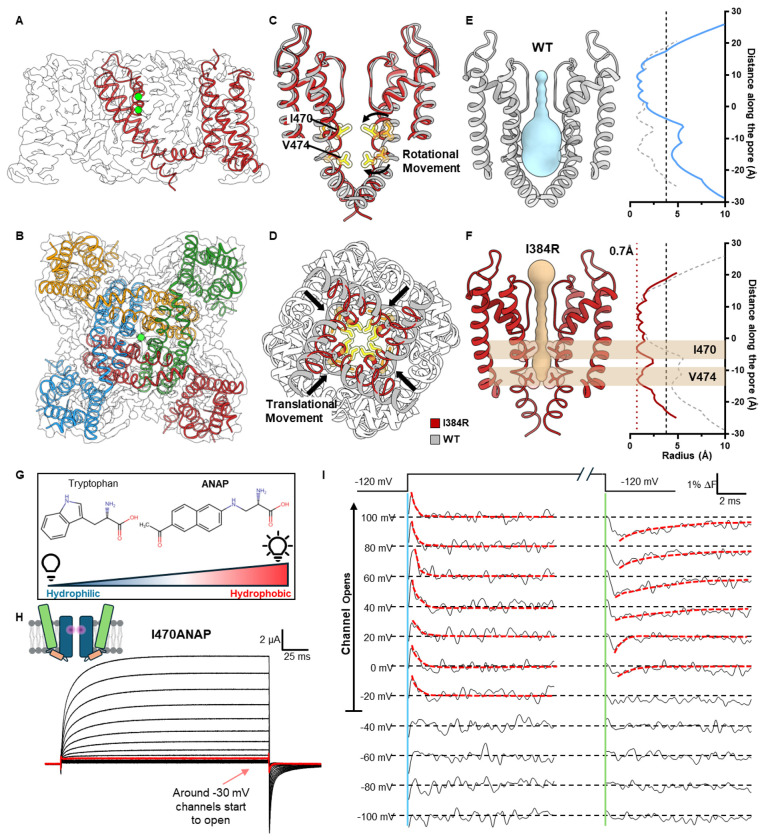 Figure 2
Figure 2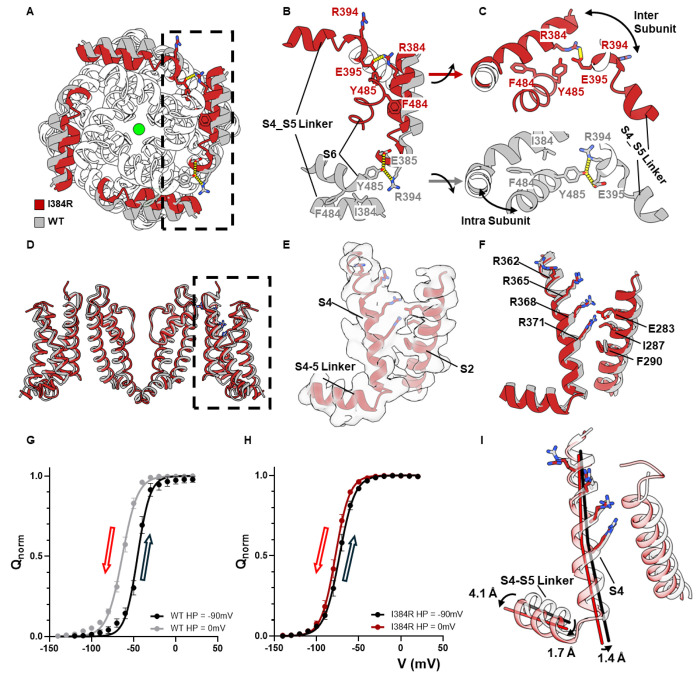 Figure 3
Figure 3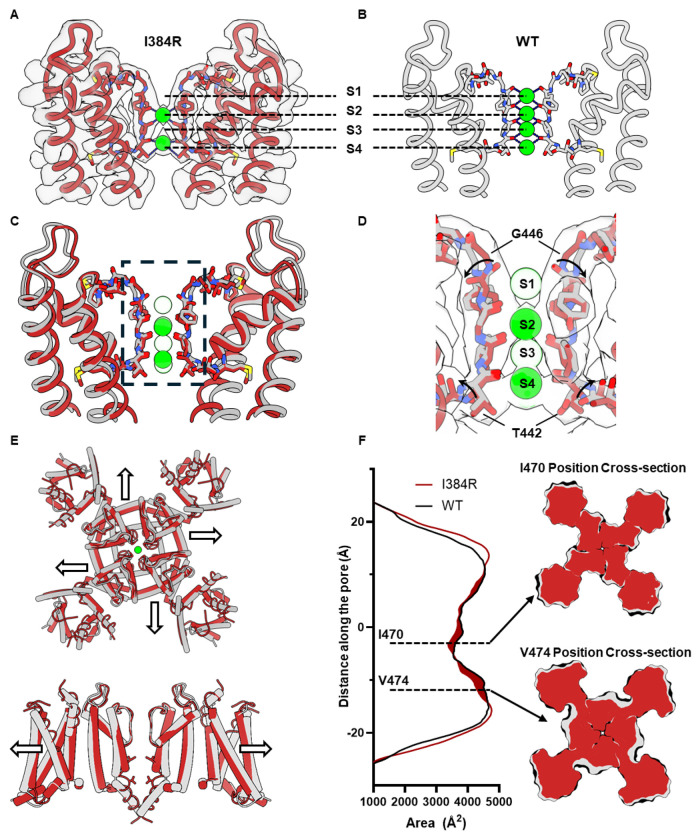 Figure 4
Figure 4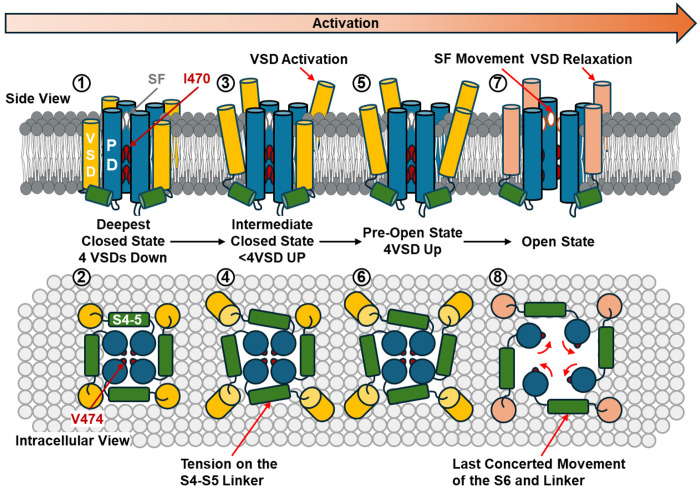 Figure 5
Figure 5Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsNanopore and Nanochannel Transport Studies · Ion channel regulation and function · Lipid Membrane Structure and Behavior
Structural studies on a variety of Kv channels have provided a great deal of information on the conformational landscape of the channels as a whole. However, the vast majority of the structures correspond to activated or inactivated conformations, since they were obtained in the nominal absence of an electric field. In “strictly coupled” Kv channels, the electromechanical coupling (EMC) between the voltage sensing domain (VSD) and the pore domain (PD) is obligatory: channel opening or closure requires activation or deactivation of the voltage sensors, respectively ^7–9^. This is in contrast to a typical allosteric coupling, as in BK ^10^, HCN ^11^ or hERG ^12,13^ channels for instance. While this tight coupling ensures a remarkably low “leak” potassium conductance at rest, it has also hindered the structural understanding of the gating mechanisms, since at 0 mV the voltage sensors typically populate the activated (Up) conformation, with the pore domain displaying an open inner bundle gate. As a result, the current gating model of strictly gated Kv channels, such as the Kv1 and Kv2 families, relies primarily on bacteria potassium channel structures ^2^ and electrophysiological data ^3,4^.
A conserved isoleucine I384 in the S4-S5 linker region was identified as pivotal for the electromechanical coupling in Kv1 family ^14^. Here, we report the closed structure of the pore in a strictly coupled Kv1 channel, Shaker K^+^ channel, by disrupting the coupling between the pore and the voltage sensors from mutations at I384. Single channel, ionic current, gating current and fluorescence measurements demonstrated that mutations to the side chain of I384 directly affect the strength of electromechanical coupling. In the most extreme case of I384R, the VSDs become completely uncoupled from the PD: the pore remained closed within a large range of voltages (−120mV to 180mV) while the VSDs activate/deactivate independently. Single-particle cryo-EM structure of the uncoupled channel revealed a collapsed permeation pathway, consistent with a fully closed conformation. Site-directed fluorimetry measurements utilizing a fluorescent unnatural amino acid (UAA) strongly supports our structural observations. Structural rearrangements were also observed in the selectivity filter (SF) and the voltage sensors likely underlying the structural basis for activation-inactivation coupling and hysteresis of the VSDs, respectively. Modifying the canonical “hinge” model, we propose a “roll and turn” gating model for strictly coupled Kv channels and a molecular mechanism of interactions among different conformational states.
Conserved isoleucine controls electromechanical coupling in Kv1 family
