Recovering high-resolution information using energy filtering in MicroED
Max T. B. Clabbers, Tamir Gonen

TL;DR
Energy filtering in electron crystallography improves data quality by reducing noise and preserving high-resolution information.
Contribution
The study demonstrates that energy filtering can recover high-resolution data from electron diffraction, even after radiation damage.
Findings
Energy filtering reduces noise and sharpens Bragg peaks in electron diffraction data.
High-resolution information is preserved and intensity statistics improve with energy filtering.
Energy filtering enhances data quality and reliability in macromolecular electron crystallography.
Abstract
Inelastic scattering poses a significant challenge in electron crystallography by elevating background noise and broadening Bragg peaks, thereby reducing the overall signal-to-noise ratio. This is particularly detrimental to data quality in structural biology, as the diffraction signal is relatively weak. These effects are aggravated even further by the decay of the diffracted intensities as a result of accumulated radiation damage, and rapidly fading high-resolution information can disappear beneath the noise. Loss of high-resolution reflections can partly be mitigated using energy filtering, which removes inelastically scattered electrons and improves data quality and resolution. Here, we systematically compared unfiltered and energy-filtered microcrystal electron diffraction data from proteinase K crystals, first collecting an unfiltered dataset followed directly by a second sweep…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
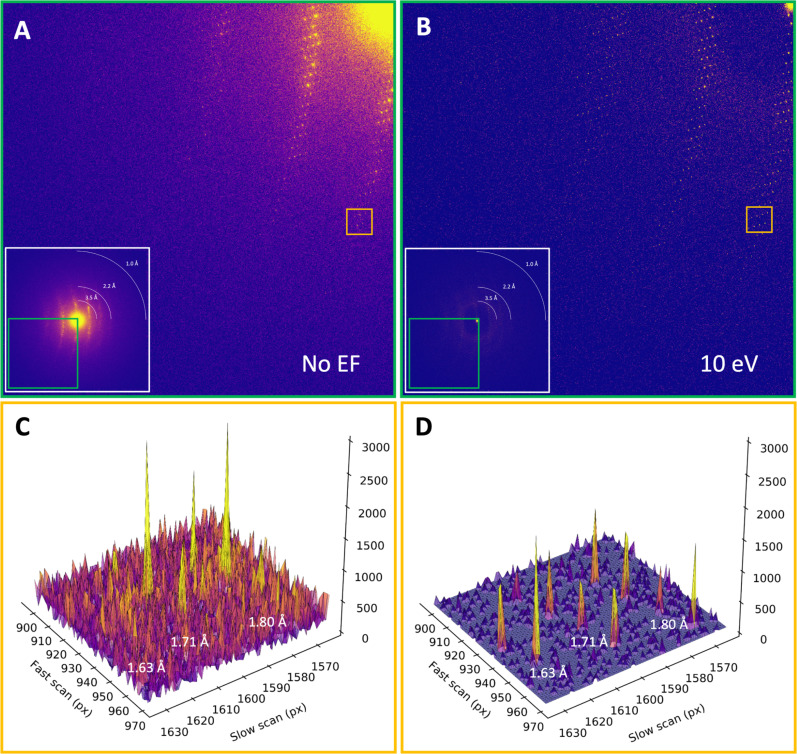 FIG. 1
FIG. 1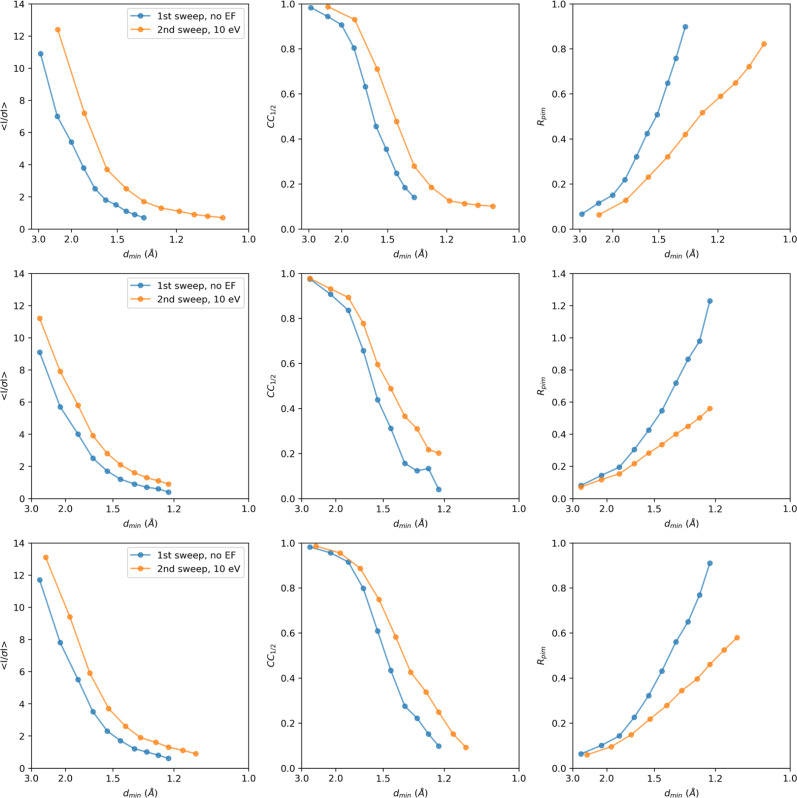 FIG. 2
FIG. 2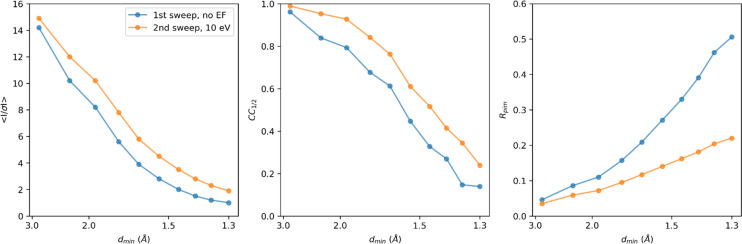 FIG. 3
FIG. 3| First pass, no EF | Second pass, 10 eV EF slit | |
|---|---|---|
| Data collection | ||
| No. of crystals | 11 | 11 |
| Flux density (e−/Å2/s) | 0.002 | 0.002 |
| Fluence (e−/Å2) | 0.84 | 0.84 |
| Total fluence (e−/Å2) | 0.84 | 1.68 |
| Dose (MGy) | 3.08 | 6.16 |
| Data integration | ||
| Space group | ||
| Unit cell dimensions | ||
| 67.62, 67,62, 106.59 | 67.64, 67.64, 106.67 | |
| 90, 90, 90 | 90, 90, 90 | |
| Resolution (Å) | 57.10–1.30 (1.33–1.30) | 43.64–1.30 (1.33–1.30) |
| Observed reflections | 1 377 854 (102 897) | 1 348 162 (102 276) |
| Unique reflections | 59 002 (4294) | 59 010 (4297) |
| Multiplicity | 23.4 (24.0) | 22.8 (23.8) |
| Completeness (%) | 96.1 (96.3) | 95.9 (96.0) |
