The complete mitochondrial genomes of the freshwater mussel Ortmanniana ligamentina (Lamarck, 1819): male and female mitotypes
Katy Klymus, Jason Coombs, Dannise Ruiz-Ramos, Aaron Maloy, Christopher Barnhart

TL;DR
This study reveals both male and female mitochondrial genomes of a freshwater mussel using non-lethal methods and high-throughput sequencing.
Contribution
The paper introduces a non-lethal method to recover both male and female mitotypes in Ortmanniana ligamentina.
Findings
Both male and female mitotypes of Ortmanniana ligamentina were successfully recovered.
Non-lethal collection methods combined with high-throughput sequencing proved effective.
Increased male mitotype data improves understanding of Unionid genetic diversity.
Abstract
Freshwater mussels of the Unionida order are important to freshwater ecosystems but are highly imperiled worldwide. Improving our understanding of these species is crucial to their continued conservation. Some Unionid mussels exhibit double uniparental inheritance (DUI) in which individuals have two mitochondrial genomes. Of those species with DUI, sequences of the female mitotype are most prevalent in genetic databases. Here, we demonstrate the ability to recover both mitotypes of Ortmanniana ligamentina (Lamarck, 1819) from a non-lethal collection method coupled with high-throughput sequencing. Increased male mitotype sequence representation facilitates understanding Unionid genetic diversity and development of molecular tools for species detection.
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
 Figure 1
Figure 1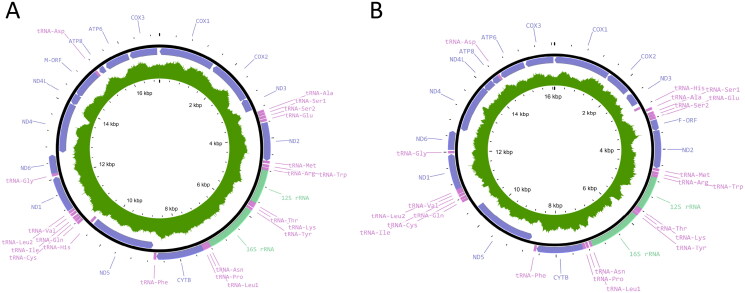 Figure 2
Figure 2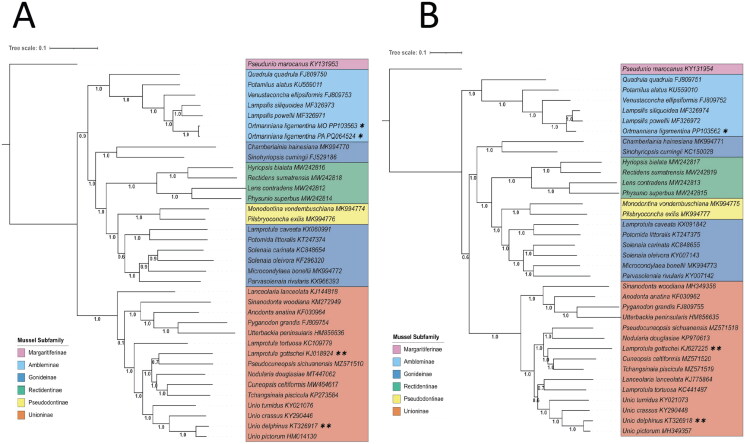 Figure 3
Figure 3| Subfamily | Species | Female accession | Male accession |
|---|---|---|---|
| Unioninae |
| ||
| Gonideinae |
| ||
| Unioninae |
| ||
| Rectidentinae |
| ||
| Gonideinae |
| ||
| Gonideinae |
| ||
| Unioninae |
| ||
| Unioninae |
| ||
| Ambleminae |
| ||
| Ambleminae |
| ||
| Rectidentinae |
| ||
| Gonideinae |
| ||
| Subfamily | Species | Female accession | Male accession |
| Pseudodontinae | |||
| Unioninae |
| ||
| Ambleminae |
| ||
| Ambleminae |
| N/A | |
| Gonideinae |
| ||
| Rectidentinae |
| ||
| Pseudodontinae |
| ||
| Ambleminae |
| ||
| Gonideinae |
| ||
| Unioninae |
| ||
| Margaritiferinae |
| ||
| Unioninae |
| ||
| Subfamily | Species | Female accession | Male accession |
| Ambleminae |
| ||
| Rectidentinae |
| ||
| Unioninae |
| ||
| Gonideinae |
| ||
| Gonideinae |
| ||
| Gonideinae |
| ||
| Unioninae |
| ||
| Unioninae |
| ||
| Rectidentinae |
| ||
| Unioninae |
| ||
| Unioninae |
| ||
| Unioninae |
| ||
| Unioninae |
|
- —Strategic Environmental Research and Development Program10.13039/100013316
- —U.S. Department of Defense10.13039/100000005
- —USGS Ecosystems Mission Area10.13039/100000203
- —U.S. Fish and Wildlife Service10.13039/100000202
- —Great Lakes Restoration Initiative
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsAquatic Invertebrate Ecology and Behavior · Mollusks and Parasites Studies · Marine Biology and Ecology Research
Introduction
A unique feature in some bivalves is the double uniparental inheritance (DUI) of mitochondrial DNA (Fisher and Skibinski 1990; Skibinski et al. 1994; Zouros et al. 1994; Walker et al. 2006). In this system, individuals have two separate mitochondrial genomes, or mitotypes, that are passed onto offspring. This differs from the more common mode of metazoan mitochondrial inheritance (strict maternal inheritance [SMI]) in which only the maternal mitochondrial genome is passed to offspring (Breton et al. 2007; Passamonti et al. 2011). Within bivalves with DUI, female mussels are believed to be homoplasmic, having the female mitotype in their somatic and gametic tissues, whereas male mussels are heteroplasmic (Garrido-Ramos et al. 1998; Soroka 2020). Male individuals’ somatic tissue contains the female mitotype, but their gametic tissue predominantly contains the male mitotype. Sequence divergence between these mitotypes is large, with differences of up to 52% between female and male mitotypes of the same species (Doucet-Beaupré et al. 2010; Soroka 2020). Among the bivalve taxa exhibiting DUI are species from at least three Unionida families (Unionidae, Margaritiferidae, and Hyriidae) (Soroka and Burzyński 2018).