A systematic mutagenesis survey of the S4-S5 linker, a region that has been demonstrated to be important for EMC ^15–17^, led to the identification of a single residue that is able to tune the coupling strength between VSDs and PD. The conserved isoleucine 384 ^14^ is located at the N-terminus end of the S4-S5 linker and forms elaborate interactions with the intracellular end of the pore-forming S6 helix ^18^ (Fig. 1A, B). Replacing I384 with smaller residues such as alanine or cysteine, strengthened the already efficient electromechanical coupling (Fig. 1C and Extended Data Fig. 1). I384C, for instance, activated in more negative voltages with much steeper voltage dependency, as illustrated by its conductance-voltage (GV) curve. More importantly, channel activation closely followed the voltage sensor movement, measured in the gating-charge-voltage (QV) curve (Fig. 1D). While in wild type (WT) channels the difference between the V_1/2_ for QV (measured with the nonconductive W434F mutant ^19^ and GV curve was > 25mV (Fig. 1E, F, Extended Data Fig. 1, and Extended Data Tables 1–2), that difference was less than 7.5mV in I384C, a hallmark for strengthened electromechanical coupling. On the other hand, mutating I384 to glutamate, asparagine or leucine led to severe uncoupling between the VSD and PD (Extended Data Fig. 1). This is characterized, in the case of I384L (Fig. 1G), by large GV curve shifts (> 70mV) to more depolarized potentials, when compared to WT Shaker, together with a shallower slope in the GV curve (Fig. 1H). Strikingly, while the GV was shifted to the right, the QV curve of I384L was shifted in the opposite direction (Fig. 1H). Indeed, similar changes in the current kinetics and GV curves were also seen in hKv1.2 and hKv1.3 with mutations at equivalent position (I316 position in hKv1.2 and I386 position in hKv1.3), suggesting that I384’s critical role in the electromechanical coupling is conserved among strictly coupled Kv channels (Extended Data Fig. 2).
Given the large V_1/2_ gap between GV and QV in I384L, gating currents were easily resolved in the presence of ionic currents (Fig. 1G inset). To confirm that no additional slow component in the VSD movements was present in I384L that is responsible for the opening of the pore, tetramethyl rhodamine (TMR) was introduced at A359C in the extracellular loops of the VSD so that the movement of the voltage sensors would be evaluated via its fluorescence signal ^20–22^. The fluorescence records showed no additional component in the VSD movement, and the voltage-dependent fluorescence (FV) curve fully overlaps on the QV curve (Fig. 1I, J). This result demonstrates that our gating current measurement reflects the true movement of VSDs. To confirm that the observed effects were indeed a consequence of impaired coupling rather than due to changes in single channel properties, we performed noise analysis and single channel recordings ^23,24^ in I384L mutant. Noise analysis demonstrated that at 195mV, the maximum opening probability was less than 70% (Extended Data Fig. 3) and single channel recordings showed that the unitary conductance level in I384L (Fig. 1K, ~ 10 pS, Extended Data Tables 3–4) was similar to the WT (~ 12 pS) ^24^, but channel flickering was increased (Fig. 1K and Extended Data Fig. 3).
