Genomic Modeling of an Outbreak of Multidrug-Resistant Shigella sonnei, California, USA, 2023–2024
Tyler Lloyd, Sana M. Khan, Dustin Heaton, Munira Shemsu, Vici Varghese, Jay Graham, Misha Gregory, Penny Dorfman, Megan Talton, Jessica DeVol, Nicola F. Müller, Kavita K. Trivedi

TL;DR
This paper describes a Shigella sonnei outbreak in California and shows how genomic sequencing can help track and manage bacterial disease outbreaks.
Contribution
The study demonstrates the practical use of phylodynamics in outbreak investigations by local health departments.
Findings
A Shigella sonnei outbreak was detected in the San Francisco Bay area in 2024.
Genomic sequencing and phylodynamics were used to trace the outbreak's spread and source.
Abstract
We report the detection of a Shigella sonnei outbreak from a small investigation in the San Francisco Bay area, California, USA, in 2024. By combining outbreak investigation with genomic sequencing, we show the utility of phylodynamics to aid outbreak investigations of bacterial pathogens by state or local public health departments.
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
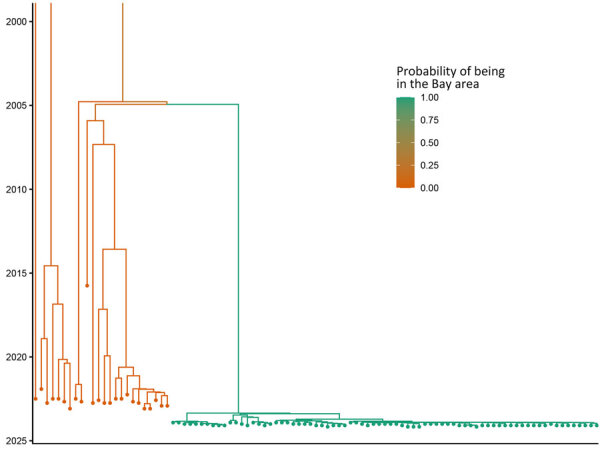 Figure 3
Figure 3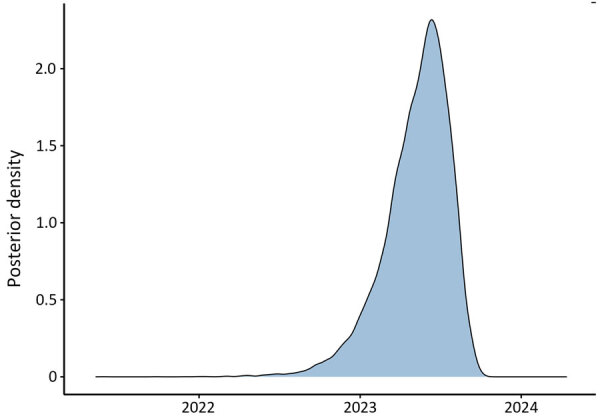 Figure 4
Figure 4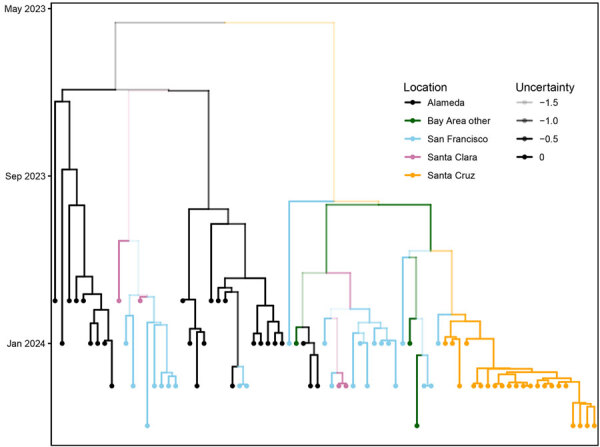 Figure 5
Figure 5Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsEscherichia coli research studies · Viral gastroenteritis research and epidemiology · Salmonella and Campylobacter epidemiology
In January 2024, a board and care facility (facility A) in the San Francisco Bay area, California, USA, reported 4 cases of Shigella sonnei infection to the Alameda County Public Health Department (ACPHD; San Leandro, California, USA). Shigellemia was confirmed in 3 patients. In February 2024, an independent living center (facility B) reported 3 cases of Shigella infection. Shigella bacteremia was confirmed in 2 patients (Figure 1,2). Shigella bacteremia (shigellemia) is rare but associated with immature immune responses or immunocompromised adults (1). We performed a 10-year retrospective review of Shigella cases in Alameda County and found 0.7% of cases had positive blood samples reported, consistent with other reviews on Shigella bacteremia (2). The 7 cases from 2 facilities prompted patient investigations at facilities A and B, and investigations into other S. sonnei patients in Alameda County during December 2023–February 2024.