| Rmerge | 0.495 (2.463) | 0.302 (1.084) |
| ⟨ | 5.2 (1.0) | 6.7 (1.9) |
| CC1/2 (%) | 95.9 (15.2) | 98.9 (23.9) |
| Refinement | ||
| Resolution (Å) | 57.10–1.30 | 43.64–1.30 |
| No. of reflections | 57 511 | 58 442 |
| Rwork/Rfree | 0.181/0.226 | 0.189/0.228 |
| factors (Å2) | ||
| Overall | 12.57 | 12.61 |
| Protein | 9.98 | 10.48 |
| Ligands/ions | 10.68 | 11.87 |
| Waters | 27.62 | 27.35 |
| R.m.s. deviations | ||
| Bond lengths (Å) | 0.011 | 0.009 |
| Bond angles (°) | 1.13 | 1.02 |
- —National Institute of General Medical Sciences 10.13039/100000057
- —Defense Threat Reduction Agency 10.13039/100000774
- —Howard Hughes Medical Institute 10.13039/100000011
- —Defense Threat Reduction Agency 10.13039/100000774
- —Yes, funding has been received from NIH 10.13039/100000002
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsEnzyme Structure and Function · Advanced Electron Microscopy Techniques and Applications · Protein Structure and Dynamics
INTRODUCTION
Inelastic scattering poses a significant challenge in electron crystallography, as inelastic events have a 3–4 times higher probability to occur compared to elastic scattering in hydrated protein crystals (Henderson, 1995; Latychevskaia and Abrahams, 2019). Inelastically scattered electrons lose coherence, causing elevated background levels and a broadening of the Bragg spots. This increase in noise can interfere with the detection of the diffraction spots and reduce accuracy of the intensity measurements. The effects are particularly detrimental to data quality from biological samples, which typically have a relatively weaker diffraction signal and experience substantial scattering contributions from the bulk solvent. This is further exacerbated during data collection, as the mean diffracted intensities rapidly fade owing to radiation damage (Hattne et al., 2018). As a result of the increasing dose, faint high-resolution reflections are lost when the diffraction signal at high scattering angles drops below the background noise. Loss of high-resolution information, due to dose-sensitivity and increased noise from inelastic scattering, compromises the accuracy of structural models and can obscure detailed features essential for biological interpretation. Therefore, mitigating these effects is crucial for improving data quality and achieving more precise structural insights.
