Lactobacillus delbrueckii subsp. allosunkii and lactis as emerging human uropathogens in elderly patients
François Guérin, Mohamed Sassi, Francois Gravey, Asma Zouari, Benjamin Quenet, Maxime Lecourt, Pauline Ract, Charlotte Michaux, Michel Auzou, Christophe Isnard, Vincent Cattoir

TL;DR
Lactobacillus delbrueckii subspecies allosunkii and lactis are emerging causes of UTIs in elderly patients, especially women, and may require changes in clinical practices.
Contribution
This is the largest case series demonstrating L. delbrueckii as a uropathogen and provides insights into its antimicrobial susceptibility and subspecies distribution.
Findings
L. delbrueckii isolates were mostly susceptible to β-lactams but intrinsically resistant to fosfomycin and metronidazole.
Most isolates belonged to subspecies allosunkii (83%), with a minority being lactis (17%).
Two isolates were resistant to tetracycline and carried the tet(W) gene.
Abstract
Lactobacillus delbrueckii has been considered a very rare cause of human urinary tract infections (UTIs). However, little is known about its clinical significance and antimicrobial susceptibility, and genomic data from clinical isolates are lacking. This study aimed at analyzing clinical, microbiological, and genomic data of L. delbrueckii urinary isolates. All L. delbrueckii isolates collected from patients hospitalized in a French university hospital from 2014 to 2016 were included. Clinical and biological data were gathered. Species identification was performed by matrix-assisted laser desorption/ionization time-of-flight (MALDI-TOF) mass spectrometry, and MICs were determined using the broth microdilution method. Whole genome sequencing (WGS) was conducted (Illumina MiSeq 2 × 300 bp), and genomes were compared using three approaches (multilocus sequence typing [MLST], average…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
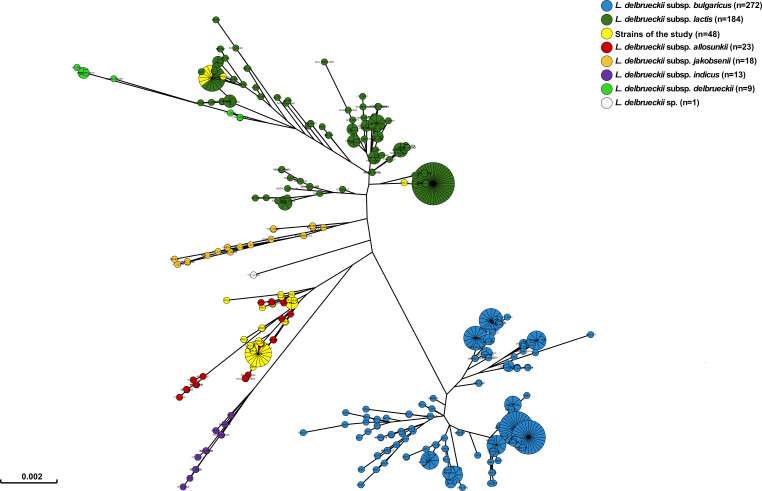 Fig 1
Fig 1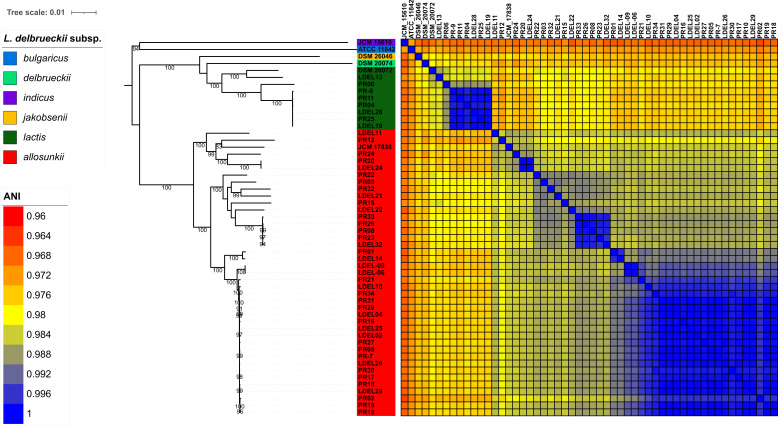 Fig 2
Fig 2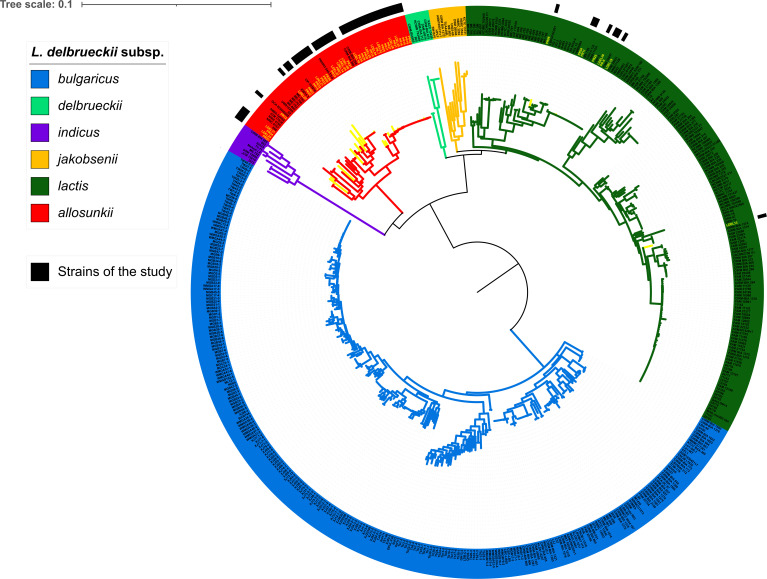 Fig 3
Fig 3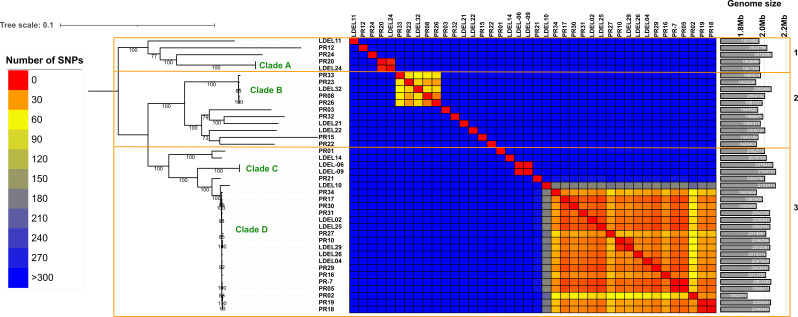 Fig 4
Fig 4| Characteristics | No. of patients (%) |
|---|---|
| Sex | |
| Male | 2 (4) |
| Female | 46 (96) |
| Age (years), median (range) | 84 (54–96) |
| Origin | |
| Community-acquired | 40 (83) |
| Hospital-acquired | 8 (17) |
| Predisposing urologic disease | 3 (6) |
| Indwelling urinary catheter | 11 (23) |
| Systemic underlying condition(s) | |
| Heart disease | 33 (69) |
| Immunosuppression | 29 (60) |
| Mellitus diabetes | 15 (31) |
| Malignancy | 14 (29) |
| Dementia | 12 (25) |
| Stroke | 6 (13) |
| None | 4 (8) |
| Diagnosis retained by the physician | |
