Genomic characterization and phylogenetic analysis of chloroplast genome of Eurya nitida (Eurya Thunberg)
Yuyu Han, Dongzhen Jiang, Lei Zhou, Tao Jiang

TL;DR
This paper reports the complete chloroplast genome of Eurya nitida and its phylogenetic relationship with another Eurya species.
Contribution
The study provides the first complete chloroplast genome sequence of Eurya nitida and its phylogenetic analysis.
Findings
The chloroplast genome of E. nitida is 157,191 bp long with a typical quadripartite structure.
Phylogenetic analysis shows E. nitida is closely related to Eurya hebeclados.
Abstract
This study aimed to characterize the chloroplast genome of Eurya nitida Korthals. The complete chloroplast genome of Eurya nitida was sequenced and assembled using high-throughput sequencing technology. The genome spans 157,191 bp and exhibits a typical quadripartite structure, comprising a large single-copy region (LSC, 87,231 bp), a small single-copy region (SSC, 18,215 bp), and two inverted repeat regions (IRs, 51,744 bp each). Phylogenetic analysis revealed a close relationship between E. nitida and Eurya hebeclados Ling. This study elucidates the structural features and phylogenetic position of the E. nitida chloroplast genome.
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
 Figure 1
Figure 1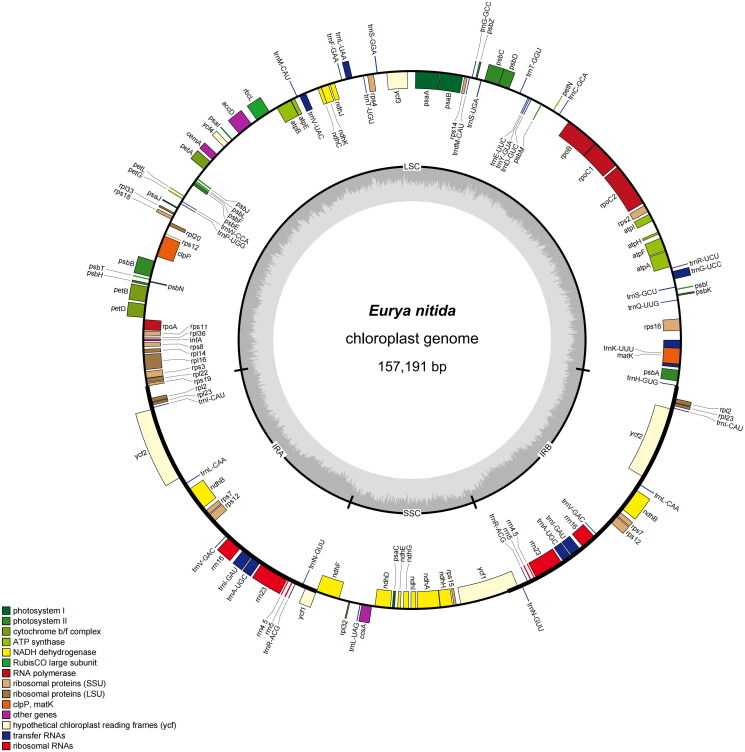 Figure 2
Figure 2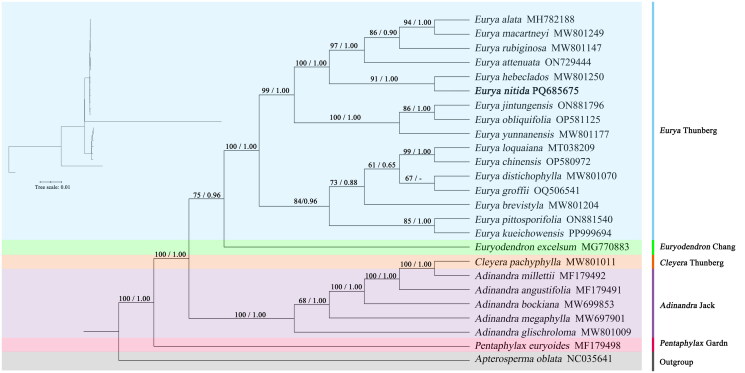 Figure 3
Figure 3- —GenBank
- —NCBI
- —Guizhou Provincial Basic Research Program (Natural Science)
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsIdentification and Quantification in Food · Genetic diversity and population structure · Genomics and Phylogenetic Studies
Introduction
Eurya nitida Korthals, first described by Korthals in 1840, belongs to the genus Eurya Thunberg. It’s taxonomic classification was historically ambiguous due to morphological similarities with related species (Haan and Korthals 1840). Advances in plant taxonomy during the 19th and 20th centuries clarified its distinction through morphological traits such as leaf shape and inflorescence structure (Ling 1951; Qiu and Zhong 1987). By the mid-twentieth century, E. nitida was definitively classified within the Pentaphylacaceae family, with boundaries delineated via morphological, ecological, and genetic comparisons against species like Eurya loquaiana Dunn and E. hebeclados Ling (Huang et al. 2004; Shi et al. 2008, 2009, 2012, 2014). Currently, E. nitida is recognized as a distinct species, primarily differentiated by shoot and leaf