Microglial reprogramming: a potential new frontier in enhancing immunotherapy for melanoma brain metastasis
Noam Savion‐Gaiger, Dorin Bar‐Ziv, Harriet Kluger

TL;DR
This paper explores how reprogramming brain microglia could improve immunotherapy for melanoma that spreads to the brain.
Contribution
The study identifies NF-κB/RelA signaling as a key driver of microglial shift and proposes targeting it to enhance antitumor immunity.
Findings
Microglia initially suppress tumors but later promote them in melanoma brain metastases.
NF-κB/RelA signaling drives the shift in microglial function.
Inhibiting this pathway reprograms microglia to support antitumor immunity.
Abstract
The brain is a common site of metastatic dissemination in advanced melanoma. Due to limited access to samples from human brain metastases, our understanding of the tumor microenvironment in the brain lags behind that of other sites, and murine studies are therefore highly informative. Rodriguez‐Baena et al. conducted elegant studies of myeloid cells within melanoma brain metastases, demonstrating their plasticity and changes before and after colonization by melanoma cells. The immune‐inhibitory changes in microglial cells may be reversed or mitigated by inhibition of RelA/NF‐κB. Microglia act as tumor suppressors during brain metastasis colonization but shift to a tumor‐promoting role after melanoma brain metastases form. NF‐κB/RelA signaling emerges as a key driver of this phenotypic shift. Targeting this pathway reprograms microglia into a pro‐inflammatory state, enhancing antitumor…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
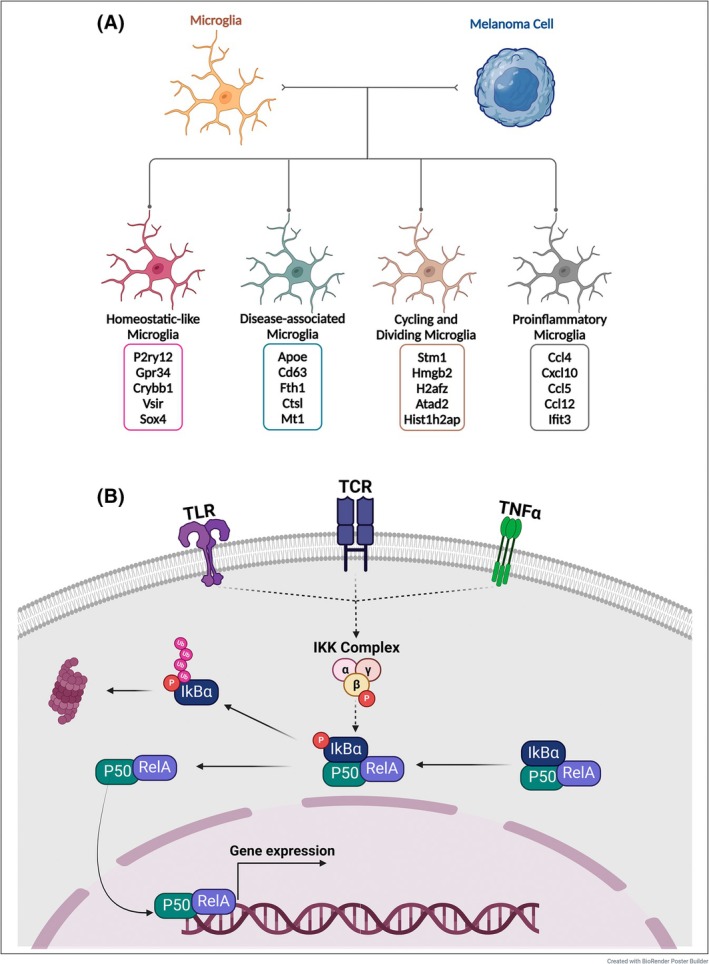 Fig. 1
Fig. 1- —NIH 10.13039/100000002
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsNeuroinflammation and Neurodegeneration Mechanisms · Immune cells in cancer · Immunotherapy and Immune Responses
Background and perspective
1
The spread of melanoma to the brain is challenging if unresponsive to immune checkpoint inhibitors (ICIs) [1]. While innumerable studies have evaluated the tumor microenvironment (TME) in extracranial metastases, it is relatively understudied in the brain due to limited access to human tissue. Small studies demonstrated differences in T‐cell content and tumor vasculature in brain versus extracranial sites in patients treated with ICIs [2, 3]. Single‐cell RNA‐sequencing (sc‐RNA‐seq) technologies have enhanced our understanding, showing that melanoma brain metastases (MBM) are enriched with myeloid cells [4]. Differences between brain‐resident microglia and peripheral myeloid cells have been well‐documented [5]. In MBM, the role of microglia has been examined, with studies demonstrating both protumorigenic and antitumorigenic roles, as reviewed [6].
In a novel study by Rodriguez‐Baena et al. [7], distinct, phenotypically diverse microglial subpopulations were identified. They showed that microglia can transition between pro‐inflammatory/antitumor states and anti‐inflammatory, tumor‐promoting phenotypes in the presence of MBM, largely driven by the RelA/NF‐κB pathway.