Energy filtering removes inelastically scattered electrons above a defined energy-loss threshold, improving the signal-to-noise ratio and quality of the diffraction measurements in both materials science (Gemmi and Oleynikov, 2013; Yang et al., 2022) and structural biology (Gonen et al., 2004, 2005; Yonekura et al., 2002, 2015, 2019). Recently, we implemented a strategy for microcrystal electron diffraction (MicroED) data collection that combines electron counting (Martynowycz et al., 2022) with energy filtering (Clabbers et al., 2025a). Using this integrated approach, energy-filtered data from proteinase K crystals showed a significant improvement in overall data quality and high-resolution information extending to 1.09 Å, compared to 1.4 Å resolution for crystals recorded without energy filtering (Clabbers et al., 2025a). Further analysis of energy filter slit widths revealed narrower slit settings, reduced noise, and yielded sharper peaks, improving spot separation and intensity measurements with better internal consistency, ultimately leading to more accurate structural models (Clabbers et al., 2025b). However, direct comparisons of intensity and model statistics between filtered and unfiltered data between crystals remain challenging due to variations in crystallinity, isomorphism, and radiation damage between different crystals.
Here, we systematically compare unfiltered and energy-filtered MicroED data from the same crystal samples. Proteinase K crystals were machined using focused ion beam milling to lamellae with an optimal thickness of 300 nm (Martynowycz et al., 2021, 2023) and transferred to the transmission electron microscope (TEM) that was aligned with low flux density conditions of ∼0.002 e^−^/Å^2^ for data collection (Clabbers et al., 2025a). For each lamella, two consecutive data collection passes were performed. First, an unfiltered dataset was recorded with the energy filter slit retracted, covering a 20.0° continuous rotation sweep with a total fluence of 0.84 e^−^/Å^2^, corresponding to an absorbed dose of ∼3.1 MGy. At this fluence, the amount of dose evidence of radiation damage should be visible (Garman 2010, Hattne et al., 2018). Immediately after, a second pass was recorded using the same protocol, but with the energy filter slit inserted. This approach was repeated for 11 lamellae to ensure sufficiently high completeness and minimize variability for a comparative analysis. To enable a direct comparison, data were processed using XDS and truncated at a cross correlation between two random half sets (CC1/2) that was still significant at the 0.1% level in the highest resolution shell (Karplus and Diederichs, 2012).