| Infection | 21 (44) |
| Colonization/contamination | 27 (6) |
| Antibiotic treatment for UTI | |
| Ceftriaxone | 9 (19) |
| Amoxicillin-clavulanic acid | 4 (8) |
| Amoxicillin | 3 (6) |
| Ofloxacin | 3 (6) |
| Ciprofloxacin | 1 (2) |
| Piperacillin-tazobactam | 1 (2) |
| Direct microscopic examination | |
| Leucocyturia (WBC count >104/mL) | 45 (94) |
| Gram staining positive for Gram-positive rods | 43 (90) |
| Quantitative culture | |
| Significant | 48 (100) |
| Concomitant uropathogen(s) | 27 (56) |
| Antibiotic | MIC (mg/L) | CLSI susceptibility breakpoint (mg/L) | % of susceptible strains | ||
|---|---|---|---|---|---|
| MIC50 | MIC90 | Range | |||
| Ampicillin | 0.12 | 0.25 | 0.03–0.5 | 8 | 100 |
| Cefotaxime | 0.5 | 1 | 0.03–2 | – | – |
| Imipenem | 0.06 | 0.12 | 0.01–0.5 | 0.5 | 100 |
| Gentamicin | 0.5 | 1 | 0.12–2 | – | – |
| Erythromycin | 0.06 | 0.12 | 0.01–0.5 | 0.5 | 100 |
| Levofloxacin | 4 | 16 | 1–16 | - | - |
| Vancomycin | 0.25 | 0.5 | 0.03–2 | 2 | 100 |
| Teicoplanin | 0.12 | 0.12 | 0.12–1 | – | – |
| Linezolid | 2 | 2 | 0.5–2 | 4 | 100 |
| Tetracycline | 1 | 2 | 0.25–>16 | – | – |
| Tigecycline | 0.25 | 0.5 | 0.12–0.5 | – | – |
| Daptomycin | 0.12 | 0.5 | 0.06–0.5 | 4 | 100 |
| Cotrimoxazole | 0.12 | 0.25 | 0.12–8 | – | – |
| Nitrofurantoin | 16 | 32 | 8–32 | – | – |
| Fosfomycin | >256 | >256 | >256 | – | – |
| Metronidazole | >256 | >256 | >256 | – | – |
| Strain ID | Assembly size (Mb) | GC% | No. of CDS | Resistance genes | No. of contigs | N50 | L50 | Coverage (X) | GenBank accession number | |
|---|---|---|---|---|---|---|---|---|---|---|
|
|
| 2.06 | 49.77 | 1,934 | – | 48 | 74,998 | 8 | 255 |
|
|
|
| 2.05 | 49.77 | 1,927 | – | 49 | 73,394 | 7 | 332 |
|
|
|
| 2.08 | 49.84 | 1,985 | 72 | 54,390 | 12 | 166 |
| |
|
|
| 2.11 | 49.77 | 2,004 | 64 | 65,483 | 11 | 130 |
| |
|
|
| 2.10 | 49.66 | 2,014 | – | 73 | 53,111 | 13 | 243 |
|
|
|
| 1.95 | 50.17 | 1,867 | – | 66 | 68,921 | 10 | 273 |
|
|
|
| 1.94 | 49.57 | 1,927 | – | 96 | 42,923 | 13 | 280 |
|
|
|
| 2.02 | 49.95 | 1,933 | – | 73 | 58,626 | 14 | 254 |
|
|
|
| 1.97 | 49.76 | 1,947 | – | 81 | 46,214 | 16 | 316 |
|
|
|
| 1.96 | 49.92 | 1,882 | – | 87 | 40,366 | 14 | 243 |
|
|
|
| 2.01 | 49.92 | 1,925 | – | 76 | 46,593 | 10 | 310 |
|
|
|
| 1.96 | 50.18 | 1,864 | – | 44 | 102,800 | 7 | 322 |
|
|
|
| 2.05 | 49.76 | 1,926 | – | 47 | 74,998 | 7 | 287 |
|
|
|
| 2.02 | 49.94 | 1,888 | – | 50 | 68,962 | 8 | 302 |
|
|
|
| 1.95 | 49.74 | 1,937 | – | 86 | 39,415 | 17 | 276 |
|
|
|
| 2.05 | 49.77 | 1,936 | – | 54 | 68,962 | 9 | 329 |
|
|
|
| 2.06 | 49.62 | 1,989 | – | 109 | 28,910 | 23 | 239 |
|
|
|
| 2.00 | 49.93 | 1,928 | – | 74 | 47,985 | 14 | 268 |
|
|
|