Microglia: Janus‐faced regulators of brain metastasis
2
Much can be learned about the role of microglia from glioblastomas, where extracranial factors are less important and tumor tissue is often available. Comparisons between primary brain tumors and metastases have shown that divergent myeloid populations have variable effects on T cells and tumor cells, suggesting that myeloid modulation might be preferred to myeloid ablation for optimizing the antitumor immune response [8].
Using two syngeneic murine melanoma models of colonization of the brain, elegant myeloid manipulation methods, and extensive scRNA‐seq of murine MBM, Rodriguez‐Baena identified distinct populations with two major types of tumor‐associated macrophages (TAMs): microglia (expressing Tmem119, P2ry12, Gpr34, Sall1) and nonmicroglial myeloid cells (expressing Lyz2, Itga4, Cd74, Plac8) [7]. Microglia were a more abundant subset within the macrophage population, consistent with studies of the MBM microenvironment [9]. Depleting microglia prior to tumor colonization increased MBM burden but did not alter macrophage or peripheral immune cell infiltration. However, when TAMs were depleted after MBM formation, tumor burden was decreased, indicating that TAMs actively contribute to tumor growth once MBMs are established.
Four microglial subpopulations were identified: pro‐inflammatory microglia with enhanced expression of T/NK cell attractant chemokines and antigen presentation genes, disease‐associated microglia with high translational activity, and two clusters linked to glial homeostasis and cell division (Fig. 1A). These important findings still require validation in models of metastasis rather than colonization, as it would be impossible to validate them in humans.
(A) In the presence of melanoma brain metastases, tumor‐associated microglia differentiate into four distinct subpopulations: Homeostatic‐like microglia, characterized by high transcription of genes and pathways associated with glial cell function and homeostasis; Disease‐associated microglia, exhibiting increased translational activity; Proliferating microglia, which include cycling and dividing microglia; and Pro‐inflammatory microglia, marked by enhanced chemokine expression and antigen presentation and strong predicted CD8+ and NK cell interactions. RelA deletion in conditional knockout mice resulted in microglia that displayed an enrichment of homeostatic‐like signature and pro‐inflammatory microglial signatures, along with a reduction in disease associated microglia‐like gene transcription. (B) The canonical NF‐κB pathway is activated by pro‐inflammatory cytokines (e.g., TNF‐α, IL‐1), pattern recognition receptors (e.g., TLRs), and antigen receptors. Upon stimulation, the receptor‐associated complex recruits and activates the IκB kinase (IKK) complex, consisting of IKKα, IKKβ, and IKKγ (NEMO). IKKβ phosphorylates IκBα, leading to its ubiquitination and subsequent degradation by the proteasome. This degradation releases the NF‐κB heterodimer, RelA (p65)/p50, allowing it to translocate into the nucleus. Once inside the nucleus, NF‐κB binds to κB‐responsive elements in target genes, promoting the transcription of pro‐inflammatory cytokines, chemokines, and cell survival genes.
Microglial NF‐κB activation can enhance brain metastasis growth after colonization
3
A striking difference between tumor‐associated microglia and nonmicroglial macrophages in the models studied by Rodriguez‐Baena et al. was in their expression of RelA target genes. RelA, a subunit of NF‐κB, is a transcription factor that plays a central role in regulating inflammation, cell survival, and apoptosis signaling in immune cells (Fig. 1B). With RelA depletion after MBM formation, tumor burden was significantly decreased without altering the number of microglia or nonmicroglial macrophages. Administration of dehydroxymethylepoxyquinomicin (DHMEQ), a brain‐penetrant NF‐κB inhibitor, decreased MBM burden and prolonged survival in mice, with no effect on subcutaneous tumors. DHMEQ has challenging pharmacological properties and has not been administered to humans, although it has activity in numerous preclinical models of cancer [10]. However, numerous drugs approved for other purposes inhibit NF‐κB and can potentially be repurposed for treating MBMs [11].
Targeting NF‐κB reprograms microglia and enhances antitumor immunity
4
RelA deletion in conditional knockout mice reduced protumorigenic disease‐associated microglia and increased the proportion of pro‐inflammatory microglia, combined with increased T and NK cell‐attracting factors and an influx of CD8^+^ T and NK cells. Given the relatively poor T‐cell infiltration in human MBM and the potential associated lack of response to ICIs, microglial manipulation might provide an avenue to overcome this challenge [12].
NF‐κB modulation for enhancing ICI activity was also studied by Rodriguez‐Baena. While RelA wild‐type mice exhibited a reduced metastatic burden in the NRas mutant following anti‐PD‐L1, combining DHMEQ with anti‐PD‐L1 enhanced the intracranial tumor response. Similar findings were demonstrated when combining anti‐PD‐1 with anti‐CTLA‐4. While improvements were seen, effects on survival were not reported, and it is unclear whether any mice had complete tumor rejection, highlighting the differences between murine models and humans, where complete tumor rejection can be seen with ICIs.