While in I384L, the impairment in the coupling was severe, we discovered another mutant that completely uncouples the voltage sensors from the pore, I384R. Despite the presence of K^+^ ions, I384R showed no ionic conduction, and only gating current could be recorded (Fig. 1L). The gating current itself activated at more hyperpolarized voltages compared to WT (Fig. 1M) and was kinetically faster compared to the WT with W434F background (Fig. 1N), as if an energetic load was taken off from the voltage sensors. This is also consistent with left-shifted QV curves seen in other uncoupled mutants such as I384N, I384L and I384E (Extended Data Fig. 1, and Extended Data Tables 1–2). In I384R, our recordings showed no discernible K^+^ current from − 120mV to + 180mV, demonstrating the exceptional structural stability of the closed pore. Additionally, a left-shifted QV curve allows the VSDs to transition more easily to the up conformation. As a result, at 0mV, I384R possesses a stably closed pore and four activated VSDs with little structural heterogeneity. This represents an appealing target for structural investigation.
Closed state structure revealed by a completely uncoupled channel
We expressed, purified and solved the structure of Shaker-IR-I384R, by single-particle cryo-EM. The structure was globally resolved to 3.5 Å, with clear densities for all the transmembrane helices (Fig. 2A, B, Extended Data Figs. 4 and 5). Similar to previously determined structures, the uncoupled channel assembled as a domain-swapped homotetramer (Fig. 2A, B) ^18^. However, unlike the strictly coupled WT channel structure, captured with an open pore, the uncoupled channel clearly displayed a collapsed permeation pathway. (Fig. 2C, D). Compared to the open conformation of Shaker, the pore-forming S6 helices in the closed pore underwent a “roll and turn” movement, where a translational movement brings the backbone of the S6 helices closer together (Fig. 2C), and a rotational movement places hydrophobic residues I470 and V474 directly into the pore. These form the two narrowest points of the closed pore (Fig. 2D). Pore radius calculations showed a closed permeation pathway with radius less than ~1 Å ^25^, leaving ion conduction an impossibility (Fig. 2E, F). Unexpectedly, the rotating-in of the I470 residue led to a total collapse of the water-filled inner cavity underneath the selectivity filter (Fig. 2E, F). Previous gating models predict a more limited “hinge-like” movement for the channel activation where a kink is created in the middle of the S6 helices around the conserved PVP motif and the intracellular half of the helices crosses or separates to close or open the channel ^26–31^.
To validate our structural observations, we set out to measure the local conformational changes around the position 470 in the WT channel utilizing a fluorescent unnatural amino acid probe. ANAP is comparable in size to a tryptophan, and its fluorescence changes according to the hydrophobicity of its local environment (Fig. 2G) ^32,33^. Utilizing the amber stop codon suppression method ^34^, we incorporated ANAP in a site-specific manner at the 470 position and recorded the ionic current and fluorescence signal simultaneously from I470ANAP (Fig. 2H, I). As channels opened, a fast transient fluorescence change was observed at the start of the depolarizing pulse and a slower one at the beginning of the hyperpolarizing pulse (Fig. 2I, highlighted with dashed line). The transient nature of the fluorescence signal seems to suggest a rotational movement where the different environments are sampled before reaching its final state, consistent with our structural observations. In the negative voltage range (< −40 mV), where the channel does not open, no fast transient fluorescent signal was observed, demonstrating that the fluorescence signal at I470 is only observed when the channel opens. These results are fully consistent with the idea that the structure of the uncoupled channel most likely represents a true closed state of the pore and suggest interactions between the bundle crossing region and the selectivity filter.
A tripartite interaction pocket essential for Electromechanical Coupling
Globally, I384R does not cause a kink in the S4-S5 linker or a local movement at the “elbow” region as was seen before in the resting bacteria Nav channels structures ^35^. Instead, we see a lateral, translational movement along the whole length of the S4-S5 linker towards the pore (Fig. 3A). Closer inspection revealed a tripartite interaction pocket among the N-terminus end of the S4-S5 linker, S6 helix in the same subunit and the C-terminus end the S4-S5 linker helix from the adjacent subunit (Fig. 3B). In the strictly coupled WT structure, I384 was securely lodged in a hydrophobic pocket formed by F484 and Y485 within the same subunit in the S6 helix (Fig. 3B – gray color). The hydroxyl group of Y485, on the other hand, interacted intimately with R394 and E395 in the S4-S5 linker from the adjacent subunit, establishing a structural coupling between the pore and the S4-S5 linker (Fig. 3C – gray color). In the uncoupled I384R structure, however, these interactions are all abolished. The full positive charge of the introduced arginine at position 384 forces itself out of the hydrophobic pocket where the side chain swings towards the adjacent S4-S5 linker. This conformational rearrangement pushes the adjacent R394, facing towards the pore previously, away from the S6 helix and allows for the formation of a salt bridge between R384 and E395 (Fig. 3B, C – red color). These newly formed interactions allows for a closer interaction among the S4-S5 linkers, creating a tight collar around the S5 and S6 segments stabilizing the closed state, and abolish the previous interactions with Y485 in the S6 helices. The observered disruption of the tripartite interactions likely underlies the structural basis for the uncoupling mechanism of I384R, similar to what was shown physiologically elsewhere ^36,37^.