| 1.84 | 49.91 | 1,788 | – | 125 | 23,936 | 24 | 305 |
|
|
|
| 1.94 | 50.03 | 1,863 | – | 113 | 24,343 | 24 | 312 |
|
|
|
| 1.87 | 50.02 | 1,858 | – | 102 | 32,919 | 19 | 347 |
|
|
|
| 2.05 | 49.78 | 1,929 | – | 52 | 74,998 | 9 | 382 |
|
|
|
| 1.86 | 49.98 | 1,818 | – | 78 | 41,365 | 13 | 341 |
|
|
|
| 2.06 | 49.76 | 1,939 | – | 50 | 73,394 | 8 | 130 |
|
|
|
| 2.00 | 49.74 | 1,937 | – | 117 | 28,753 | 21 | 329 |
|
|
|
| 1.95 | 49.70 | 1,934 | – | 84 | 46,215 | 14 | 102 |
|
|
|
| 2.05 | 49.78 | 1,931 | – | 61 | 65,416 | 12 | 283 |
|
|
|
| 1.95 | 49.75 | 1,942 | – | 96 | 33,599 | 21 | 310 |
|
|
|
| 2.02 | 50.07 | 1,909 | – | 69 | 52,964 | 11 | 250 |
|
|
|
| 1.94 | 49.85 | 1,860 | – | 95 | 46,718 | 15 | 309 |
|
|
|
| 2.01 | 49.87 | 1,912 | – | 56 | 67,249 | 8 | 316 |
|
|
|
| 1.98 | 50.02 | 1,873 | – | 55 | 73,383 | 7 | 270 |
|
|
|
| 2.05 | 49.78 | 1,936 | – | 66 | 64,800 | 10 | 288 |
|
|
|
| 2.06 | 49.76 | 1,933 | – | 51 | 73,393 | 8 | 247 |
|
|
|
| 1.96 | 50.17 | 1,869 | – | 53 | 90,809 | 9 | 244 |
|
|
|
| 2.00 | 49.84 | 1,913 | – | 75 | 54,621 | 12 | 197 |
|
|
|
| 1.93 | 50.03 | 1,843 | – | 86 | 35,400 | 15 | 380 |
|
|
|
| 1.92 | 49.94 | 1,844 | – | 101 | 27,694 | 20 | 346 |
|
|
|
| 2.07 | 49.91 | 1,972 | – | 57 | 84,947 | 8 | 293 |
|
|
|
| 1.98 | 49.72 | 1,957 | – | 76 | 41,571 | 16 | 333 |
|
|
|
| 1.98 | 49.84 | 1,900 | – | 112 | 27,934 | 22 | 285 |
|
|
|
| 2.01 | 49.95 | 1,896 | – | 54 | 73,394 | 8 | 276 |
|
|
|
| 2.04 | 49.78 | 1,925 | – | 57 | 68,962 | 8 | 155 |
|
|
|
| 1.93 | 50.14 | 1,832 | – | 122 | 26,713 | 23 | 205 |
|
|
|
| 2.05 | 49.77 | 1,933 | – | 52 | 67,160 | 8 | 275 |
|
|
|
| 1.99 | 49.90 | 1,912 | – | 66 | 57,585 | 10 | 256 |
|
|
|
| 1.97 | 49.89 | 1,884 | – | 114 | 24,314 | 22 | 317 |
|
|
|
| 1.93 | 50.03 | 1,833 | – | 56 | 59,273 | 11 | 223 |
|
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsProbiotics and Fermented Foods · Urinary Tract Infections Management · Gut microbiota and health
INTRODUCTION
Urinary tract infections (UTIs) are among the most common human infections in both community and hospital settings, affecting 150 million people each year worldwide, with significant morbidity and high medical costs (1). In both settings, Enterobacterales are responsible for the vast majority of UTIs, predominantly uropathogenic Escherichia coli isolates (1). Besides Gram-negative bacilli, UTIs can also be caused by Gram-positive bacteria such as enterococci, Staphylococcus saprophyticus, or Streptococcus agalactiae (2). Furthermore, emerging Gram-positive species have recently been described as true uropathogens: Actinotignum (formerly Actinobaculum) schaalii and Aerococcus spp. (A. urinae and A. sanguinicola) (2).