Consistent with past studies (Clabbers et al., 2025a, 2025b), energy filtering visibly improved the MicroED data quality, with reduced background noise, sharper Bragg peaks, and high-resolution information extending out further compared to unfiltered data (Fig. 1). Interestingly, the filtered MicroED data consistently showed improved intensity statistics, and in all but two instances higher-resolution information, even though the absorbed dose for the second pass was effectively doubled compared to any of the unfiltered datasets (Figs. 2, S1, and S2). For example, a first crystal recorded without filtering was truncated at 1.34 Å resolution, with a CC1/2 of 19.8% and a mean I/σI of 0.89 in the highest resolution shell. In contrast, the second energy-filtered pass provided significantly higher information up to 1.06 Å resolution, with a CC1/2 of 10.2% and a mean I/σI of 0.71 (Fig. 2). In two cases where the resolution did not improve, intensity statistics of the filtered data were significantly better in all resolution shells compared to their unfiltered counterparts (Figs. 2 and S1). Variations in attainable resolution for different lamellae likely stem from differences in crystallinity, lamella quality, and lattice orientation.
Comparison of unfiltered and filtered MicroED data. (a) Magnified panel from an unfiltered MicroED diffraction frame, summed over the last 10 s of continuous rotation data collection with a flux density of 0.002 e−/Å2/s and an accumulated dose of ∼3.1 MGy. The inset shows the entire frame, with the magnified panel highlighted in green. (b) Magnified diffraction frame covering the same wedge in reciprocal space as shown in (a) for a second energy-filtered MicroED pass, with a flux density of 0.002 e−/Å2/s and a total accumulated dose of ∼6.2 MGy. The inset shows the entire frame, with the magnified panel highlighted in green. (c) Peak profiles shown for the area highlighted in orange in (a), showing rows of diffraction spots at ∼1.6, ∼1.7, and ∼1.8 Å resolution. (d) Peak profiles for filtered MicroED data for the same area as shown in (c) and highlighted orange in (b), showing rows of diffraction spots at ∼1.6, ∼1.7, and ∼1.8 Å resolution that have a much better separation of the Bragg peaks from the background noise compared to the unfiltered data in (c).
Intensity statistics for three MicroED data collection series. For each series, two datasets were collected from the same lamella, where the first pass did not use energy filtering (blue), and the second pass had the 10 eV energy filter slit inserted (orange). For each lamella, the crystallographic quality indicators mean I/σI, CC1/2, and Rpim are plotted as a function of the resolution. Individual datasets were truncated at CC1/2 that was still significant at the 0.1% level in the highest resolution shell.
The merged unfiltered data from the first data collection sweeps showed signification information up to 1.3 Å resolution (Table I). To enable a direct comparison of the intensity and model statistics, data recorded with energy filtering were truncated at the same resolution, even though the crystals diffracted to higher resolution in the second sweep (Table I and Fig. 3). At the same resolution, the energy-filtered data show better statistics such as the mean I/σI, CC1/2, and R-factors, across all resolution shells, despite having absorbed twice the dose compared to the unfiltered data (Fig. 3). Notably, the mean I/σI in the highest resolution shell was twice as high, and the CC1/2 was improved overall, indicating a better internal consistency between the intensity measurements of the filtered data (Table I). The quality of the resulting models was rather similar, showing a Rwork/Rfree of 0.189/0.228 for the filtered data compared to the unfiltered data at 0.181/0.226 (Table I). The fact that the model R-factors for the filtered data did not improve, even though the intensity statistics were significantly better, likely is the result of the higher accumulated dose and increased radiation damage compromising structural integrity. Visual inspection of the map did not reveal any major differences in model quality, and B-factors were rather similar (Table I).
Merging statistics for unfiltered and filtered MicroED data. The crystallographic quality indicators mean I/σI, CC1/2, and Rpim, are plotted as a function of the resolution. For a side-by-side comparison, both data were merged and truncated at 1.3 Å resolution.