Unionida are among the most threatened freshwater taxa (Lydeard et al. 2004; Galbraith et al. 2015; Graf and Cummings 2021; USFWS 2024). New tools for species monitoring, such as environmental DNA could be useful for their conservation (Prié et al. 2021; Klymus et al. 2021). Development of eDNA assays requires extensive genetic sequence data but currently male mitotype sequence data is limited in public databases. Here we assemble both female and male mitotypes of the Mucket, Ortmanniana ligamentina (Lamarck, 1819) as part of an effort to increase male mitotype representation in genetic databases.
Materials and methods
Sample collection and extraction
Individuals of Ortmanniana ligamentina (Figure 1(A)) were collected by the Missouri Department of Conservation (Saint Francis River, Missouri; 37° 10′ N; 90° 28′ W) 1 June 2020 and the Pennsylvania Fish and Boat Commission (Allegheny River, Pennsylvania; 41° 28′ N; 79° 29′ W) 24 May 2016. Each specimen was identified by shell morphology in the field and confirmed by later examination of the mantle and gill morphology by at least two workers familiar with the local range of variation and similar species (McMurray, Faiman, et al. 2012; Pennsylvania Fish & Boat Commission Division of Environmental Services 2018). Tissue samples consisting of 100 µL of gonadal fluid from Missouri mussels were obtained with a 1-mL syringe and 20–18 g needle. A hypodermic needle inserted into the anterior visceral mass was slowly rotated as the syringe withdrew fluid containing mainly male gametes and precursor cells. With care, samples can be taken without causing mortality (Shiver, 2002; Tsakiris et al. 2016). Microscopy of the gonad fluid was used to identify the sex of the individual mussels from Missouri (Figure 1(B,C)) and a sample from an identified male individual was used in subsequent sequencing. Sample of a gill biopsy collected from a female Pennsylvania specimen was also sequenced to validate the data collected from the syringe method. Samples were preserved in 95% EtOH. Total genomic DNA was extracted with either the IBI Scientific gMAX Mini Genomic DNA kit or Qiagen DNeasy Blood and Tissue kit. Extracted DNA and tissue samples (B065) from Missouri are archived at the Columbia Environmental Research Center, Columbia, Missouri, USA (https://www.usgs.gov/centers/columbia-environmental-research-center, Dr. Catherine Richter, [email protected]). Extracted DNA (B16-023) from the Pennsylvania specimen is archived at the Northeast Fishery Center, Lamar, Pennsylvania, USA Center (https://www.fws.gov/office/northeast-fishery-center).
(A) Mucket mussel. Photo taken by Katy Klymus at Columbia Environmental Research Center, Missouri, USA. (B) Magnified image (40X) of male gametic fluid removed via a syringe. The material was used to verify the sex of individual mussels and for sequencing. Sequencing of the male gametic tissue ensured we recovered the male mitotype genome. We were also able to recover the female mitotype genome from the same sample. Photo taken by Chris Barnhart at Missouri State University. (C) Magnified image (20X) of female gametic fluid removed via a syringe. The material was used to verify the sex of individual mussels. We did not sequence female gametic tissue. Photo taken by Chris Barnhart at Missouri State University.