Lactobacilli are non-spore-forming, facultatively anaerobic or microaerophilic, non-nitrate-reducing, non-motile Gram-positive rods. These microorganisms are ubiquitous and widespread commensals in human and animal microbiota. In humans, they are usually considered non-pathogenic as they are common inhabitants of the oropharynx, the gastrointestinal tract, and the female genital tract. Like other lactic acid bacteria (e.g., pediococci, lactococci) and bifidobacteria, they are widely used as starter cultures in the food industry (for fermented milk products or sausages) or as probiotic preparations (3). Due to their fastidious growth under aerobic conditions and their resemblance to bacterial species of the resident microbiota, lactobacilli have frequently been overlooked or regarded as contaminants (4). Additionally, it has long been challenging to identify them to the species level using conventional phenotypic-based methods (5, 6). Although the advent of matrix-assisted laser desorption/ionization time-of-flight (MALDI-TOF) mass spectrometry has greatly improved their identification, molecular identification is still required in some cases (7). Nonetheless, they can occasionally cause invasive infections, such as bacteremia and endocarditis, which are the most frequent, followed by intra-abdominal and respiratory infections, UTIs, endophthalmitis, chorioamnionitis, and meningitis as well as various abscesses and wound infections (4–6, 8–14). The main species recovered from human infections, knowing that the genus Lactobacillus was extensively modified and several species have been reassigned to novel genus names, include Lacticaseibacillus rhamnosus (formerly Lactobacillus rhamnosus), Lacticaseibacillus casei (formerly Lactobacillus casei), and Lactobacillus gasseri followed by Limosilactobacillus fermentum (formerly Lactobacillus fermentum), Lacticaseibacillus paracasei (formerly Lactobacillus paracasei), Lactobacillus jensenii, Lactobacillus acidophilus, and Lactiplantibacillus plantarum (formerly Lactobacillus plantarum) (4–6, 8–11, 13–15). These infections are often associated with underlying conditions such as immunosuppression, neutropenia, cancer, diabetes, previous surgery or endoscopy, invasive procedures (central venous or urinary catheterization, intubation/ventilation, dental procedure), prosthetic material, renal failure, or prolonged ineffective antibiotic therapy (4, 5, 8).
Lactobacillus delbrueckii is the type species of the genus Lactobacillus and includes six different subspecies, namely L. delbrueckii subsp. bulgaricus, L. delbrueckii subsp. delbrueckii, L. delbrueckii subsp. indicus, L. delbrueckii subsp. jakobsenii, L. delbrueckii subsp. lactis, and L. delbrueckii subsp. alloallosunkii (formerly L. delbrueckii subsp. sunkii) (16–18). Among these subspecies, bulgaricus and lactis have been extensively studied as key microorganisms used in the food industry for producing fermented dairy products. By contrast, almost nothing is known about the other four subspecies. Interestingly, L. delbrueckii has rarely been reported as a cause of human infections, almost exclusively as a causative agent of UTIs, mainly in the elderly (10, 11, 13, 19–25). However, current knowledge (clinical relevance, antimicrobial susceptibility) about this species remains very limited. Additionally, it is not known whether human UTIs are due to specific subspecies.
The aim of this study was to (i) assess the clinical relevance of L. delbrueckii by analyzing the clinical and microbiological characteristics of 48 cases of probable UTIs, (ii) evaluate in vitro susceptibility to 15 antimicrobial agents and identify resistance genes using WGS, and (iii) determine the subspecies by comparing three genomic approaches.
MATERIALS AND METHODS
Isolates, media, growth conditions, and identification
We included 48 different clinical isolates of L. delbrueckii collected consecutively from urine samples of patients hospitalized at the Caen University Hospital between 2014 and 2016. From clinical specimens, isolates were grown on 5% horse blood agar plates (bioMérieux, Marcy-l’Etoile, France) and incubated anaerobically or under 5% CO_2_ at 35°C after 48 h of incubation with no difference in growth rates. Identification at the species level was performed using MALDI-TOF mass spectrometry (Microflex LT; Bruker Daltonics, Wissembourg, France; Biotyper software v3.0) after chemical extraction (ethanol, formic acid) in accordance with the manufacturer’s instructions.
Antimicrobial susceptibility testing
MIC values were determined using the broth microdilution method as recommended by the 2022 EUCAST guidelines (http://www.eucast.org/ast_of_bacteria/). Mueller-Hinton broth with lysed horse blood (5%) and β-NAD (20 mg/L) (MHF; bioMérieux) was used, and plates were incubated for 48 h under 5% CO_2_ at 35°C. The following 15 antibiotics were tested: ampicillin, cefotaxime, imipenem, gentamicin, erythromycin, levofloxacin, tetracycline, tigecycline, vancomycin, teicoplanin, linezolid, daptomycin, cotrimoxazole, fosfomycin, nitrofurantoin, and metronidazole. Interpretation of results was based on CLSI breakpoints (https://em100.edaptivedocs.net/Login.aspx) for Lactobacillus spp. (ampicillin, imipenem, erythromycin, vancomycin, linezolid, and daptomycin). For daptomycin, the calcium content of MHF was adjusted to a final concentration of 50 mg/L. Streptococcus pneumoniae ATCC 49619 and L. delbrueckii subsp. delbrueckii CNRZ225 served as internal quality controls for each batch tested.
