The underappreciated, underrecognized problem of fourth chromosome trisomy in Drosophila melanogaster stocks and a simple, general method for building diplo-4 stocks from triplo-4 stocks
Kevin R. Cook

TL;DR
This paper addresses a common but overlooked issue in fruit fly genetics involving an extra fourth chromosome and offers a method to correct it.
Contribution
A new method is introduced to identify and correct fourth chromosome trisomy in Drosophila stocks using a haploinsufficient gene mutation.
Findings
Fourth chromosome trisomy is common in Drosophila stocks and can affect experimental results.
A mutation in a haploinsufficient gene can be used to identify trisomic stocks.
The method allows for the creation of disomic stocks from trisomic ones.
Abstract
Trisomy of the small, fourth chromosome is common in Drosophila melanogaster stocks. Even though the presence of an extra chromosome can confound the interpretation of experimental crosses, many Drosophila geneticists are unaware of this potential problem. Here I describe a simple method employing a mutation in a haploinsufficient gene to recognize fourth chromosome trisomy and establish disomic stocks from trisomic stocks.
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
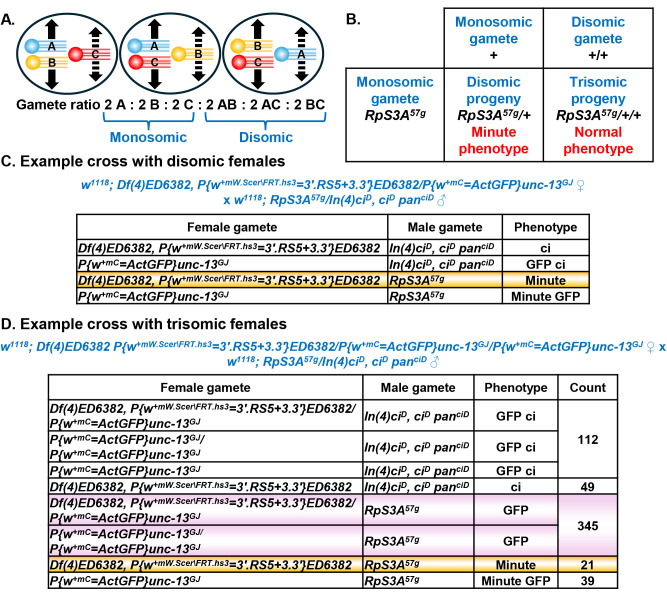 Figure 1
Figure 1|
Stock number |
Genotype |
Available from |
RRID |
|
647 |
|
Bloomington |
RRID:BDSC_647 |
|
1229 |
|
Bloomington |
RRID:BDSC_1229 |
|
9422 |
|
Bloomington |
RRID:BDSC_9422 |
|
9549 |
|
Bloomington |
RRID:BDSC_9549 |
|
90851 |
|
Bloomington |
RRID:BDSC_90851 |
|
90852 |
|
Bloomington |
RRID:BDSC_90852 |
|
150529 |
|
Kyoto |
RRID:DGGR_150529 |
|
150531 |
|
Kyoto |
RRID:DGGR_150531 |
|
150532 |
|
Kyoto |
RRID:DGGR_150532 |
|
602664 |
|
Bloomington |
RRID:BDSC_602664 |
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsGenomic variations and chromosomal abnormalities · Prenatal Screening and Diagnostics
Description
While diploid flies with an extra second or third chromosome invariably die early in development and those with an extra X chromosome have low viability, trisomic flies with an extra fourth chromosome survive at rates equivalent to normal, disomic flies and show nearly imperceptible phenotypic differences (Morgan et al. 1925). Flies trisomic for the fourth chromosome arise from nondisjunction, which occurs spontaneously in roughly 1 out of every 500 meioses in normal flies (Baker and Carpenter 1972)—meaning that trisomic flies appear frequently in stocks. Once a trisomic fly appears, it can produce trisomic progeny. ( Figure 1A shows that half the gametes of a trisomic fly are disomic if meiotic chromosome pairing and segregation are random.) If trisomy provides a selective advantage, trisomic flies can become a large proportion of the stock population. For example, it is likely that an extra normal chromosome can ameliorate deleterious effects associated with a deletion chromosome being heterozygous to a single normal chromosome. Indeed, in my recent work with the Fourth Chromosome Resource Project (Weasner et al. 2025), I found that several deletion stocks in the Bloomington and Kyoto stock centers were trisomic.
It is easy to see how using a stock with an unexpected, extra fourth chromosome can lead to confusing experimental results. Extra gene copies can cause higher-than-usual gene expression—with downstream consequences to gene regulation across the genome including modified phenotypes. Since most fourth chromosomes do not have dominant phenotypic markers, extra fourth chromosomes can be inadvertently carried along in crossing schemes and end up contaminating newly built stocks or making mutations appear to complement when they really do not.