Our results highlight the advantages of energy filtering in MicroED for macromolecular crystallography. By directly comparing filtered and unfiltered data recorded from two sweeps of the same proteinase K lamellae, we demonstrate that energy filtering recovers high-resolution information, even after the crystal has absorbed double the amount of radiation dose. These improvements translate into better intensity statistics and greater internal consistency. Importantly, our results further suggest that high-resolution reflections, which would otherwise have been lost due to dose-sensitivity, could be partially recovered using energy filtering, which reduces the noise. While model quality was similar in these experiments, previous comparisons between unfiltered and filtered data collected from separate but comparable crystals at the same fluence showed improve data quality with energy filtering (Clabbers et al., 2025a). Together, these findings underscore the potential of energy filtering to overcome key limitations in electron crystallography and, indeed, all cryoEM modalities, offering a robust approach that reduces noise and partly mitigates information loss, yielding higher-resolution structures with improved reliability and precision.
METHODS
Crystallization
Proteinase K microcrystals were are grown as previously described (Masuda et al., 2017; Clabbers et al., 2025a). Briefly, crystals were grown by mixing 40 mg/ml protein solution in 20 mM MES-NaOH pH 6.5 at a 1:1 ratio with a precipitant solution of 0.5 M NaNO_3_, 0.1 M CaCl_2_, and 0.1 M MES-NaOH pH 6.5. The mixture was incubated at 4 °C, and microcrystals with dimensions of approximately 7–12 μm appeared within 24 h.
Sample preparation
A standard holey carbon electron microscopy grid (Quantifoil, Cu 200 mesh, R2/2) was glow discharged for 30 s at 15 mA on the negative setting. The sample was prepared using a Leica GP2 vitrification device set at 4 °C and 90% humidity, where 3 μl of crystal solution was deposited onto the grid, incubated for tens, and any excess liquid was blotted away from the back side. The sample was then soaked on-grid for 30s with 3 μl cryoprotectant solution of 30% glycerol, 250 mM NaNO_3_, 50 mM CaCl_2_, and 60 mM MES-NaOH pH 6.5. Any excess solution was blotted away using filter paper, and the grid was rapidly vitrified using liquid ethane.
Focused ion beam milling
The grid was loaded onto a Helios Hydra 5 CX dual-beam plasma FIB/SEM (Thermo Fisher Scientific). Prior to milling, the grid was coated with a thin protective layer of platinum for 45s using the gas injection system. Microcrystals of proteinase K were machined using a 30 kV Argon plasma ion beam with a stepwise protocol as described previously (Martynowycz et al., 2023; Clabbers et al., 2024). Briefly, coarse milling steps were performed using a 2.0 nA current to a thickness of approximately 3 μm. Finer milling steps at 0.2 nA were used to thin the lamellae to 600 nm. Final polishing steps were performed at 60 pA down to an optimal thickness of 300 nm, equal to approximately one time the inelastic mean free at 300 kV (Martynowycz et al., 2021). In one session, 11 lamellae were prepared sequentially using this same protocol on the same grid. After milling, the grid was directly cryo-transferred to the transmission electron microscope (TEM) for data collection. During the transfer step, the grid was rotated by 90° relative to the milling direction such that the rotation axis on the microscope is perpendicular to the milling direction.
Data collection
Diffraction data were collected on a Titan Krios G3i TEM (Thermo Fisher Scientific) equipped with a X-FEG operated at an acceleration voltage of 300 kV, a post-column Selectris energy filter, and a Falcon 4i direct electron detector. The microscope was aligned for low flux density conditions using the 50 μm C2 aperture, spot size 11, gun lens setting 8, and a parallel electron beam of 10 μm diameter. Under these conditions, the flux density was approximately 0.002 e^−^/Å^2^/s. The energy spread of the emitted electrons was characterized as ΔE = 0.834 ± 0.006 eV at full-width half max (FWHM). The zero-loss peak of the energy filter was aligned in the defocused diffraction mode and centered within the SA aperture. Data were collected using the 150 μm selected area (SA) aperture, defining an area with ∼3.5 μm diameter at the sample plane. The detector distance was 1402 mm and calibrated prior to data collection using a standard evaporated aluminum grid (Ted Pella). For each lamella, MicroED data were collected using the continuous rotation method (CITE) over two consecutive passes. The first pass involved unfiltered data collection with the energy filter slit retracted, covering a 20.0° rotation range with an angular increment of 0.0476 °/s, an exposure time of 420 s, and a total fluence of 0.84 e^−^/Å^2^ (equal to an absorbed dose of ∼3.1 MGy). The second pass followed immediately after, using the same protocol but with the 10 eV energy filter slit inserted. The total absorbed dose after the second sweep was approximately 6.2 MGy. Equivalent dose values were calculated using the EMED subprogram of RADDOSE-3D (Dickerson et al., 2024). Data were recorded on a Falcon 4i direct electron detector in the electron counting mode, operating at an internal frame rate of ∼320 Hz. The proactive dose protector was manually disabled. Raw data were written in electron event representation (EER) format with an effective readout speed of ∼308 frames per second, not counting gap frames.