Genome sequencing, assembly, and annotation
Genomic DNA from the 48 isolates was extracted using the Quick-DNA Fungal/Bacterial Miniprep Kit (Zymo Research, Irvine, CA, USA). DNA libraries were prepared using the NEBNext Ultra DNA Library Prep Kit for Illumina (New England Biolabs, Ipswich, MA, USA) and sequenced as paired-end reads (2 × 300 bp) on an Illumina MiSeq platform with the MiSeq Reagent Kit version 3. To ensure high-quality data, raw Illumina reads were trimmed using Trimmomatic (26) with a sliding window of four bases, discarding bases with a quality score below 20 and retaining reads with a minimum length of 50 bp. Genome assembly was performed using Unicycler v0.5.0 (27) using conservative mode and a minimum contig size of 1,000 bp. Genome annotation was conducted using Prokka (28), applying standard settings optimized for bacterial genomes. The assembled genomes have been deposited in GenBank under project accession PRJNA880510.
To investigate the presence of antimicrobial resistance genes, we used ResFinder 4.0 (29) with a minimum identity threshold of 90% and a minimum length threshold of 70%. Virulence genes were identified by aligning the genome sequences against the VFDB (30) database using BLAST with an E-value cutoff of 1e-5, ensuring the detection of genes with significant homology to known virulence factors.
Comparative genomics and phylogenetics
For comparative analysis, 480 publicly available L. delbrueckii genomes were downloaded from the NCBI database (Table S1). Isolate discrimination was performed using multilocus sequence typing (MLST), average nucleotide identity (ANI), and phylogenomic approaches. In silico, MLST analysis was conducted using seven housekeeping genes (fusA, gyrB, hsp60, ileS, pyrG, recA, and recG) from 40 additional strains described by Tanigawa et al. (31) (Table S2). The concatenated sequences were aligned using MAFFT (32), and a minimum spanning tree was generated using GrapeTree (33) with the MStreeV2 algorithm to visualize the genetic relationships among the isolates.
Genomic similarity was assessed using Mash (34), a rapid tool for estimating pairwise genomic distances, with a sketch size of 1,000 and a k-mer size of 16. Mash outputs were processed using a custom Python script to generate an all-versus-all comparison of genomic distances and ANI values. We used an ANI cutoff below 98% for subspecies differentiation.
Pangenomic analysis was performed using Panaroo (35) with a 98% nucleotide identity cutoff in strict mode to reduce assembly artifacts, and core genome alignment was performed using MAFFT (32). To account for potential recombination events, we used Gubbins (36) to identify and mask regions affected by homologous recombination. Single nucleotide polymorphisms (SNPs) were extracted from the recombination-masked core genome alignment using SNP-sites (37). Pairwise SNP distances between isolates were calculated using PairSNP (https://github.com/gtonkinhill/pairsnp), using SNP alignment as input. A maximum-likelihood phylogenetic tree was constructed using IQ-TREE 2 (38), employing model selection and performing 1,000 ultrafast bootstrap replicates to ensure robust branch support. The resulting phylogenetic tree was midpoint-rooted and visualized using iTOL (39).
RESULTS
Clinical and microbiological characteristics
From 2014 to 2016, 48 clinical isolates of L. delbrueckii were recovered from the urine of 48 patients, including 37 (77%) from mid-stream samples and 11 from indwelling urinary catheters (Table 1). The median age of the patients was 84 years (range 54–96 years), with a large predominance of female patients (sex ratio M/F = 0.04) (Table 1). The majority of infections were community-acquired (n = 40, 83%). Only three (6%) patients had a predisposing urological disease, while 44 (92%) had at least one systemic underlying condition, including heart disease (n = 33, 69%), immunosuppression (n = 29, 60%), diabetes mellitus (n = 15, 31%), malignancy (n = 14, 29%), dementia (n = 12, 25%), and stroke (n = 6, 13%) (Table 1).
Based on clinical and microbiological features, 21 (44%) patients were considered infected and initiated an antibiotic therapy, knowing that physicians were not aware of a potential uropathogenic role of L. delbrueckii. Of the 21 infected patients, 11 (52%) and 10 (48%) suffered from cystitis and pyelonephritis, respectively (Table 1). None of the patients developed severe complications such as endocarditis or urosepsis (data not shown). At the direct examination of the 48 urine specimens, 45 (94%) exhibited more than 10^4^ white blood cells (WBCs) per milliliter, while 43 (90%) showed Gram-positive rods (Table 1). All quantitative cultures had an L. delbrueckii bacterial count of ≥10^5^ CFU/mL, with a median of 10^6^ CFU/mL. L. delbrueckii was recovered in pure culture in 21 (44%) cases, while in mixed cultures, concomitant uropathogens were Gram-negative bacilli (incl. 16 E. coli, 3 K. pneumoniae, 2 E. cloacae complex, 1 P. mirabilis, and 1 C. freundii) or 4 E. faecalis (Table 1).