morphology. Chloroplast genomes have become pivotal in resolving taxonomic ambiguities and phylogenetic relationships, particularly in complex taxa (Ran et al. 2024; Xiao et al. 2024). However, the absence of chloroplast genomic data for E. nitida has hindered its comprehensive study. This research addresses this gap by characterizing the E. nitida chloroplast genome, thereby facilitating resource development, phylogenetic reconstruction, and evolutionary classification within Eurya Thunberg.
Materials and methods
Sample collection and DNA extraction
2.1.
Fresh leaves of E. nitida were collected from the Maolan National Nature Reserve (25.22015921 N, 107.92730734 E), Guizhou Province, China (Figure 1). Voucher specimens (Contact Person: Tao Jiang, [email protected], voucher number JT-20241111) were deposited at the Tree Specimen Laboratory, College of Forestry, Guizhou University. Chloroplast DNA was extracted using a modified CTAB protocol (Pahlich and Gerlitz 1980). DNA integrity was assessed via 1% agarose gel electrophoresis, while purity and concentration were measured using a NanoDrop spectrophotometer.
Reference image of Eurya nitida. (this photograph was taken by the authors of this article, Dongzhen Jiang and Lei Zhou. Leaves thinly leathery, elliptic, oblong-elliptic, margins densely serrate or finely obtusely toothed; fruit orbicular, blue-black at maturity.).
Sequencing and genome assembly and repeat sequence analysis
2.2.
Sequencing libraries were prepared through fragmentation, end repair, and adapter ligation. Libraries were sequenced on the Illumina NovaSeq 6000 platform. Raw reads were assembled de novo using GetOrganelle v.1.7.5.3 (Jin et al. 2020), followed by annotation with CPGAVAS2 (Shi et al. 2019). The complete chloroplast genome was deposited in NCBI (Accession: PQ685675) and visualized using OGDRAW (Lohse et al. 2007). In this study, simple sequence repeats (SSRs) within the E. nitida chloroplast genome were analyzed using MISA v.2.1 software (Beier et al. 2017). The detection parameters for repeat units were defined as follows: mononucleotides (≥10 repeats), dinucleotides (≥5 repeats), trinucleotides (≥4 repeats), tetranucleotides (≥3 repeats), pentanucleotides (≥3 repeats), and hexanucleotides (≥6 repeats). For larger repeat sequences, the REPuter online tool (Kurtz et al. 2001) was employed with a Hamming distance of 3 and a minimum repeat size of 30 bp. Four repeat types were identified: forward (F), reverse (R), complement (C), and palindromic (P).
Phylogenetic analysis
2.3.