Data processing and refinement
Individual MicroED datasets in EER format were binned by two and converted to SMV format after applying post-counting gain corrections using the MicroED tools (available at https://cryoem.ucla.edu/downloads). Diffraction data were summed in batches of 308, such that each summed image represents a 1 s exposure. Individual MicroED datasets were processed using XDS (Kabsch, 2010). Data were integrated up to a cross correlation between two random half sets that was still significant at the 0.1% level (Karplus and Diederichs, 2012). Individual datasets were merged using XSCALE (Kabsch, 2010). The merged data were truncated at a 1.3 Å resolution, enabling a direct comparison between intensity and model statistics. Data were merged using Aimless (Evans and Murshudov, 2013) and refined against the same proteinase K model (PDB ID 9DHO), which we reported previously at 1.09 Å resolution from energy-filtered MicroED data (Clabbers et al., 2025a). Both structures were refined using the same protocol in phenix.refine (Afonine et al., 2012), including electron scattering factors, automated optimization of the geometry and ADP weights, and individual anisotropic B-factors for all non-hydrogen atoms (see the supplementary material).
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Afonine, P. V., Grosse-Kunstleve, R. W., Echols, N., Headd, J. J., Moriarty, N. W., Mustyakimov, M., Terwilliger, T. C., Urzhumtsev, A., Zwart, P. H., and Adams, P. D., Acta Crystallogr. D 68, 352–367 (2012).10.1107/S 090744491200130822505256 PMC 3322595 · doi ↗ · pubmed ↗
- 2Clabbers, M. T. B., Hattne, J., Martynowycz, M. W., and Gonen, T., Nat. Commun. 16, 2247 (2025 a).10.1038/s 41467-025-57425-140050283 PMC 11885823 · doi ↗ · pubmed ↗
- 3Clabbers, M. T. B., Hattne, J., Martynowycz, M. W., and Gonen, T., “Characterization of energy filtering slit widths for Micro ED data collection,” bio Rxiv (2025 b).10.1101/2025.02.24.639939 · doi ↗
- 4Dickerson, J. L., Mc Cubbin, P. T. N., Brooks-Bartlett, J. C., and Garman, E. F., Protein Sci. 33, e 5005 (2024).10.1002/pro.500538923423 PMC 11196903 · doi ↗ · pubmed ↗
- 5Evans, P. R. and Murshudov, G. N., Acta Crystallogr. D 69, 1204–1214 (2013).10.1107/S 090744491300006123793146 PMC 3689523 · doi ↗ · pubmed ↗
- 6Garman, E. F., Acta Crystallogr. D 66, 339–351 (2010).10.1107/S 090744491000865620382986 PMC 2852297 · doi ↗ · pubmed ↗
- 7Gemmi, M. and Oleynikov, P., Z. Krist. - Cryst. Mater. 228, 51–58 (2013).10.1524/zkri.2013.1559 · doi ↗
- 8Gonen, T., Sliz, P., Kistler, J., Cheng, Y., and Walz, T., Nature 429, 193–197 (2004).10.1038/nature 0250315141214 · doi ↗ · pubmed ↗