Antimicrobial susceptibility profiles and treatments
All 48 L. delbrueckii isolates were fully susceptible to ampicillin, imipenem, erythromycin, vancomycin, linezolid, and daptomycin, while the majority exhibited low-level MIC_50_/MIC_90_ values for gentamicin (100%), cefotaxime (92%), tetracycline (90%), and cotrimoxazole (96%) (Table 2). MICs of teicoplanin and tigecycline were also low, suggesting a good in vitro activity of these antibiotics (Table 2). Levofloxacin appeared poorly active in most cases (MICs from 1 to 16 mg/L), while L. delbrueckii was likely intrinsically resistant to both fosfomycin and metronidazole, with MICs >256 mg/L for all tested strains (Table 2). Two isolates (LDEL-06 and LDEL-09) exhibited high MIC values for tetracycline (>16 mg/L), indicating acquired resistance compared to the putative wild-type population (MICs from 0.25 to 4 mg/L) (Table 2). Additionally, two putative resistant isolates (PR-7 and PR-9) were observed for cotrimoxazole (MIC = 8 mg/L), whereas wild-type strains showed MICs from 0.12 to 0.5 mg/L (Table 2).
Genomic analysis
All 48 isolates were unambiguously identified at the species level using MALDI-TOF mass spectrometry; however, phenotypically distinguishing the different subspecies was not possible. Consequently, the genomes of all 48 isolates were sequenced and analyzed. The mean genome size was 1.99 ± 0.06 Mb with a G+C content of 49.9% ± 0.1%, and the mean number of coding sequences was 1,911 ± 49 (Table 3). The core genome comprised 1,123 orthologous gene families, while the pan-genome was open, consisting of 5,616 orthologous gene families.
Subspecies identification was initially performed using MLST, revealing that most L. delbrueckii clinical isolates belonged to the subspecies allosunkii (n = 40; 83%), followed by L. delbrueckii subsp. lactis (n = 8; 17%) (Fig. 1). This distribution was further confirmed through ANI analysis. The ANI values for the 40 strains identified as allosunkii, compared to the L. delbrueckii subsp. allosunkii reference strain JCM 17838 ranged from 98.12% to 98.74%. Similarly, ANI values for the eight strains identified as lactis, relative to the L. delbrueckii subsp. lactis reference strain DSM 20072, ranged from 98.18% to 98.68% (Fig. 2; Fig. S1; Table S3). The subspecies distribution was also supported by a core genome SNP-based phylogenomic approach (Fig. 3), which included an analysis of 480 additional L. delbrueckii genomes. To assess genomic diversity within L. delbrueckii subsp. allosunkii (n = 40), a phylogenetic tree was constructed based on the alignment of 10,187 recombination-masked SNPs from 1,207 core gene sequences. This analysis revealed three major lineages, each comprising distinct sublineages (Fig. 4). The average genome sizes for lineages 1, 2, and 3 were 1.88, 1.94, and 1.97 Mb, respectively (Fig. 4). Notably, four clades of clonally-related strains were identified: clade A (2 isolates, 7 SNPs), clade B (5 isolates, 24 to 55 SNPs), clade C (2 isolates, 1 SNP), and clade D (18 isolates, 3 to 58 SNPs) (Fig. 4; Table S4). These findings highlight the genomic diversity and structure within the L. delbrueckii subsp. allosunkii population.
Minimum spanning tree based on MLST data from 568 L. delbrueckii isolates. The collection of isolates includes 480 isolates from NCBI, 40 strains described by Tanigawa et al. (Table S2) (31), and the 48 clinical isolates of the study. The seven housekeeping genes (fusA, gyrB, hsp60, ileS, pyrG, recA, and recG) were aligned against all genomes, and the tree was built from concatenated sequences using GrapeTree (https://github.com/achtman-lab/GrapeTree). Clinical isolates of the study are colored in yellow.
ANI analysis of the 48 isolates compared with L. delbrueckii reference subspecies genomes. The maximum-likelihood phylogenetic tree was constructed using a core genome SNP-based approach with IQ-TREE 2 (TVM+F+ASC+G4 best-fit model) and 1,000 ultrafast bootstrap iterations (http://www.iqtree.org/). The tree was visualized using iTOL (https://itol.embl.de). The reference strains included are L. delbrueckii subsp. allosunkii JCM 17838, L. delbrueckii subsp. lactis DSM 20072, L. delbrueckii subsp. bulgaricus ATCC 11842, L. delbrueckii subsp. delbrueckii DSM 20074, L. delbrueckii subsp. indicus JCM 15610, and L. delbrueckii subsp. jakobsenii DSM 26046.
Core genome SNP-based maximum-likelihood tree of 528 L. delbrueckii isolates. This data set comprises 480 isolates from NCBI and 48 clinical isolates from this study. The tree was constructed using IQ-TREE 2 (GTR+F+ASC+G4 best-fit model) with 1,000 ultrafast bootstrap iterations (http://www.iqtree.org/), based on 132,691 polymorphic sites within the core genome. Visualization was performed using iTOL (https://itol.embl.de). Clinical isolates from this study are indicated by black rectangles, and their names are highlighted in yellow.
Maximum-likelihood phylogenetic tree based on the alignment of 10,137 recombination-masked SNPs derived from 1,207 core gene sequences from the 40 L. delbrueckii subsp. allosunkii clinical isolates. A heatmap is provided to illustrate SNP distances among the isolates. The three primary lineages (1–3) and four clades (A–D) are indicated.