Trisomy can be recognized from altered chromosome inheritance: each distinct fourth chromosome does not appear in half of the gametes as expected in disomic flies. Simplistically, one would expect that two of the three fourth chromosomes in a trisomic individual would pair at random and segregate from each other and that the segregation of these two chromosomes would not influence the segregation of the remaining chromosome ( Figure 1A ). In fact, such random, unbiased chromosome behavior, producing six gamete types in equal numbers, is rarely, if ever, seen. Sturtevant (1936) showed that, in general, two chromosomes preferentially pair and the remaining chromosome tends to segregate with one of them—leading to unequal numbers of the six gamete types. For example, I saw, in several dozen crosses with a trisomic * Df(4)ED6382/P{ActGFP}unc-13 ^GJ^ /P{ActGFP}unc-13 ^GJ^ * stock, that progeny from monosomic Df(4)ED6382 gametes were generally underrepresented.
To establish a disomic stock from a trisomic stock, one simply needs to identify a disomic fly with the chromosome of interest and mate it to disomic, balancer-bearing flies to recover siblings that can be mated to produce a stable stock. For example, I recovered the deletion chromosome from a trisomic * Df(4)ED6380/l(4)102EFf ^1^ /l(4)102EFf ^1^ * stock (which the DrosDel Project (Ryder et al. 2007) established using the unmarked lethal mutation * l(4)102EFf ^1^ * ) by crossing trisomic females to males homozygous for the recessive * ey ^1^ * mutation. Disomic * Df(4)ED6380/ey ^1^ * male progeny were easy to identify from their eyeless phenotypes and then to mate to * In(4)ci ^D^ , ci ^D^ pan ^ciD^ * females to recover * Df(4)ED6380/In(4)ci ^D^ , ci ^D^ pan ^ci^ * sibs for mating. Since noncomplementation is often not a realistic option for identifying disomic progeny carrying a chromosome of interest, fly geneticists have typically crossed flies from a trisomic stock to balancer-bearing flies and blindly set up multiple crosses with individual progeny in the hope of establishing at least one line with a disomic founder. For example, I attempted to establish disomic, deletion stocks by outcrossing * Df(4)ED6369/l(4)102EFf ^1^ /l(4)102EFf ^1^ * and * Df(4)ED6384/l(4)102EFf ^1^ /l(4)102EFf ^1^ * flies to disomic, balancer-bearing flies, backcrossing individual male progeny to balancer-bearing females and establishing sib matings. While this brute-force approach can undoubtedly be successful with enough effort, I found, after establishing roughly a dozen lines per deletion, that the * l(4)102EFf ^1^ * chromosome had been present in every founder male and that, consequently, every line was trisomic. Preferential segregation of the deletion chromosomes with the * l(4)102EFf ^1^ * chromosome in the original trisomic stocks (à la Sturtevant) likely reduced my chances of success.
After my frustrating experiences with those two trisomic stocks, I wanted a simpler solution to establishing disomic stocks from trisomic stocks, so I devised a general approach that employs a mutation in a haploinsufficient gene on the fourth chromosome, * RpS3A * (Morgan et al. 1926). * RpS3A * encodes a protein component of ribosomes and, when only a single functional gene copy is present, flies display Minute phenotypes (short and thin bristles, small body, reduced fertility and reduced viability) (Marygold et al. 2007). As shown in Figure 1B, the * RpS3A ^57g^ * loss-of-function mutation (Hochman et al. 1964) gives Minute phenotypes when it is combined with a single fourth chromosome but gives no mutant phenotypes when it is combined with two other fourth chromosomes. From comparing the example cross of disomic flies to * RpS3A ^57g^ * flies in Figure 1C to the example cross of trisomic flies to * RpS3A ^57g ^ * flies in Figure 1D, it is evident that parents from a trisomic stock will produce a class of non-Minute progeny (highlighted in pink in Figure 1D ) that is not present among the progeny of disomic parents. Furthermore, it is easy to select the appropriate class of disomic, Minute progeny (highlighted in yellow in both Figures 1C and D) to found a disomic stock. From the progeny of the cross shown in Figure 1D, I crossed the * w ^1118^ /Y; Df(4)ED6382/ RpS3A ^57g^ * males to * y ^1^ w ^^ ; TI{GMR-HMS04515}Gat ^eya^ /In(4)ci ^D^ , ci ^D^ pan ^ciD ^ * females (Nyberg et al. 2020) to establish a disomic * w ^^ * ; * Df(4)ED6382/In(4)ci ^D^ , ci ^D^ pan ^ciD^ * stock via sib mating. I have also used this approach to establish disomic stocks for Df(4)ED6369 and Df(4)ED6384 .