Voltage-sensing domain captured in its activated but not relaxed state
Gating current measurements demonstrated that at 0mV, the QV curve of I384R had reached its maximum, suggesting that all the voltage sensors had activated. However, since there are multiple intermediate states for the VSDs, the electrophysiological data cannot unequivocally define whether the voltage sensors reach the fully activated state in the uncoupled channel or even if they move in a similar way as WT channels. To address this, we compared the VSD structures in I384R and the WT (Fig. 3D). Clear densities were resolved for all gating charges in S4 (R362, R365, R368, R371) ^38,39^ as well as the key residues that form the hydrophobic plug in S2 (I287, F290) ^40–42^ and the countercharge (E283) (Fig. 3E). The I384R structure shows that all four gating charges have moved pass the hydrophobic plug in the uncoupled conformation and are accessible to the extracellular solution (Fig. 3F), as seen in the WT structure. This suggests that in the uncoupled channel, voltage sensors move similarly to the WT channels to reach the fully activated state.
However, unlike the WT, voltage sensors in I384R do not appear to enter the relaxed state, a conformation that has been observed in Kv, Nav, Cav channels and voltage sensitive phosphatases (VSP) and is driven by prolonged depolarization ^43–48^. In WT channels with the W434F background, holding the channels at 0 mV for extended periods of time shifts the QV curve almost 20 mV to more negative potentials when compared to holding at −90 mV (Fig. 3G). This relaxed state was not observed in the uncoupled channel. Holding I384R at 0mV for > 30 s did not cause significant shifts in the QV curve, suggesting that the voltage sensors in I384R did not enter in the relaxed state, at least at this voltage (Fig. 3H). Since the cryo-EM structures were captured at 0 mV, the physiological evidence would then argue that the structure of VSDs in the open channel represented the relaxed state while in the uncoupled channel, the VSDs likely resided in the non-relaxed state. Structurally speaking, the major difference between VSDs in the WT and I384R lies mostly in the lateral displacement of the S4 and S4-S5 linker alpha helixes. In I384R, the S4-S5 linkers displayed a considerable lateral shift, particularly at the C-terminus end of the helix, 4.1 Å away from the WT structure (Fig. 3I). This movement was transduced to the N-terminus end of S4-S5 linker and the S4 helixes, dragging them 1.7 Å and 1.4 Å away from the open state structure, respectively. Since entry into the relaxed state has been associated with opening the pore ^14^, it is possible that the observed helical displacement in I384 formed the structural basis for the relaxed state of the VSDs.
Noncanonical conformation of the selectivity filter and the decreased volume of the closed state channel
One surprising observation of the I384R closed state structure comes from the selectivity filter conformation. Instead of the now classical linear coordination of 4 K^+^ ion densities seen in the conductive filter (Fig. 4B) ^18,49^ only 2 putative bound K^+^ ions, at the S2 and S4 sites of the SF were resolved (Fig. 4A). It is intriguing to see that the S3 K^+^ was absent in the structure, since generally, the S3 position typically displays the strongest K^+^ occupancy in K channel structural determinations ^50^. In our case, the coulombic density suggests that occupancy was similar for these two positions with a slightly higher occupancy at S4 position (Extended Data Fig. 6). Structurally, two small twists were observed at the S1 and S4 binding site when compared to the WT structure (Fig. 4C). The carbonyl group of the last glycine in the TVGYG selectivity filter, G446, flipped away from the pore, directly altering the binding site at S1 position (Fig. 4D). A similar twist was also observed at the bottom of the selectivity filter at the T442 position, which might account for the slight shift of the K^+^ ion at the S4 position compared to the WT.
Another intriguing observation in the closed state structure was the decrease of the protein volume in the transmembrane region. When comparing the WT with I384R structures, we noticed that the protein expanded laterally in the open state (Fig. 4E). This expansion was due to the translational movement of S4-S5 linker and the S6 helices. Area calculations with CHARMM_GUI show an asymmetric increase of the cross-section area of the open channel compared to the closed one (Fig. 4F) ^51^. The most significant expansion happened around the I470 and V474 region, where the cross section of the open channel increased by almost 10% (Extended Data Fig. 7). This expansion in volume might be the underlying mechanism of the reported mechanosensitivity of the Kv1 channels ^52,53^.
Discussion
Electromechanical coupling in voltage-gated ion channels and its energetics
Most voltage-gated channels share two basic functional modules: the voltage sensing domain and the pore domain. Electromechanical coupling describes the communication between these two modules. In the present study, we identified a tripartite pocket that we argue is essential for electromechanical coupling in the Shaker potassium channel, a strictly coupled channel. Structurally, interactions among Y485, F484, I384, E395 and R394 establish the functional connectivity between the pore and the S4-S5 linker. These intersubunit interactions likely contribute to the cooperativity of the voltage sensors and the pore opening as well ^54^. It has been demonstrated previously that reducing the side chain volume at positions Y485 and F484 in the S6 leads to shallower and right-shifted GV curves, ^14,55,56^, similar to what we show here in I384E, I384L and I384N. Mutagenesis experiments and thermodynamic cycle analysis among E395, R394 and Y485 have confirmed their energetic coupling and demonstrated their importance for electromechanical coupling ^36,37,57^. The identified tripartite pocket is thus likely responsible for the efficient transfer of the movement from the voltage sensor to the pore seen in the Kv channels.
In strictly coupled voltage-gated channels, the opening of the pore requires the activation of the voltage sensors. Understanding the energetics of this coupling is of fundamental importance to define the nature of EMC. It has long been debated whether it is energetically favorable for the pore to stay in the open or the closed state. In other words, are the voltage sensors doing work to “pull” the channel open or to “push” to keep the pore closed. While some computational work suggests the pore prefers to stay open in the absence of an external energy bias ^58^, our results argue otherwise. In the I384 mutants, all the uncoupled mutants (I384L, I384E and I384N) show a right shifted GV curve and a left shifted Q-V curve, an expected behavior if an energetic load to the sensor is decreased. In the partially uncoupled mutants, these results indicate that the pore is now less firmly coupled to the VSD, making it more difficult to open for a given charge movement, preferring to stay in the closed state.