Unanswered questions and future directions
5
This study provides a deep characterization of myeloid subsets in MBM with solid evidence that NF‐κB pathway activation in microglia drives the transition to a protumor state when MBM are present, suggesting a novel avenue to treat MBM. Previous research on protumorigenic and pro‐inflammatory microglial phenotypes has primarily focused on glioblastoma and brain metastasis from lung and breast cancer. While this study did not compare melanoma to other tumors, further exploration of this phenomenon in other malignancies is warranted, as microglial modulation might depend on interactions with transformed melanocytes, cells of neural crest origin.
The leap between preclinical and clinical studies is often limited by the relevance of the murine models. B16/F10 does not mimic human melanoma genetically, while the NRas model reflects the biology of up to 20% of melanomas [13]. Carotid artery injection and intracranial injections guarantee the formation of brain deposits but do not reflect the metastatic process, and these provocative results support the need for further work in models of metastasis, which are challenging, as reviewed [14].
Research on MBM has been hampered by limited human tissue to validate results. Some groups have generated human tissue resources in recent years, providing an opportunity to verify that NF‐κB expression varies in microglia from tumors of responders and nonresponders to ICIs [1].
While microglial subtypes resemble the traditional M1/M2 classification of peripheral macrophages—where M1 are pro‐inflammatory and M2 are pro‐tumorigenic—this binary model fails to fully capture their plasticity. The identification of additional microglial subtypes in this study underscores the complexity of their functional states, necessitating further investigation into their roles within the TME.
These findings have important clinical implications. The evidence provided by Rodriguez‐Baena of the role of NF‐κB in microglia in MBM warrants considering human investigation using novel NF‐κB or existing drugs that might be repurposed [15].
Conflict of interest
Dr. Kluger has received research grants from Pfizer, Apexigen, and Merck and personal fees from Iovance, Merck, Chemocentryx, Bristol‐Myers Squibb, Signatero, Gigagen, GI reviewers, Pliant Therapeutics, Esai, Invox, Wherewolf, Teva, and Replimmune, all unrelated to this work.
Author contributions
All three authors contributed to conceptualizing the commentary, and to writing and proofreading the manuscript.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Karz A , Dimitrova M , Kleffman K , Alvarez‐Breckenridge C , Atkins MB , Boire A , et al. Melanoma central nervous system metastases: an update to approaches, challenges, and opportunities. Pigment Cell Melanoma Res. 2022;35(6):554–572.35912544 10.1111/pcmr.13059 PMC 10171356 · doi ↗ · pubmed ↗
- 2Kluger HM , Chiang V , Mahajan A , Zito CR , Sznol M , Tran T , et al. Long‐term survival of patients with melanoma with active brain metastases treated with pembrolizumab on a phase II trial. J Clin Oncol. 2019;37(1):52–60.30407895 10.1200/JCO.18.00204 PMC 6354772 · doi ↗ · pubmed ↗
- 3Weiss SA , Djureinovic D , Wei W , Tran T , Austin M , Markowitz J , et al. A phase 2 trial of pembrolizumab in combination with bevacizumab for untreated melanoma brain metastases. J Clin Oncol. 2025. 10.1200/JCO-24-02219 PMC 1205841540048689 · doi ↗ · pubmed ↗
- 4Biermann J , Melms JC , Amin AD , Wang Y , Caprio LA , Karz A , et al. Dissecting the treatment‐naïve ecosystem of human melanoma brain metastasis. Cell. 2022;185(14):2591–2608.e 30.35803246 10.1016/j.cell.2022.06.007PMC 9677434 · doi ↗ · pubmed ↗
- 5Ginhoux F , Greter M , Leboeuf M , Nandi S , See P , Gokhan S , et al. Fate mapping analysis reveals that adult microglia derive from primitive macrophages. Science. 2010;330(6005):841–845.20966214 10.1126/science.1194637 PMC 3719181 · doi ↗ · pubmed ↗
- 6Izraely S , Witz IP . Site‐specific metastasis: a cooperation between cancer cells and the metastatic microenvironment. Int J Cancer. 2021;148(6):1308–1322.32761606 10.1002/ijc.33247 PMC 7891572 · doi ↗ · pubmed ↗
- 7Rodriguez‐Baena FJ , Marquez‐Galera A , Ballesteros‐Martinez P , Castillo A , Diaz E , Moreno‐Bueno G , et al. Microglial reprogramming enhances antitumor immunity and immunotherapy response in melanoma brain metastases. Cancer Cell. 2025;43:413–427.e 9.39919736 10.1016/j.ccell.2025.01.008 · doi ↗ · pubmed ↗
- 8Klemm F , Maas RR , Bowman RL , Kornete M , Soukup K , Nassiri S , et al. Interrogation of the microenvironmental landscape in brain tumors reveals disease‐specific alterations of immune cells. Cell. 2020;181(7):1643–1660.e 17.32470396 10.1016/j.cell.2020.05.007PMC 8558904 · doi ↗ · pubmed ↗