Chloroplast genomes of 24 species (15 Eurya species and and the Apterosperma oblata (Accession: NC035641) was used as an outgroup) were retrieved from National Center for Biotechnology Information (NCBI). Sequences were aligned using MAFFT 7 (Katoh and Standley 2014), and MEGAX (Kumar et al. 2018) was used to manually correct and select the best maximum likelihood (ML) method to construct the phylogenetic tree model (GTR+I + G). The phylogenetic tree was constructed in IQ-TREE v.2.2.0, and the self-expansion support was set to 1000 (Trifinopoulos et al. 2016). The optimal model (HKY+G + I) was identified using MrModeltest v.2.3, and a Bayesian inference (BI) phylogenetic tree was subsequently reconstructed using MrBayes v.3.2.7 (Huelsenbeck and Ronquist 2001). Finally the phylogenetic tree was landscaped using the online tool iTOL V4 (Letunic and Bork 2021).
Results
Chloroplast genome structure
3.1.
The minimum and average read mapping depths were 367× and 1548.7×, respectively (Figure S1). Meanwhile, the structures of the trans-splicing and cis-splicing genes are shown in Figures S2 and S3. The E. nitida chloroplast genome (157,191 bp) comprises an LSC (87,231 bp), SSC (18,215 bp), and two IRs (51,744 bp each) (Figure 2). The GC content was 37.34% overall, varying regionally: LSC (35.32%), SSC (31.04%), and IRs (42.98%). The genome harbors 130 genes (85 protein-coding, 37 tRNA, 8 rRNA) (Table S1).
Chloroplast genome map of Eurya nitida. (Genes shown inside the circle are transcribed clockwise, those outside the circle are counterclockwise transcribed. The light grey and the darker grey in the inner circle represent AT and GC content, respectively. Different functional groups of genes are signed according to the colored boxes. LSC: large single copy; SSC: small single copy; IRA/IRB: Inverted repeat regions.).
Phylogenetic tree obtained via the maximum likelihood (ML) and bayesian inference (BI) methods of 25 species on the basis of complete cp genomes. (outgroup: Apterosperma oblata (NC035641); maximum likelihood (ML) and bayesian (BI) trees, BS ≥ 50% and PP ≥ 0.95 are indicated above branches as BS/PP) The following sequences, of which some already existed in NCBI database but were unpublished, were used: Eurya alata MH782188 (Zhang et al. 2021), Eurya brevistyla MW801204, Eurya chinensis OP580972, Eurya distichophylla MW801070, Eurya groffii OQ506541, Eurya hebeclados MW801250, Eurya jintungensis ON881796, Eurya kueichowensis PP999694, Eurya loquaiana MT038209 (Wang et al. 2020), Eurya macartneyi MW801249, Eurya obliquifolia OP581125, Eurya pittosporifolia ON881540, Eurya rubiginosa MW801147, Eurya attenuata ON729444 (Li et al. 2023), pentaphylax euryoides MF179498, Adinandra glischroloma MW801009, Adinandra bockiana MW699853 (Nguyen et al. 2023), Adinandra megaphylla MW697901, Adinandra millettii MF179492, Euryodendron excelsum MG770883 (Shi et al. 2019), Cleyera pachyphylla MW801011, Adinandra angustifolia MF179491, Apterosperma oblata NC035641.
Repeat sequence analysis
3.2.
A total of 209 SSRs were identified, predominantly mononucleotide repeats (127), followed by tri- (72), tetra- (6), di- (3), and pentanucleotide (1) repeats (Figure S4A). SSRs were enriched in the LSC (62.7%) and sparse in IRs (18.2%) (Figure S4B, C). Long repeats included forward (21), palindromic (27), and reverse (2) types (Figure S4D).
Phylogenetic analysis
3.3.
In this study, phylogenetic analysis (Figure 3) resolved E. nitida as a member of Eurya Thunberg, forming a clade with E. hebeclados (ML bootstrap = 91, BI posterior probability = 1.00). Sister branches included E. alata, E. macartneyi, E. rubiginosa, and E. attenuata.