In an uncoupled mutant, it is expected that the movement of the voltage sensor is independent of pore opening, therefore the left-shift of the QV curve observed in such mutant would reflect the energy required to open the pore. However, in the case of the uncoupled I384R mutant, the presence of four salt bridges between R384 and E395 introduces additional stabilization of the preopen state, a bias that is not expected to be present in the normal operation of the channel. While the newly formed interactions stabilize the closed state of the pore, they are likely to influence the voltage sensor movements as well, given the intimate connection between the voltage sensors and the S4-S5 linkers. The QV of I384R is shifted leftwards compared to the WT, indicating a lessened energy load, which was estimated to be at 3.41 ± 2.25 kcal/mol using the V_median_ approach ^59^. While it is tempting to conclude that this would be the energy for the pore opening, the presence of the aforementioned salt bridges invalidates this assumption. Most likely, the energy estimated has not only contributions of salt bridges, but also all the other energetic components present in the gating process.
Kv channel structure captured with a closed pore
Among the uncoupling mutants studied, I384R is the most extreme case, where only gating currents are seen. This is quite different from the classical W434F mutant, even though they both have minimal ionic conductance. W434F mutant speeds up the C-type inactivation and stabilized the channel in the inactivated state ^60^. Thus, I384R represented an ideal candidate to probe the conformation of the inner bundle gate, and we were able to capture the closed pore structure of Shaker potassium channel by disrupting the electromechanical coupling (Fig. 1, 2). It is worth pointing out that this uncoupled pore most likely represented the closed state, instead of an inactivated state, another nonconductive state in Kv channels. It is the EMC that was altered by substituting isoleucine at position 384, as indicated by the relative shifts in QV and GV curves (Fig. 1 & Extended Data Fig. 1). There are informative differences in the behavior of the gating currents measured in mutants W434F and I384R. In I384R, the gating current was significantly faster. This is consistent with severing the electromechanical coupling between PD and VSD, as the sensor is free to move in the absence of a mechanical “load”. In this case we are not tampering with the PD itself and therefore, the structure of the closed pore likely represents the true closed state of the pore in the WT channels. In contrast, gating currents in W434F, are a reflection of the VSD movement under a physiological load. The suppression of ionic currents is derived from effects downstream to the activation gating.
A surprising finding from the closed pore structure is the large degree of conformational changes happening along the entire length of the S6 helices. In contrast to the simple hinge-like movement seen in prokaryotes ^30,61^. Inner gate opening in eukaryotic strictly coupled Kv channels appears to be more complex, with both, a lateral movement of the PVP motif plus an additional rotational movement at the inner cavity of pore. This type of movement leads to the collapse of the inner cavity in the closed state, where hydrophobic I470 rotates and points directly into the permeation pathway. Our fluorescence experiments with ANAP directly demonstrated that a conformational change happens near position 470 as the channels open and close, supporting the structural observation. This is also consistent with early pharmacological studies with tetraethyl ammonium (TEA) ion. TEA exerts its blocking effects by entering the inner cavity of the channel in the open state and needs to be expelled out the channel before the pore closure can happen, a foot in the door effect ^62^. Mutating I470 to a smaller residue like cysteine, allows TEA to stay in the inner cavity in the closed state ^63^.
Conformation of selectivity filter in closed and open state
The selectivity filter in the I384R closed state structure was captured in what appears to be a noncanonical conformation. Current SF conformation is different from the typical conductive filter where four bound K^+^ ions orderly occupy the S1-S4 positions. It is also dissimilar to alternative “dilated” inactivated filter where the dilation of the top part of the filter abolish S1 and S2 binding sites (Extended Data Fig. 8A) ^5^ or the “pinched” inactivated filtered captured in prokaryotic Kv channel (Extended Data Fig. 8B). In I384R, two bound K^+^ ions were found at S2 and S4 position in the absence of any expansion at Y445 position or pinching at G444 ^6^ (Extended Data Fig. 8). It is then possible that as the channel opens, a conformational change around the SF occurs. Fluorescence measurements seem to be in line with this hypothesis; TMR labeled channels at T449 position, around the selectivity filter region, gave rise to a fluorescence change when the channels opened and the FV curve followed very closed the GV curve ^20^. In Nav channel, similar phenomenon has been suggested in the fast inactivated state as well ^64^. It is possible that the S6 movements during activation are allosterically communicated to the selectivity filter, triggering the transition from the noncanonical conformation to the typical conductive conformation ^65^. However, further investigation is certainly required before a definitive conclusion can be reached.
Isoleucine 470 as a shared structural element for both activation and inactivation
Pore radius calculation identified I470 in the inner cavity as the upper gate in closed state structure. This is a very interesting observation because I470 has been mostly associated with the C-type inactivation of Kv channels, previously. Physiological experiments mutating I470, together with T449, demonstrated that the inactivated state was rendered conductive by the double mutations ^66^, and computationally, long-time scale simulation also observed a coincidence of I470 movement and closure of the permeation pathway in the inactivated channels ^67^. Moreover, structural results from Kv4.2 identified the role of the equivalent isoleucine in inactivation mechanisms ^68^. In the case of Shaker K^+^ channel, the inactivation state is entered after the activation of the channel and the open inactivated state is the most dominant inactivated state in the channel ^69–71^. It appears that I470 is a shared structural element for both activation and inactivation and could serve as the link for the interplay of these two functional states.