Methods and Materials
Case investigations were limited but included symptom onset, severity, housing status, and other attainable risk factors. The outbreak investigation linked patients from facility B and 2 unhoused community members to a third location (facility C) where marginally housed community members gather. No clear transmission pattern was determined through epidemiologic investigation. We identified 19 genotypically identical S. sonnei isolates during December 2, 2023–February 26, 2024, among all cases in facilities A, B, and C.
Of the 19 patients, 13 (68%) were male and 6 (32%) female; median age was 59 years, and 9 (47%) were White and 10 (52%) non-Hispanic. Case investigations were completed on 16 of the 19 patients; 5 (26%) were experiencing homelessness, 4 (21%) were associated with facility A, 3 (15%) were associated with facility B, 4 (21%) had stable housing, and 3 (15%) had unknown housing. Drug use history was known in 3 patients. Of the 5 patients with shigellemia, 1 reported drug use. Sexual contact was unknown or denied during the incubation period for all patients. All treatment regimens where data were available were appropriate for the antimicrobial drug susceptibility data; all patients recovered.
Of the 3 patients associated with facility C, 2 were experiencing homelessness and 1 volunteered as a food handler at facility C while ill. The third patient from facility B visited facility C and had symptoms develop 14 days after exposure to the food handler at facility C. No other epidemiologic links were established among the 19 cases. No comorbidities were found in electronic medical records. However, determining precise risk factors in patients experiencing homelessness, such as where they sheltered during their infectious period, contact with each other, using the same resources, public restrooms or transportation, was not possible.
Results
Whole-genome sequencing (WGS) of Shigella isolates revealed highly similar sequences, suggesting an epidemiologic link. The time from notification of a potential outbreak in facility A to WGS confirmation was 8 days. We genotyped the isolates, which belonged to genotype 3.7.26, as previously described (3). This method removes repetitive regions with higher rates of potentially erroneous single-nucleotide polymorphisms (SNPs). The method simplified interpretation by providing a numerical genotype, making it easy to determine close ancestry and it was part of our routine bioinformatics workflow (4). References for genotype 3.7.26 are from the United Kingdom (2013) and France (2014). We confirmed phenotypic multidrug resistance by using antimicrobial drug resistance gene detection (Appendix).
During the retrospective sequencing of S. sonnei from patients treated in Alameda County, we identified patients with highly similar isolates in neighboring counties. The lack of specific links in the investigation and detection of cases from neighboring counties prompted ACPHD to notify the California Department of Public Health in March 2024, which led to a prioritization of Shigella isolates for sequencing. A total of 75 genetically related isolates were identified by California Department of Public Health by using PulseNet whole-genome multilocus sequence typing (MLST) (5), which showed relatedness but did not incorporate metadata. To reconstruct the spatial transmission dynamics of the outbreak, we performed a time-resolved, phylogeographic method known as the marginal approximation of the structured coalescent (MASCOT-skyline) in collaboration with the University of California, San Francisco. This approach uses Bayesian inference to reconstruct spatiotemporal transmission of pathogens and is implemented in the open-source program BEAST2 (6). MASCOT-skyline incorporates sampling time and sampling location of isolates. MASCOT-skyline then infers a posterior estimate of where the bacterial lineage was in the past. From this result, we inferred that all isolates were the result of a single introduction into the area (7; N.F. Müller, et al., unpub. data, https://pmc.ncbi.nlm.nih.gov/articles/PMC10942421). We obtained the molecular clock rate by contextualizing outbreak samples with 24 S. sonnei Pulsenet sequences from 2015–2023 to ensure an appropriate number of samples and the time span to effectively estimate the clock rate. The mean clock rate for the core-genome SNP alignment (length 1491 bp) was 3.341 × 10^−3^ substitutions per site per year. The sequences formed a distinct cluster from all other historical sequences (Figure 3). We inferred time to most recent common ancestor was most likely June 2023 (95% CI November 2022–August 2023), providing an upper bound on the time of introduction (Figure 4). We analyzed the outbreak at a more granular scale by using the same regional alignment and fixing clock rate while removing contextual sequences. We found the samples were geographically clustered by county within the outbreak (Figure 5).
Maximum clade credibility tree of outbreak samples and contextualized Shigella sonnei isolates from the United States, 2015–2023. The color scale denotes the probability of the node below each branch being in the San Francisco Bay, California, USA, area. Inferred by using MASCOT (6).
Distribution curve of the predicted dates of the most recent common ancestor of the Shigella sonnei outbreak isolates, California, USA, 2023–2024. The plot shows the posterior density for the common ancestor times of the Shigella sequences collected in the San Francisco Bay, California, USA, area. For a single introduction, the common ancestor time provides a lower bound on the timing of the introduction into the San Francisco Bay area.
Phylogenetic tree of Shigella sonnei outbreak isolates in the San Francisco Bay, California, USA, area with spatiotemporal metadata and tree uncertainty, 2023–2024. Branches are colored according to location. The opacity of the branches is equal to the uncertainty of the placement of each branch. Phylodynamic methods are incorporated into phylogenetic trees with time and location.