Discussion
Chloroplast genomes are highly conserved in structure and maternally inherited, making them ideal for phylogenetic studies (Chen et al. 2010; Zou et al. 2021). The E. nitida genome aligns with reported Eurya chloroplast genomes (150–160 kb, quadripartite structure) and exhibits conserved GC content trends (Wang et al. 2020).
In this study, the whole genome sequence of E. nitida chloroplast was included in the NCBI database, which added new content to the Eurya Thunberg chloroplast genome data. Simple repeat sequences (SSRs) are commonly used in plant identification and genetic mapping because of their high content and polymorphism (Du et al. 2012). In this study, we found that the chloroplast genome of E. nitida contains 209 SSR sites, mainly single nucleotide repeats, with no C-type long repeats, and mainly polyA and polyT, which provides a basis for genetic analysis (Kuang et al. 2011). Chloroplast genome sequences can be used to infer phylogenetic relationships and determine species affinities (Xue et al. 2021). The present study supports that E. nitida is a member of Eurya Thunberg and obtains a high support rate; it also further finds its closest related species, E. hebeclados, and its sister branches. This is a crucial step in exploring the molecular phylogenetic relationships of E. nitida, which will help the study of molecular systematics and population evolution of Eurya Thunberg plants.
Conclusions
This study presents the first complete chloroplast genome of E. nitida, revealing its structural features, repeat content, and phylogenetic position. The data provide a foundation for future studies on Eurya taxonomy, evolution, and conservation.
Supplementary Material
Supplementary Figure.docx
Supplementary Table S1.xls
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Beier S, Thiel T, Münch T, Scholz U, Mascher M. 2017. MISA-web: a web server for microsatellite prediction. Bioinformatics. 33(16):2583–2585. doi:10.1093/bioinformatics/btx 198.28398459 PMC 5870701 · doi ↗ · pubmed ↗
- 2Chen SL, Yao H, Han JP, Liu C, Song JY, Shi LC, Zhu YG, Ma XY, Gao T, Pang XH, et al. 2010. Validation of the ITS 2 region as a novel DNA barcode for identifying medicinal plant species. P Lo S One. 5(1):e 8613. doi:10.1371/journal.pone.0008613.20062805 PMC 2799520 · doi ↗ · pubmed ↗
- 3Du QZ, Wang BW, Wei ZZ, Zhang ZZ, Zhang DQ, Li BL. 2012. Genetic diversity and population structure of Chinese white poplar (Populus tomentosa) revealed by SSR markers. J Hered. 103(6):853–862. doi:10.1093/jhered/ess 061.23008443 · doi ↗ · pubmed ↗
- 4Haan WD, Korthals PW. 1840. Verhandelingen over de natuurlijke geschiedenis der Nederlandsche Overzeesche Bezittingen. Leiden. In: commissie bij. S. en J. Luchtmans en C.C. van der Hoek.
- 5Huelsenbeck JP, Ronquist F. 2001. MRBAYES: Bayesian inference of phylogenetic trees. Bioinformatics. 17(8):754–755. doi:10.1093/bioinformatics/17.8.754.11524383 · doi ↗ · pubmed ↗
- 6Huang XT, Fang YL, Liu JQ. 2004. Comparative study on the leaf venation about Eurya in Fujian. J Fujian Normal Univ. 20(02):82–90.
- 7Jin JJ, Yu WB, Yang JB, Song Y, Depamphilis CW, Yi TS, Li DZ. 2020. Getorganelle: a fast and versatile toolkit for accurate de novo assembly of organelle genomes. Genome Biol. 21(1):241. doi:10.1186/s 13059-020-02154-5.32912315 PMC 7488116 · doi ↗ · pubmed ↗
- 8Kurtz S, Choudhuri JV, Ohlebusch E, Schleiermacher C, Stoye J, Giegerich R. 2001. RE Puter: the manifold applications of repeat analysis on a genomic scale. Nucleic Acids Res. 29(22):4633–4642. doi:10.1093/nar/29.22.4633.11713313 PMC 92531 · doi ↗ · pubmed ↗