Genomic sequencing, assembly, and annotation
Library preparation and sequencing took place at either the University of Missouri Genomics Technology Core or the Northeast Fishery Center. Libraries were prepared from genomic DNA per manufacturer’s protocol with reagents supplied in the Illumina DNA Prep or Nextera XT preparation Kits. Final libraries were bead purified and sequenced on a NovaSeq 6000 platform (Illumina, San Diego, CA) with a 2 × 150 or 2 × 250 bp paired-end read kit. Adapters and low-quality bases were removed before merging paired-end reads. Merged reads were de novo assembled using the Tadpole assembler in Geneious to obtain a draft list of contigs. From the Missouri specimen, a total of 502,682,066 raw sequences were recovered resulting in 145,843,052 merged reads. From the Pennsylvania specimen, a total of 15,449,292 reads were recovered, resulting in 6,346,335 merged reads. Gene annotations from Lampsilis siliquoidea (GenBank accession MF326973) were temporarily mapped based on similarity to a subset of contigs which fell within the expected mitogenome size range. Temporary annotations were used to identify contigs of mitochondrial origin. Sequences of mitochondrial origin served as the seed sequence against which all merged reads were mapped over successive iterations. Reads successfully mapped were then de novo assembled to recover two circular mitochondrial genomes. Gene locations were determined based on similarity to other Unionid species and verified using MitoAnnotator (Iwasaki et al. 2013). Gene arrangement and presence of the male and female open reading frames were used to identify the longer mitogenome as the male mitotype. All trimming and assembly steps were conducted using Geneious Prime version 2022.0.2 (Biomatters Ltd., Auckland, New Zealand). The average read depth of the mitogenomes from the Missouri specimen was 60 while that of the Pennsylvania specimen was 146. Genome maps were made using Proksee (Grant et al. 2023) (Figure 2(A,B), Supplemental Figures S1 and S2).
Mitogenome maps of (A) the male mitotype (accession number PP103562) from the Missouri individual (B065) and (B) the female mitotype (accession number PP103563) from the Missouri individual (B065). The map of the female mitotype from the Pennsylvania individual B16-023 (accession number PQ064524) is the same as that of the female mitotype from Missouri. The outer track shows the gene features and positions, with purple representing the coding DNA sequence, pink representing tRNAs, and light green representing rRNAs. The dark green inner track represents read depth for each assembly.
The new genomes were relationally assessed through generation of phylogenetic trees using sequence data from GenBank of 36 Unionida species with complete male and female mitogenome data (Table 1). Genes were extracted and aligned within Geneious, and concatenated and exported in SequenceMatrix version 1.9 (Vaidya et al. 2011). Trees were generated with the FastTree (Price et al. 2009, 2010) plugin in Geneious using default settings and rooted with Pseudunio marocanus a species from the subfamily Margaritiferinae. Tree visualization was with Interactive Tree of Life (iTOL) (Letunic and Bork 2021).
Results
Sampling of gonadal fluid resulted in genomic DNA extract containing 0.12% mitochondrial reads for the male mitotype and 0.0034% mitochondrial reads of the female mitotype. This discrepancy is expected, as less somatic tissue containing the female mitotype was present in the collected gonadal fluid.
From the Missouri specimen, the assembled male mitogenome (GenBank accession PP103562) was 17,015 bp, and the female mitogenome (GenBank accession PP103563) was 16,070 bp (Figure 2(A,B), Supplementary Figures S1 and S2). The length of the COX2 region heavily influenced the difference in length; the female COX2 region was 681 bp, while the male COX2 was 1242 bp. The female mitogenome of the Pennsylvania specimen (GenBank accession PQ064524) was 16,075 bp in length. Phylogenetic analyses place the two female mitotype genomes as sister to one another within the Ambleminae subfamily and sister to Lampsilis siliquoidea and Lampsilis powellii (Figure 3(A)). Similarly, the sequenced male mitotype genome was placed among male mitotype genomes of other Ambleminae individuals (Figure 3(B)). The trees of male and female mitotypes have similar topology, with sequences grouping in the same subfamily clades. Interestingly, the Unioninae clade changes places with the Ambleminae clade between the two mitotype topologies, but this could be an artifact of limited sampling.
Inferred phylogenetic trees based on the 13 coding genes and two ribosomal RNAs of the mitochondrial genome using approximately maximum likelihood methods through FastTree (Price et al. 2009, 2010). Numbers denote Shimodaira–Hasegawa test support values of the associated split. An asterisk () indicates the individuals genotyped in this study. Subfamily names are indicated to the right of each clade. Double asterisks (**) indicate two individuals with different subfamily designations than that in which they group with. The sequences and their accession numbers used can be found in Table 1. (A) Phylogenetic tree of the female mitotypes. (B) Phylogenetic tree of male mitotypes.*
Although not a focus of this study, the use of sequences from GenBank reveals discrepancies between current taxonomy using MolluscaBase (MolluscaBase 2025) and the species identification of the downloaded sequences for two individuals (Unio delphinus (KT326917/KT32918) and Lamprotula gottschei (KJ018924/KJ627225) (Figure 3(A,B), Table 1). These results suggest that these specimens might have been mis-identified in the original study or that further systematic study of these species is warranted.