One patient was a food handler at facility C, but no evidence of foodborne transmission was found. The shigellemia cases prompted us to investigate this cluster; however, patients were immunocompetent, and virulence markers were identical to nonbloodstream infections. Host factors such as intravenous drug use and sexual contact were incomplete and remain possible factors for shigellemia.
Discussion
The advent of phylodynamic approaches and genomic epidemiology has provided public health with additional insight into the spread of diseases, transmission chains, and mutations when using laboratory data paired with epidemiologic information. In this article, we demonstrate the use of phylodynamic modeling alongside a traditional case investigation to better determine outbreak dynamics and inform public health actions. Bacterial genomic epidemiology has historically relied on MLST, SNPs, whole-genome MLST, or a combination of technologic tools. However, those tools do not enable us to characterize the direction and timing of disease spread. SNP cutoff levels have shown variable specificity and sensitivity in identifying closely related bacterial isolates (8).
We also describe the role of the local public health laboratory to initiate enhanced WGS of S. sonnei to discover unlinked cases and identify a regional outbreak. We describe the timeline of the outbreak identification, notification of the state public health department, and phylodynamic methods to provide evidence of a single introduction and incorporate metadata into bacterial genomic epidemiology. However, those models do not guarantee complete ascertainment of transmission, and the inability to gather complete data on risk factors to link specific case manifestations, symptoms, or other factors associated with shigellemia or its mode of transmission is a limitation of our study. Ideally, genomic sequencing paired with epidemiologic information gathered, such as case manifestation, risk factors identified, and symptoms, can provide improved insights into the drivers of transmission. This information can be particularly helpful when investigating outbreaks in communities such as persons experiencing homelessness, when epidemiologic information may be limited. We recommend public health prevention measures focus on the proper maintenance, routine disinfection, and cleaning of public restroom facilities and handwashing stations, particularly in places that are frequented by persons experiencing homelessness.
AppendixAdditional information about genomic modeling of an outbreak of multidrug resistant Shigella sonnei, California, USA, 2023–2024.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Rotramel HE, Zamir HS. Shigella bacteremia in an immunocompetent patient. Cureus. 2021;13:e 19778.34950556 10.7759/cureus.19778 PMC 8687798 · doi ↗ · pubmed ↗
- 2Stefanovic A, Matic N, Ritchie G, Lowe CF, Leung V, Hull M, et al. Multidrug-resistant Shigella sonnei bacteremia among persons experiencing homelessness, Vancouver, British Columbia, Canada. Emerg Infect Dis. 2023;29:1668–71. 10.3201/eid 2908.23032337486309 PMC 10370870 · doi ↗ · pubmed ↗
- 3Hawkey J, Paranagama K, Baker KS, Bengtsson RJ, Weill FX, Thomson NR, et al. Global population structure and genotyping framework for genomic surveillance of the major dysentery pathogen, Shigella sonnei. Nat Commun. 2021;12:2684. 10.1038/s 41467-021-22700-433976138 PMC 8113504 · doi ↗ · pubmed ↗
- 4Libuit KG, Doughty EL, Otieno JR, Ambrosio F, Kapsak CJ, Smith EA, et al. Accelerating bioinformatics implementation in public health. Microb Genom. 2023;9:mgen 001051. 10.1099/mgen.0.00105137428142 PMC 10438813 · doi ↗ · pubmed ↗
- 5Ribot EM, Freeman M, Hise KB, Gerner-Smidt P. Pulse Net: entering the age of next-generation sequencing. Foodborne Pathog Dis. 2019;16:451–6. 10.1089/fpd.2019.263431241352 PMC 6653803 · doi ↗ · pubmed ↗
- 6Bouckaert R, Vaughan TG, Barido-Sottani J, Duchêne S, Fourment M, Gavryushkina A, et al. BEAST 2.5: An advanced software platform for Bayesian evolutionary analysis. PLOS Comput Biol. 2019;15:e 1006650. 10.1371/journal.pcbi.100665030958812 PMC 6472827 · doi ↗ · pubmed ↗
- 7Müller NF, Rasmussen D, Stadler T. MASCOT: parameter and state inference under the marginal structured coalescent approximation. Bioinformatics. 2018;34:3843–8. 10.1093/bioinformatics/bty 40629790921 PMC 6223361 · doi ↗ · pubmed ↗
- 8Mitchell HD, Mikhail AFW, Painset A, Dallman TJ, Jenkins C, Thomson NR, et al. Use of whole-genome sequencing to identify clusters of Shigella flexneri associated with sexual transmission in men who have sex with men in England: a validation study using linked behavioural data. Microb Genom. 2019;5:e 000311. 10.1099/mgen.0.00031131682221 PMC 6927305 · doi ↗ · pubmed ↗