Both tetracycline-resistant clinical isolates (LDEL-06 and LDEL-09) belonged to the subspecies allosunkii and harbored the tet(W) gene on their chromosomes, which encodes a ribosomal protection protein that confers resistance by preventing tetracycline from binding to the ribosome (Table S5). By analysis of sequences from NCBI, we also identified a tet(W) gene in the strain L. delbreuckii subsp. indicus ME-792 (Table S5). For cotrimoxazole, no acquired resistance genes or mutations were detected. Finally, no obvious virulence genes were found (Table S6).
DISCUSSION
Similar to the aforementioned emerging Gram-positive uropathogens, lactobacilli are likely underreported in the medical literature as they are typically regarded as commensals or contaminants, leading to the absence of an actual prevalence. For information, the prevalence over the period of these “minor” uropathogens represented less than 2% of bacterial species recovered from urine specimens, incl. A. urinae (1%), A. schaalii (0.4%), L. delbrueckii (0.2%), and A. sanguinicola (0.2%) (data not shown). The most commonly reported infections due to lactobacilli are bacteremia and infective endocarditis, with L. rhamnosus, L. paracasei, L. plantarum, L. fermentum, L. casei, Lactobacillus crispatus, Lactobacillus johnsonii, L. jensenii, L. acidophilus, Ligilactobacillus salivarius (formerly Lactobacillus salivarius), L. gasseri, and Latilactobacillus curvatus (formerly Lactobacillus curvatus) being the most frequent species (4–6, 8–10, 14, 16). Studies on virulence have mainly focused on L. rhamnosus and L. paracasei (12). These species possess potential virulence factors, such as enzymes that break down human glycoproteins and proteins that bind to extracellular proteins such as fibronectin, fibrinogen, and collagen, facilitating early-stage colonization and adherence. Some strains also have the ability to aggregate human platelets and produce biofilm (13).
Recent metagenomic analyses revealed that L. delbrueckii inhabits the intestines of humans and animals (40). In healthy individuals, this species represents the fourth most prevalent species among lactobacilli after Ligilactobacillus ruminis (formerly Lactobacillus ruminis), L. casei, and L. gasseri in the gut microbiota (40). UTIs caused by lactobacilli are very uncommon, with the vast majority attributed to L. delbrueckii (only 14 reported cases) (19–25). Among these 14 patients, there was a predominance of women (12/14, 86%), and most patients were elderly, with a median age of 82 years (range: 49–94 years). The two male patients suffered from acute bacterial prostatitis (23, 25). For all patients, urine dipsticks were negative for nitrite detection, consistent with the fact that lactobacilli do not reduce nitrate to nitrite (16). As reported in our study, urinalysis showed in all cases pyuria, positive direct examination with Gram-positive rods, and significant culture (>10^5^ CFU/mL), usually monobacterial (19–25). The main putative risk factors reported here and in previous case reports are old age, diabetes mellitus, hypothyroidism, dementia, benign prostate hyperplasia, and urolithiasis (19, 21–25). Invasive infections caused by L. delbrueckii are much less frequently reported, with only a few cases of bacteremia described (8–10, 24), and no invasive infections were observed in our case series.
Only two subspecies were involved in UTIs in our study. L. delbrueckii subsp. allosunkii was first isolated from sunki, a traditional Japanese non-salted pickle, while L. delbrueckii subsp. lactis is heavily used for the production of Parmesan- and Emmental-type cheeses (15, 17, 18). Out of the 14 cases of UTIs caused by L. delbrueckii reported in the literature, the subspecies has been mentioned in only three cases (one lactis, one delbrueckii, and one allosunkii). This is surprising since the identification in these three clinical cases was only obtained by MALDI-TOF mass spectrometry (22–24), which is not a reliable method for identification at the subspecies level.
Genomic analysis of our collection revealed a mean genome size of 1.99 ± 0.06 Mb, a G+C content of 49.9% ± 0.1%, and a core genome of 1,123 genes, similar to what has been recently described in a comparative analysis of 31 L. delbrueckii genomes, which reported a genome size of around 1.93 ± 0.16 Mb, a G+C content of 49.8% ± 0.4%, and a core genome of 1,069 orthologous gene families (17).
According to the type of sugar fermentation pathway, lactobacilli are separated into three major metabolic groups that form distinct phylogenetic clades: obligately homofermentative, facultatively heterofermentative, and obligately heterofermentative (12, 15). Interestingly, homofermentative lactobacilli (e.g., L. acidophilus, Lactobacillus amylovorus, L. crispatus, L. delbrueckii, L. gasseri, Lactobacillus helveticus, L. jensenii, L. johnsonii) are susceptible to vancomycin and teicoplanin, whereas heterofermentative lactobacilli (e.g., Levilactobacillus brevis [formerly Lactobacillus brevis], L. casei, L. curvatus, L. fermentum, L. paracasei, Lactiplantibacillus pentosus [formerly Lactobacillus pentosus], L. plantarum, Limosilactobacillus reuteri [formerly Lactobacillus reuteri], L. rhamnosus, Latilactobacillus sakei [formerly Lactobacillus sakei], L. salivarius, Lacticaseibacillus zeae [formerly Lactobacillus zeae]) are inherently highly resistant to these antibiotics (4–6, 9, 41–43). As anticipated for L. delbrueckii, all isolates in our study demonstrated susceptibility to vancomycin (MIC_50_/90: 0.25/0.5 mg/L) and teicoplanin (MIC_50_/90: 0.12/0.12 mg/L). Lactobacilli are also intrinsically susceptible to aminopenicillins, imipenem, erythromycin, tetracycline, and linezolid, whereas the in vitro activity of cephalosporins, gentamicin, fluoroquinolones, cotrimoxazole, and nitrofurantoin appears more variable depending on the species (4–6, 9, 41, 43, 44). Among newer antibiotics, tigecycline and linezolid typically exhibit good activity, whereas daptomycin shows variable efficacy (42). Finally, the intrinsic resistance to metronidazole of lactobacilli is well documented (41, 42).