Discussion and conclusions
We report the first complete male mitotype genome of O. ligamentina (GenBank accession PP103562) and 2 complete female mitotype genomes (GenBank accession PP103563 and PQ064524). Our study’s phylogenetic placement of all three mitogenomes is supported by the current literature in that O. ligamentina is in the Ambleminae subfamily and sister to Lampsilis siliquoidea and Lampsilis powellii (Inoue et al. 2020; Stodola et al. 2021). We also demonstrate the ability to sequence and recover both mitotypes from non-lethal collection methods, using high throughput sequencing of gonadal tissue taken with a syringe. Previously, Sanger sequencing of the different mitotypes required sacrificing the animal to dissect clean gonadal tissue to avoid female mitotype contamination (Breton et al. 2006). This non-lethal method can increase the number of published male mitotype genomes which in turn can aid the study of DUI evolution and conservation practice. For instance, male mitotype sequences can be used to design molecular assays that target the identification of mussel male gametes (Prié et al. 2021). This application could identify reproductive triggers and spawning events which will be useful in the captive propagation being conducted to reintroduce populations in the wild.
Supplementary Material
SuppFig1_031025.docx
SuppFig2_031025.docx
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Breton S, Burger G, Stewart DT, Blier PU. 2006. Comparative analysis of gender-associated complete mitochondrial genomes in marine mussels (Mytilus spp.). Genetics. 172(2):1107–1119. doi:10.1534/genetics.105.047159.16322521 PMC 1456209 · doi ↗ · pubmed ↗
- 2Breton S, Beaupré HD, Stewart DT, Hoeh WR, Blier PU. 2007. The unusual system of doubly uniparental inheritance of mt DNA: isn’t one enough? Trends Genet. 23(9):465–474. doi:10.1016/j.tig.2007.05.011.17681397 · doi ↗ · pubmed ↗
- 3Breton S, Beaupré HD, Stewart DT, Piontkivska H, Karmakar M, Bogan AE, Blier PU, Hoeh WR. 2009. Comparative mitochondrial genomics of freshwater mussels (Bivalvia: unionoida) with doubly uniparental inheritance of mt DNA: gender-specific open reading frames and putative origins of replication. Genetics. 183(4):1575–1589. doi:10.1534/genetics.109.110700.19822725 PMC 2787441 · doi ↗ · pubmed ↗
- 4Breton S, Stewart DT, Shepardson S, Trdan RJ, Bogan AE, Chapman EG, Ruminas AJ, Piontkivska H, Hoeh WR. 2011. Novel protein genes in animal mt DNA: a new sex determination system in freshwater mussels (Bivalvia: unionoida)? Mol Biol Evol. 28(5):1645–1659. doi:10.1093/molbev/msq 345.21172831 PMC 3107663 · doi ↗ · pubmed ↗
- 5Burzyński A, Soroka M. 2018. Complete paternally inherited mitogenomes of two freshwater mussels Unio pictorum and Sinanodonta woodiana (Bivalvia: unionidae). Peer J. 6:e 5573. doi:10.7717/peerj.5573.30221094 PMC 6138038 · doi ↗ · pubmed ↗
- 6Burzyński A, Soroka M, Mioduchowska M, Kaczmarczyk A, Sell J. 2017. The complete maternal and paternal mitochondrial genomes of Unio crassus: mitochondrial molecular clock and the overconfidence of molecular dating. Mol Phylogenet Evol. 107(2017):605–608. doi:10.1016/j.ympev.2016.12.007.27956259 · doi ↗ · pubmed ↗
- 7Doucet-Beaupré H, Breton S, Chapman EG, Blier PU, Bogan AE, Stewart DT, Hoeh WR. 2010. Mitochondrial phylogenomics of the Bivalvia (Mollusca): searching for the origin and mitogenomic correlates of doubly uniparental inheritance of mt DNA. BMC Evol Biol. 10:50. doi:10.1186/1471-2148-10-50.20167078 PMC 2834691 · doi ↗ · pubmed ↗
- 8Fisher C, Skibinski DOF. 1990. Sex-biased mitochondrial DNA heteroplasmy in the marine mussel Mytilus. Proc R Soc Lond B. 242:149–156. doi:10.1098/rspb.1990.0118. · doi ↗