Mechanosensitivity of Kv channel gating
In the closed state structure, we saw a collapsed inner cavity in the center of the protein which led to a global shrinkage of the protein volume. Compared to the open state structure, the protein occupied less area on the membrane in the closed state. It is worth noting that the open state structure was solved in lipid nanodisc and the current closed state structure was solved in detergents. The difference in structural determination method may result in an underestimation of the actual change in protein volume between closed and open state since it has been reported that reconstitution of proteins into nanodisc could introduce additional lateral forces from the lipid bilayer to the proteins ^72^. Therefore, it is possible that the actual cross section area of the channel in the closed state would be smaller and the actual expansion from closed to open would be more significant in a physiological condition.
One prediction coming from the observed decrease of the protein volume is that to open the channel, additional energy is required to push against the lateral pressure from the membrane and the amount of the lateral pressure will modulate the gating of the channel, giving rise to some mechanosensitivity to the channel. Some physiological experiments support this prediction. Excised patch clamp experiments have demonstrated that the GV curve of Shaker-IR-WT channels shifted to the left, opening more easily, with increased suction in the pipette ^52,73,74^. Additionally, mechanical modulation on the gating of the channel seems to be mostly limited to the change in the V_1/2_, which mostly reflects changes in steady-state energy level, without altering the number of charges that move. It is possible that the transition from the closed to the open state requires a global expansion of the protein which could serve as the molecular basis of the mechanosensitivity in strictly coupled Kv channels and possible other members of voltage gated channels, such as Nav channels ^75^.
Activation mechanism for strictly gated Kv channels
The present experimental results argue for a new gating model in strictly gated Kv channels (Fig. 5). At very hyperpolarizing voltage, the channels occupy the deepest closed state, where all four VSDs are in the down state. In this deep closed state, the permeation pathway is gated by I470 in the inner cavity as well as V474 at the bundle crossing region. Upon depolarization, asynchronous activation of the VSDs moves the channels into an ensemble of intermediate closed states, where some but not all VSDs are activated. The movement of the S4 in the VSDs creates a direct pull on the S4-S5 linker. However, this pull does not provide enough energy to open the channel and the VSDs are stabilized in the non-relaxed state. When all four voltage sensors move up, the channels enter a transient pre-open state, which is represented by the structure of the uncoupled channel. In this pre-open state, all the VSDs populate the activated (up) conformation which provides enough energy into the linkers to drive the pore into its open conformation. As part of a last concerted movement. S4-S5 linkers expand, allowing the opening of the pore and the relaxation of the VSDs. During opening, the S6 undergoes a roll and turn movement that rotates away the upper and lower activation gate, creating a water-filled permeation pathway. This movement is allosterically communicated to the selectivity filter leading to a small local conformation change, allowing for the selective permeation of the K^+^ ions and predisposing itself for C-type inactivation. This allosteric communication forms the structural and molecular basis for interaction among different functional states.
Methods
Site-directed mutagenesis and cRNA in-vitro synthesis
Shaker zH4 K + channel with fast inactivation removed (Δ6–46 - Shaker-IR ^76^)in pBSTA vector, flanked by β-globin sequences, was used in this study for all the physiological experiments. Point mutations were generated using mismatched mutagenic primers. The PCR product was digested in DpnI to remove the template and was used to transform the XL-gold ultra-competent cells. After ampicillin resistance screening, plasmids were purified from the colonies using standard miniprep protocols. Purified plasmids were sent for either Sanger (Genomics Facility at University of Chicago) or nanopore (Plasmidsaurus) sequencing to confirm the introduction of the point mutation and the absence of off-target mutations. cRNA was then transcribed in vitro from the linearized plasmids (T7 RNA expression kit; Ambion Invitrogen, Thermo Fisher Scientific, Waltham, MA).
Xenopus laevis oocyte preparation and channel expression
Ovaries of Xenopus laevis were purchased from XENOPUS1 (Dexter, Michigan). The follicular membrane was removed using collagenase type II (Worthington Biochemical Corporation) 2 mg/mL with bovine serum albumin at 1mg/mL (BSA). After defolliculation, stage V–VI oocytes were then selected and microinjected with 5-100 ng cRNA. Injected oocytes were incubated at 18°C for 1–5 days in SOS solution (in mM: 100 NaCl, 5 KCl, 2 CaCl_2_, 0.1 EDTA, and 10 HEPES at pH 7.4) supplemented with 50 μg/mL. Unless otherwise stated, all chemicals were purchased from Sigma-Aldrich.
Cut-open voltage clamp on Xenopus laevis oocytes
Macroscopic ionic and gating current were recorded using cut-open voltage clamp technique ^77^. Micropipettes filled with 3M CsCl or NaCl, with resistance between 0.4 and 0.8 MΩ were used to measure the internal voltage of the oocytes. Current data were online filtered at 20 kHz with a low-pass 4 pole Bessel filter and sampled by a 16-bit A/D (USB-1604; Measurement Computing, Norton, MA) converter at 1 MHz. All experiments were conducted at room temperature (~17 °C). For ionic current experiments, unless otherwise stated, were conducted in external solution consisted of in mM: 12 K methylsulfonate (MES), 108 N-methyl-D-glucamine (NMG) MES, 2 Ca MES, 10 HEPES, 0.1 EDTA, pH = 7.4 and internal solution consisted of in mM: 120 K MES, 10 HEPES, 2 EGTA, pH = 7.4. The capacitive transient was manually compensated with a dedicated circuit and in some cases, further removed by an online P/−4 protocol with a holding voltage of either − 80 or −90 mV ^78^. For gating current experiments, all experiments were conducted in external solutions consisted of in mM: 120 NMG MES, 2 Ca MES, 10 HEPES, 0.1 EDTA, pH = 7.4 and in internal solution consisted of in mM: 120 NMG MES, 2 Ca MES, 10 HEPES, 2 EGTA, pH = 7.4. The gating current was recorded with W434F background with the exception of I384R, which by itself produced no discernable ionic current.