Most data on L. delbrueckii originates from food isolates, particularly the subspecies bulgaricus and lactis. Though few human isolates have been tested for antimicrobial susceptibility, they seem to share similar profiles with food isolates, showing susceptibility to ampicillin, imipenem, gentamicin, erythromycin, tetracycline, vancomycin, linezolid, and daptomycin (9, 11, 19, 21, 24, 25, 42, 45). Fluoroquinolones exhibit variable activity, with ciprofloxacin being less active than levofloxacin/moxifloxacin, and some evidence of acquired resistance exists (9, 11, 19). Notably, high-level resistance to fosfomycin has been reported (20, 22, 23).
Tetracycline and erythromycin resistance genes are the most common in lactobacilli, with tet(M) and erm(B) being the most widespread and often genetically linked (42, 46). Few resistance genes have been found in some food isolates of L. delbrueckii subsp. bulgaricus such as tet(M), aph(3’)-III, and ant(6) (42, 45). Together with our results, this suggests that acquired resistance in L. delbrueckii is uncommon.
UTIs caused by L. delbrueckii have been successfully treated with β-lactams (aminopenicillins, piperacillin-tazobactam, cefixime) or clarithromycin, indicating that the recommended treatment for L. delbrueckii-related infections should rely on β-lactams, particularly aminopenicillins (20–22, 24). It is important to note that recurrent UTIs caused by L. delbrueckii were often linked to treatment failures with fluoroquinolones, fosfomycin, or cefuroxime (19, 21–23, 25).
Despite the retrospective observational monocentric design, this study shows the potential uropathogenic role of L. delbrueckii (especially the subspecies allosunkii) in UTIs among elderly patients and those with predisposing conditions. We also demonstrate that these clinical isolates are generally susceptible to many antibiotics, with only a few intrinsic and acquired resistances. Based on in vitro data, the antibiotic treatment of these infections should rely on aminopenicillins rather than fosfomycin or fluoroquinolones.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Flores-Mireles AL, Walker JN, Caparon M, Hultgren SJ. 2015. Urinary tract infections: epidemiology, mechanisms of infection and treatment options. Nat Rev Microbiol 13:269–284. doi:10.1038/nrmicro 343225853778 PMC 4457377 · doi ↗ · pubmed ↗
- 2Kline KA, Lewis AL. 2016. Gram-positive uropathogens, polymicrobial urinary tract infection, and the emerging microbiota of the urinary tract. Microbiol Spectr 4. doi:10.1128/microbiolspec.UTI-0012-2012 PMC 488887927227294 · doi ↗ · pubmed ↗
- 3Vaughan EE, Heilig HGHJ, Ben-Amor K, de Vos WM. 2005. Diversity, vitality and activities of intestinal lactic acid bacteria and bifidobacteria assessed by molecular approaches. FEMS Microbiol Rev 29:477–490. doi:10.1016/j.femsre.2005.04.00916125009 · doi ↗ · pubmed ↗
- 4Salminen MK, Rautelin H, Tynkkynen S, Poussa T, Saxelin M, Valtonen V, Järvinen A. 2004. Lactobacillus bacteremia, clinical significance, and patient outcome, with special focus on probiotic L. rhamnosus GG. Clin Infect Dis 38:62–69. doi:10.1086/38045514679449 · doi ↗ · pubmed ↗
- 5Felten A, Barreau C, Bizet C, Lagrange PH, Philippon A. 1999. Lactobacillus species identification, H 2O 2 production, and antibiotic resistance and correlation with human clinical status. J Clin Microbiol 37:729–733. doi:10.1128/JCM.37.3.729-733.19999986841 PMC 84537 · doi ↗ · pubmed ↗
- 6Salminen MK, Rautelin H, Tynkkynen S, Poussa T, Saxelin M, Valtonen V, Järvinen A. 2006. Lactobacillus bacteremia, species identification, and antimicrobial susceptibility of 85 blood isolates. Clin Infect Dis 42:e 35–44. doi:10.1086/50021416447101 · doi ↗ · pubmed ↗
- 7Anderson AC, Sanunu M, Schneider C, Clad A, Karygianni L, Hellwig E, Al-Ahmad A. 2014. Rapid species-level identification of vaginal and oral lactobacilli using MALDI-TOF MS analysis and 16S r DNA sequencing. BMC Microbiol 14:312. doi:10.1186/s 12866-014-0312-525495549 PMC 4272787 · doi ↗ · pubmed ↗
- 8Cannon JP, Lee TA, Bolanos JT, Danziger LH. 2005. Pathogenic relevance of Lactobacillus: a retrospective review of over 200 cases. Eur J Clin Microbiol Infect Dis 24:31–40. doi:10.1007/s 10096-004-1253-y 15599646 · doi ↗ · pubmed ↗