The * RpS3A ^57g^ * mutation was caused by a Doc transposable element insertion upstream of * RpS3A . * It drastically reduces * RpS3A * mRNA expression (van Beest et al. 1998; Kronhamn and Rasmuson-Lestander 1999). Even though * RpS3A ^57g^ * is likely a null allele, the * RpS3A ^57g^ * stock grows better than most stocks for multigene deletions removing * RpS3A * . * RpS3A ^57g^ * heterozygotes had lower viability than other progeny classes in Figure 1D, but I had little trouble using heterozygous males to establish new disomic stocks. The Minute bristle phenotypes are much easier to score than the other Minute phenotypes, and the bristle phenotypes are easier to score in females than males. I generally cross a single Minute male to multiple females to found a line and set up multiple lines in case I mistake non-Minute males for Minute males. A deletion for * RpS3A * could probably be substituted for * RpS3A ^57g^ * in isolating disomic progeny, but I would anticipate somewhat lower viability with deletion heterozygotes. In principle, females heterozygous for * RpS3A ^57g^ * can be used to establish disomic stocks, but the crosses would be less convenient, and the relatively low fecundity of these females would likely make the task more difficult.
The idea of distinguishing disomic flies from trisomic flies using a Minute mutation is not new. For example, Hawley et al. (1992) used a Minute mutation to screen for trisomic flies (though the authors did not report the identity of the specific Minute aberration or point mutation they used). After testing the use of * RpS3A ^57g^ * for distinguishing disomic and trisomic flies, I found that Bushey and Locke (2004) had probably used it to assure flies in some of their crosses were disomic, but they did not elaborate on the idea. Whether or not I reinvented the wheel, I hope this approach proves valuable to anyone undertaking crosses involving the fourth chromosome.
Methods
Fly cultures were reared under standard conditions. Genetic nomenclature reflects FlyBase release 2025_1 (Öztürk-Çolak et al. 2024). Information about the stocks listed in the Reagents section may be obtained via the websites for the Bloomington Drosophila Stock Center at Indiana University (https://bdsc.indiana.edu/) and the Department of Drosophila Genomics and Genetic Resources at the Kyoto Institute of Technology (https://www.dgrc.kit.ac.jp/).
Reagents
**: **
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Baker Bruce S Carpenter Adelaide T C 197261 GENETIC ANALYSIS OF SEX CHROMOSOMAL MEIOTIC MUTANTS IN DROSOPHILA MELANOGASTER Genetics 7121943-263125528610.1093/genetics/71.2.2554625747 PMC 1212782 · doi ↗ · pubmed ↗
- 2Bushey Daniel Locke John 2004111 Mutations in Su(var)205 and Su(var)3-7 Suppress P-Element-Dependent Silencing in Drosophila melanogaster Genetics 16831943-26311395141110.1534/genetics.104.02691415579693 PMC 1448784 · doi ↗ · pubmed ↗
- 3Hawley R. Scott Irick Holly Haddox Deana A. Whitley Michelle D. Arbel Tamar Jang Janet Mc Kim Kim Zitron Anne E. New Christine Childs Geoffrey Lohe Allan 199211 There are two mechanisms of achiasmate segregation in Drosophila females, one of which requires heterochromatic homology Developmental Genetics 1360192-253X 44046710.1002/dvg.10201306081304424 · doi ↗ · pubmed ↗
- 4Hochman B. Gloor H. Green M. M. 1964121 Analysis of chromosome 4 in Drosophila melanogaster. I. Spontaneous and X-ray-induced lethals Genetica 3510016-670710912610.1007/bf 0180487914170379 · doi ↗ · pubmed ↗
- 5Kronhamn Jesper Rasmuson-LestanderÅsa 1999121 Genetic organization of the ci-M-pan region on chromosome IV in Drosophila melanogaster Genome 4260831-27961144114910.1139/g 99-08510659782 · doi ↗ · pubmed ↗
- 6Marygold Steven J Roote John Reuter Gunter Lambertsson Andrew Ashburner Michael Millburn Gillian H Harrison Paul M Yu Zhan Kenmochi Naoya Kaufman Thomas C Leevers Sally J Cook Kevin R 20071010 The ribosomal protein genes and Minute loci of Drosophila melanogaster Genome Biology 8101474-760X 10.1186/gb-2007-8-10-r 216PMC 224629017927810 · doi ↗ · pubmed ↗
- 7Nyberg Kevin G. Nguyen Joseph Q. Kwon Yong-Jae Blythe Shelby Beitel Greg J. Carthew Richard 2020101 A pipeline for precise and efficient genome editing by sg RNA-Cas 9 RN Ps in Drosophila Fly 141-41933-6934344810.1080/19336934.2020.183241633016195 PMC 7746241 · doi ↗ · pubmed ↗
- 8Öztürk-Çolak Arzu Marygold Steven J Antonazzo Giulia Attrill Helen Goutte-Gattat Damien Jenkins Victoria K Matthews Beverley B Millburn Gillian dos Santos Gilberto Tabone Christopher J Fly Base Consortium Perrimon Norbert Gelbart Susan Russo Broll Kris Crosby Madeline dos Santos Gilberto Falls Kathleen Gramates L Sian Jenkins Victoria K Longden Ian Matthews Beverley B Seme Jolene Tabone Christopher J Zhou Pinglei Zytkovicz Mark Brown Nick Antonazzo Giulia Attrill Helen Goutte-Gattat Damien Larkin Aoife Marygold Steven Mc Lachl · doi ↗ · pubmed ↗