Single channel recordings and noise analysis
Single channel recordings and noise analysis were performed on excised inside-out Xenopus Laevis oocyte patches. Briefly, the cells were injected with 5ng of RNA and maintained at 12 C° the day before the experiment. To remove the vitelline membrane, oocytes were incubated for 5 minutes in a hypertonic solution (SOS solution supplemented with 300 mM sucrose). This procedure shrank the oocyte separating the plasma membrane from the vitelline membrane, making the mechanical removal of the vitelline membrane easier without compromising the integrity of the cell. After removal of the vitelline membrane, oocytes were immediately and gently washed three times with intracellular solution containing in mM: 120KMES, 2EGTA, 10HEPES, pH = 7.40. They were placed in a recording chamber onto an inverted microscope. Current was recorded with an Axopatch 200B patch-clamp amplifier. The pipette solution consisted of, in mM, 120KMES, 2KCl, 2CaMES, 10HEPES, 0.1 EDTA, pH = 7.40. The resistances of the tips were between 13 ~ 17 MΩ for single channel experiments and 6 ~ 10 MΩ for noise analysis experiment. To reduce stray capacitance, the tips of the pipette were covered by Sylgard 184 (Dow Corning Corporation). Current was filtered with a digital 8 pole Bessel filter set at 10kHz (3384 Krohn-Hlite).
For noise analysis, we applied hundreds of depolarizing voltage pulses. The ionic currents elicited were averaged to obtain the mean. The variance and the mean were obtained using our Analysis software. For single channels, we recorded several current traces from voltage pulses to + 140mV. The histograms were obtained using Analysis.
Unnatural amino acid incorporation and voltage-clamp fluorimetry
To incorporate ANAP, we utilized the amber suppression technique. Briefly, the oocytes were injected with a mixture of ANAP-synthetase, ANAP methylester, ANAP-tRNA, Xenopus release factor 1 with D55E mutation and messenger RNA encoding the channel with the amber stop codon introduced at I470 position ^32,33,79^. The oocytes were incubated in the dark for 3 to 5 days prior to recording. The voltage clamp fluorometry setup and the filter set used in the study was similar to what was previously described ^21,34^. However, instead of using a photo diode, a photomultiplier was used to maximize the sensitivity of the signal, while allowing a decrease of the excitation light, decreasing photobleaching. The fluorophore was excited with a LED at 365nm (Thor lab M365L3). TMR labeling was done similarly as previous described ^20^. Briefly, oocytes were incubated in 1mM DTT for 15 minutes. Three washes were administered before transferring the cells to the labeling solution with 20 μM Tetramethylrhodamine-5-Maleimide (Thermo Fisher T6027) in a depolarizing solution (in Mm: 120 KMES, 2 CaMes, 10HEPES, 0.1EDTA, pH = 7.40) for 30 minutes.
Data Analysis
Ionic current was taken by the steady-state current level and converted to conductance using the following relationship:
where, I is the ionic current in steady state, V is the membrane voltage, and V_rev_ is the reversal potential for the conducting ion. The GV curves were then fitted using a two-state model given by the equation:
where z is the apparent charge expressed in units of elementary charge (e0), V is the voltage and V_1/2_ is the voltage of half-maximal conductance. R is the ideal gas constant; T is the temperature in Kelvin and F is the Faraday constant.
The gating charge was obtained by integrating the on and off-gating currents. They were plotted against the voltage to obtain the QV curves.
For the analysis of the normalized QV curves, we used a two-state model fitting equivalent to the one in Eq. 2 to fit the gating from the I384E, I384L, I384N, and I384R mutants. The equation was the following:
For WT, I384A, I384C, we used a three-state model fitting given by the following equation:
where N, z_1_, z_2_, V_1_ and V_2_ are the number of channels, the charges associated and equilibrium voltages for the first and second transition, respectively.
For nonstationary noise analysis, the mean variance data were fitted using a parabolic equation as follows:
where σ ^2^ is the variance, i is the single-channel current, < I > is the mean current, and N is the number of channels in the patch. We can estimate the maximal open probability (Po_max_) by knowing the maximum mean current (I_max_) using the following equation:
For single channel analysis data, we obtained all-points histogram binned at 0.05 pA using our Analysis software. The data was then fitted using a mixture of k Gaussian Distributions as follows ^80^:
Where a_i_ and f_i_ (y), are, respectively, the relative areas of the components and the Gaussian equation described by:
And
Where μ i and σ i are the mean and the standard deviation of the individual component i.
Data is presented as Mean ± SEM. For noise analysis, ionic and gating currents experiments we used at least 3 oocytes from different batches. For single channels, we recorded several oocytes and only for presentation purposes show the representative traces from one cell.
Expression and purification of mutant shaker potassium channel
Shaker-IR-I384R was subcloned into a modified pEG BacMam vector containing a C-terminal HRV 3C protease site, an eGFP tag and an 8×-His tag utilizing the 5’ NotI and 3’ XbaI restriction sites. The Bacmid plasmid was generated using the Bac-to-Bac system which was then used to transfect the Sf9 insect cells with Cellfectin (Thermo Fisher). After 4 to 7 days of incubation at 27°C, P_0_ virus was then collected after removing the remaining cell and debris. P_0_ was then amplified to produce P_1_ and P_2_ virus. 2L of HEK293S GnTI^−^ cells were infected with 200mL of P_2_ virus. After 24 h of shaking incubation at 37°C, sodium butyrate was added to the cells at a final concentration of 10mM and the culture was transferred to 30°C. Cells were collected 48 h after infection. The cell pellets were then washed with phosphate-buffered saline (PBS) at pH 7.4, collected by centrifugation, flash-frozen and stored at − 80°C for later purification.
All purification steps were conducted at 4°C. Frozen cell pellets were thawed in water bath and homogenized with a Dounce homogenizer in suspension buffer (150mM KCl, 2mM TCEP, 1mM EDTA, 50mM TRIS, Pierce protease inhibitor tablet (Thermo Fisher Scientific) PH 7.5). Resuspended cells culture were supplemented with DDM:CHS (10:1) at 1% final concentration (m. / v.) and were extracted for 2 hours with gentle agitation. The cell debris was pelleted by ultracentrifugation for 1 hr at 40000 rpm in Ti45 rotor (Beckman Coulter). The supernatant was collected and incubated with 2 ml CNBR-activated Sepharose beads (GE Healthcare) coupled with 4 mg high-affinity GFP nanobodies purified in-house for 3 hours. The Sepharose beads were then rinsed three times 10 column volume with the suspension buffer supplemented with 0.1% DDM:CHS (10:1), 0.05% DDM:CHS + 0.02% GDN, 0.02% GDN respectively. Then beads were incubated with HRV 3C protease overnight to release the purified channels. The next day, 3 column volume of suspension buffer was used to elute the beads. Eluted solution was concentrated with Amicon Ultra Centrifugal Filter unit (Millipore) with 100kDa cutoff to approximately 1mL and loaded onto a Superose6 (10 × 300 mm) gel filtration column (GE Healthcare) and separated with suspension buffer supplemented with 0.02% GDN.
Cryo-EM sample preparation and data acquisition
Samples purified from size-exclusion chromatography were pooled and concentrated to around 0.7 mg/mL measured with a nanodrop machine. 3.5 μL concentrated protein was applied to glow-discharged (30 s, 20 W Solarus Plasma Cleaner) Quantifoil grids (R 1.2/1.3 Au 300 mesh). The grids were blotted for 3s with blot-force 3 in a FEI Vitrobot Mark IV (Thermo Fisher) chamber with 100% humidity at room temperature before plunge froze in liquid ethane. Clipped grids were subsequently loaded onto a Titan Krios microscope. Single-particle movies were acquired using a K3 direct electron detector in super-resolution mode, coupled with a 20 eV GIF energy filter. Data collection was performed at a nominal magnification of ×81,000, corresponding to a pixel size of 0.5315 Å, binned by 2 during acquisition. The total electron dose was calibrated to 60 e−/Å^2^, distributed across 50 frames per movie.
Single-particle cryo-EM analysis
All steps for structure determination were performed using CryoSPARC ^81^, including motion correction and contrast transfer function (CTF) estimation. A subset of 2,000 particles was initially picked and classified in 2D to generate templates for template-based particle picking. Approximately 4,500,000 initial particles were picked and subjected to multiple rounds of 2D classification. From these, 180,000 particles were selected to generate three ab initio models with C1 symmetry. Particles from the best class (~ 116,000) were then processed for 3D refinement with C4 symmetry, yielding a nominal resolution of 5.7 Å. We performed 3D refinement using non-uniform refinement algorithm ^82^, which consistently yields best result in our data. A subsequent 3D classification was performed using the best-refined model from the previous round and a junk model generated ab initio. The best class from this step was further refined using C4 symmetry, resulting in a 3.55 Å nominal resolution. After identifying a subset of ݎ 80,000 particles for final refinement, Reference-Based Motion Correction was applied to estimate per-particle movement trajectories and empirical dose weights. A final 3D refinement step was performed, followed by postprocessing using a tighter mask and C4 symmetry enforcement. Local resolution was calculated using CryoSPARC. Validation was done using MOLprobity ^83^.
Model building and structural refinement
The Shaker-IR model (PDB 7SIP) was used as a template to build atomic models for the Shaker-IR-I384R mutants into our density map. All structural models were constructed using unsharpened maps. Initial models were generated through iterative rounds of manual model building in COOT ^84^ and real-space refinement in Phenix ^85^. Final refined atomic models were obtained using interactive flexible fitting in ISOLDE ^86^. All structural analyses and figure generation were performed using UCSF ChimeraX ^87,88^.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Jan L. Y. & Jan Y. N. Voltage-gated potassium channels and the diversity of electrical signalling. J. Physiol. 590, 2591–2599 (2012).22431339 10.1113/jphysiol.2011.224212 PMC 3424718 · doi ↗ · pubmed ↗
- 2Doyle D. A. The structure of the potassium channel: molecular basis of K + conduction and selectivity. Science 280, 69–77 (1998).9525859 10.1126/science.280.5360.69 · doi ↗ · pubmed ↗
- 3Tombola F., Pathak M. M. & Isacoff E. Y. How Does Voltage Open an Ion Channel? Annu. Rev. Cell Dev. Biol. 22, 23–52 (2006).16704338 10.1146/annurev.cellbio.21.020404.145837 · doi ↗ · pubmed ↗
- 4Holmgren M., Shin K. S. & Yellen G. The activation gate of a voltage-gated K + channel can be trapped in the open state by an intersubunit metal bridge. Neuron 21, 617–621 (1998).9768847 10.1016/s 0896-6273(00)80571-1 · doi ↗ · pubmed ↗
- 5Stix R. Eukaryotic Kv channel Shaker inactivates through selectivity filter dilation rather than collapse. Sci. Adv. 9, (2023).10.1126/sciadv.adj 5539 PMC 1070819638064553 · doi ↗ · pubmed ↗
- 6Cuello L. G., Jogini V., Cortes D. M. & Perozo E. Structural mechanism of C-type inactivation in K(+) channels. Nature 466, 203–208 (2010).20613835 10.1038/nature 09153 PMC 3033749 · doi ↗ · pubmed ↗
- 7Islas L. D. & Sigworth F. J. Voltage Sensitivity and Gating Charge in Shaker and Shab Family Potassium Channels. J. Gen. Physiol. 114, 723–742 (1999).10539976 10.1085/jgp.114.5.723PMC 2230542 · doi ↗ · pubmed ↗
- 8Hirschberg B., Rovner A., Lieberman M. & Patlak J. Transfer of twelve charges is needed to open skeletal muscle Na + channels. J. Gen. Physiol. 106, 1053–1068 (1995).8786350 10.1085/jgp.106.6.1053 PMC 2229305 · doi ↗ · pubmed ↗